
Abstract
Expansion of the coyote’s (Canis latrans) distribution in North America has included most urban areas. Concerns for human safety have resulted in the need to understand the spatial relationship between humans and coyotes in urban landscapes. We examined the space use of coyotes with varying degrees of urban development in the Chicago metropolitan area, IL, USA, between March 2000 and December 2002. We compared home-range size, land use, and habitat use of 41 radio-collared coyotes (5 coyotes residing in developed areas, 29 in less-developed areas, and 7 in a matrix of developed and less-developed areas). The partitioning of coyotes into groups based on their level of exposure to urban development allowed us to examine if differences in use of land types by coyotes was evident in our study area. Coyotes in developed areas had home ranges twice the size of animals in less-developed areas. Nonurban habitats were used by all coyotes in the study area, while urban land was avoided. Coyotes in developed areas had large home ranges and high amounts of urban land in their range, but preferred nonurban habitat. This required the coyotes to travel through a matrix of urban land, thus encountering human activity and possibly increasing the risk of conflict with humans. However, coyotes in developed areas avoided crepuscular times when human activity was highest, suggesting that coyotes in developed areas may reduce conflicts with humans by traveling through the matrix of urban land late at night when the risk of contact with humans is lowest. Coyotes in less-developed areas were less affected by human activity at night and likely posed less risk to humans.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Coyotes (Canis latrans) are opportunistic carnivores that have adapted to the encroachment of urbanization, as well as inhabiting urban landscapes that were previously unoccupied. Coyotes have been documented in large metropolitan areas from Los Angeles to New York City (Howell 1982; Gehrt 2004). Although their use of commercial areas is uncommon (Quinn 1997a; Grinder and Krausman 2001a), they have been found in Central Park in New York City (Martin 1999). Coyotes are found in the suburbs where cover and food are available (Gehrt et al. 2009). These suburbs provide habitat for coyotes in forest preserves, city parks, golf courses, and wooded residential areas, and abundant water sources from streams, lakes, ponds, and wetlands. Suburbs also provide natural prey (e.g., rodents, deer, leporids), as well as human-associated foods such as pets, garbage, vegetable gardens, and pet food (McClure et al. 1995; Quinn 1997b; Morey et al. 2007).
The close proximity of coyotes to humans in urban environments has caused some coyote–human conflicts (Carbyn 1989; Baker and Timm 1998; Timm et al. 2004). Problems with nuisance coyotes have gained the attention of many city officials. Coyotes have become a concern in many urban areas for being where they are not wanted or where they pose potential risks to humans and pets. The perceived threat of coyotes to humans is a concern shared by many residents in metropolitan areas. Attacks on domestic pets are more common in urban areas (Andelt and Manan 1980; Grinder 1999). Coyotes can also carry diseases that can threaten humans and pets (Holzman et al. 1992a; Clark et al. 1994; Grinder and Krausman 2001b).
Knowledge of the factors affecting the potential interactions of coyotes and humans in urban environments is of increasing importance because of the continuing urbanization of natural environments. To understand the potential for these interactions, an examination of the spatial and temporal use of urban landscapes by coyotes is necessary. By examining urban landscape use by coyotes, managers will have a better understanding of where coyotes are in the urban landscape, the proximity of coyotes to humans and their activities, what landscape characteristics are important for their existence, and their role as a predator on native and domestic prey. This will aid in understanding where and when coyotes pose risks to humans, and what landscape features can be managed to control coyote populations.
There are many types of land uses (e.g., forest preserves, urban, undeveloped, agricultural) in the Chicago metropolitan area, representing a variety of human activities. Documenting how coyotes use this human landscape provides an understanding of where coyotes are in relation to areas of high and low human use. It is also important to know where coyotes are within these landscapes. By examining habitat use (e.g., woods, riparian, natural grass) by urban coyotes, we may explain which habitats they are using, which are important for their existence, and how they inhabit urban areas that by appearance seem inhospitable to a medium-size carnivore. The purpose of this study is to examine the space use by coyotes in a matrix of developed and less-developed environments in the Chicago metropolitan area, and how this use may influence the potential for conflicts with humans. We investigated this by examining home-range size, and space use of coyotes at both the land type (i.e., human use) and habitat type (i.e., vegetative cover) scales.
Materials and methods
Study area
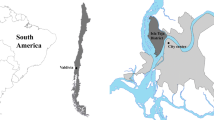
We conducted our study in western Cook, northern Dupage, eastern Kane, and southern McHenry counties in northeast IL. These counties are part of the greater Chicago metropolitan area, which is the third largest US metropolitan area with 9.1 million people (US Census Bureau 2000). Winter temperatures and precipitation averaged −3 °C and 3.8 cm/month, respectively, and summer temperatures and precipitation averaged 19 °C and 9.9 cm/month, respectively. The study area was defined by a minimum convex polygon drawn around the outer limits of all coyote home-range polygons in our study (Fig. 1). Land types, as defined by the type of human activity, included developed urban land (64 %), agriculture (14 %), protected forest preserves (13 %), undeveloped land (5 %), and water (4 %). Habitat types, as defined by vegetation cover, included impervious surfaces (33 %), woods (23 %), natural grass (19 %), crops (11 %), manicured grass (7 %), and riparian (7 %). Impervious surfaces consisted of roads, parking lots, and buildings. Manicured grass habitat included lawns such as golf courses, city parks, and residential areas.

Location of study area encompassing the home ranges of all radio-collared coyotes tracked in the Chicago metropolitan area, northeastern IL, USA, 2000–2002
The majority of coyotes we studied resided in 2 public forest preserves, 1 private preserve in eastern Kane County, and a town in western Cook County. The forest preserves included Poplar Creek Forest Preserve, Ned Brown Forest Preserve, and Max McGraw Wildlife Foundation. Ned Brown and Poplar Creek were open to the public and received recreational use during the summer. Ned Brown was 1,499 ha in size and was located approximately 5 km west of Chicago-O’Hare International Airport. Ned Brown was surrounded by high-density residential and commercial areas, and was bordered on 2 sides by 8-lane highways. Poplar Creek received considerably fewer visitors per year; it was 1,825 ha in size and was located on the western border of Cook County. Poplar Creek was bordered by medium-density housing, a commercial area, and an 8-lane highway. Max McGraw consisted of 495 ha and was located on the eastern border of Kane County. This privately owned area was managed as a natural area and hunting–fishing preserve, although coyotes were not hunted or controlled on the property. The Village of Schaumburg was located between Ned Brown and Poplar Creek, and consisted of a human population of 75,400 distributed in medium-density residential and commercial areas. It included 58 small city parks, 2 golf courses, 4 small nature areas, and a water treatment plant.
Animal capture and telemetry
We captured coyotes using padded foothold traps and nonlethal neck snares from March 2000 to October 2002. Coyotes were immobilized for handling, weighed, measured, aged by tooth wear (Gier 1968), sexed, ear tagged, and fitted with a radiocollar (Advanced Telemetry Systems, Isanti, MN, USA). We classified coyotes as subadults (<2 years old) and adults. We released all coyotes at the capture location. The Institutional Animal Care and Use Committee at Utah State University approved the capture and handling protocol. We located radioed coyotes from vehicles equipped with 4-element Yagi antennas following the procedures of White and Garrott (1990). We determined locations by triangulating ≥2 bearings using LOCATE II (Pacer, Truro, NS, Canada). Location estimation error from reference collars was 108 ± 87 m (SD).
We acquired diurnal point and nocturnal sequential locations on radio-collared coyotes (Gese et al. 1990). Diurnal locations were recorded ≥3 times/week and separated by one night. Urban coyotes have been shown to be less active in urban landscapes during diurnal times (Shargo 1988; Quinn 1997a; Grinder 1999); we therefore concentrated sequential tracking sessions during crepuscular and nocturnal times. Because of potential triangulation error associated with tracking moving coyotes, time between bearings was limited to ≤3 min (\( \bar{X} \) = 2 min). Coyotes have shown increased movement rates near midnight and just before dawn in urban areas (Grinder 1999). Thus, to observe variation in activity patterns throughout the night, tracking from 1800 to 0600 was divided into 6 2-h periods and locations were recorded every 1 h for the 2-h period. We allowed >1 night between tracking periods on the same coyote.
Estimation of home-range size
We used locations from diurnal and nocturnal radio-tracking sessions to calculate seasonal home-range estimates (Gese et al. 1990; Reynolds and Laundre 1990). We used the home-range extension (Rodgers and Carr 2002) for ArcView 3.2 geographical information system (GIS) software (Environmental Systems Research Institute, Redlands, CA, USA) to plot 95 % fixed-kernel (FK) home-range estimates (Fig. 2; Worton 1989). We used individual home ranges in analyses if area-observation curves reached an asymptote (Odum and Kuenzler 1955).

Example of 95 % kernel home-range isopleths and locations for 2 coyotes in developed and less-developed land use groups in the Chicago metropolitan area, northeastern IL, USA, 2000–2002
Preliminary radio-tracking showed that certain coyotes stayed almost exclusively in forest preserves and undeveloped areas, others occupied urban areas comprising almost entirely residential, city parks, and commercial areas, while others utilized a mix of land-use types. To examine space use among these groups, we classified each coyote into 1 of 3 landscape groups, similar to Gese et al. (1988) and Morey et al. (2007). We classified a coyote into the “developed” group if >70 % of its home range consisted of developed land (urban land), “less-developed” if >70 % of its home range consisted of less-developed land types (agriculture, forest preserve, undeveloped), and “mixed” if its home range consisted <70 % of either type. We examined space use among biological seasons: breeding (January 1–April 30), pup-rearing (May 1–August 31), and dispersal (September 1–December 31) (adapted from Laundrè and Keller 1981). We classified a coyote as a resident if it used one unique area (i.e., a home range) for ≥1 biological season, and a transient if it did not settle in one area within one season (Gese et al. 1988).
Coyote space use
We used 2 types of classifications to examine coyote space use: “land” type was based upon the degree of human use, while “habitat” type was based upon vegetative structure; both parameters were equally important to how coyotes used the landscape but were ecologically different. We created land type and habitat type maps with 28.5 m resolution from 1997 Chicago Wilderness/NASA Landsat Thematic Mapper images for use in ArcView GIS software. We reclassified 164 Landsat categories into 5 land types based on the degree of human use (agriculture, forest preserve, undeveloped, urban land, water) and 6 habitat types based on vegetative structure (crops, impervious surfaces, natural grass, riparian, manicured grass, woods). We calculated seasonal percentage of use per type (e.g., land, habitat) for each animal by overlaying telemetry points on GIS maps.
We used compositional analysis (Aebischer and Robertson 1992; Aebischer et al. 1993) using Resource Selection for Windows software (Leban 1999) to investigate use versus availability of land types and habitat types. For our compositional analyses we used the coyote’s seasonal use as our sampling unit and examined use at two orders of selection (Johnson 1980): second-order selection (i.e., use within the home range versus availability in the study area) and third-order selection (i.e., use versus availability within the home range). We used our study area polygon (Fig. 1) to calculate availability for both land and habitat types within the study area (i.e., 2nd-order selection). We calculated land and habitat availability for home ranges from seasonal home ranges for each coyote (i.e., 3rd-order selection). We calculated Wilks’ likelihood ratios (Λ) to examine selection of the land types and habitat types within landscape groups, and a ranking matrix of t tests was formed to rank the selection of land types and habitat types (Aebischer et al. 1993).
Compositional analysis (Aebischer et al. 1993) requires a minimum of 6 individuals per group for proper statistical analyses. Therefore, we used only groups with a minimum sample size of 6 coyotes. Because use and availability of land types and habitat types were defined by home-range polygons, we excluded coyotes without adequate locations to define a home range from compositional analyses. Also, transient coyote home ranges were too large to infer meaningful estimates of available habitat, and thus they were excluded.
Results
We recorded 15,723 diurnal and nocturnal locations from 41 resident coyotes (16 F, 25 M; 17 adults, 20 subadults; 4 monitored as both adults and subadults) having adequate sample sizes where the home-range area-observation curve reached an asymptote. Five (2 F, 3 M; 5 adults) of these coyotes were in developed areas, 27 (10 F, 17 M; 8 adults, 15 subadults, and 4 monitored as subadult then adult) in less-developed areas, 7 (4 F, 3 M; 2 adults, 5 subadults) in mixed areas, and 2 (2 M; 1 adult, 1 subadult) coyotes were found in less-developed areas and a mix of land-use areas during different periods of the study. They were located an average of 74 times/season (range = 31–135) and tracked for an average of 11.8 months (range = 4–32 months).
Home-range size
We estimated 114 seasonal home ranges for 41 resident coyotes and 13 seasonal home ranges for 10 transient coyotes. During the breeding season, resident home-range size for female coyotes [\( \bar{X} \) = 11.2 km2 ± 5.5 (±SD)] was not different than male coyotes (\( \bar{X} \) = 10.5 ± 6.0 km2; t 26 = −0.305, P > 0.35). During the pup-rearing season, resident home-range size for females (\( \bar{X} \) = 8.2 km2 ± 5.7) was not different than males (\( \bar{X} \) = 9.4 ± 7.7 km2; t 43 = 0.567, P > 0.25). Similarly, during the dispersal season, resident home-range size for female coyotes (\( \bar{X} \) = 10.8 km2 ± 6.8) was not different than male coyotes (\( \bar{X} \) = 10.5 ± 7.0 km2; t 40 = −0.150, P > 0.40). Home-range size of resident coyotes was smaller than transients (t 49 = 4.12, P < 0.01). Home-range size of resident coyotes averaged 9.4 km2 ± 0.9 (±SE; range = 1.0–27.5 km2) while that of transients averaged 80.4 km2 ± 18.7 (range = 29.5–272.3 km2). In general, coyotes in developed areas had home ranges approximately 2× the size of coyotes in less-developed areas for all seasons (Table 1). Coyotes in mixed areas showed more variability compared with the other landscape groups across seasons, with the pup-rearing home-range size being the smallest.
Selection of land types
For second-order selection, the utilization of land types by coyotes (Fig. 3) in the developed group differed from study area availability (Λ = 0.021, df = 4, P < 0.001). Urban land ranked highest, but was not significant when ranked next to undeveloped land, although urban land use was considerably higher than all land types. Coyotes in the less-developed group also used land types out of proportion with study area availability (Λ = 0.097, df = 4, P < 0.001); forest preserves and urban land were ranked first and second, respectively. Although urban land was ranked above water, agriculture, and undeveloped land types, its availability was much higher than its use. The use of land types by coyotes in the mixed landscape group also differed from study area availability (Λ = 0.296, df = 4, P < 0.001). Although undeveloped areas and water ranked first and second, other land types, particularly urban land, were utilized in high proportions.

Compositional analysis rankings (left to right = highest to lowest) of 2nd-order land type selection by coyotes in 3 groups, Chicago metropolitan area, IL, USA, 2000–2002. Underlined land types indicate classes not significantly different in their rankings. Land types: Ag agriculture, FP forest preserves, Und undeveloped, UrLand urban land, Water water. Error bars represent 1 SE
At the third-order selection of land types (Fig. 4), some coyotes lacked the water category in their home range. To correct this problem for compositional analyses, this category was removed from the dataset; it represented <4 % of total locations. The third-order selection of land types by coyotes in the developed group differed from home range availability (Λ = 0.296, df = 3, P < 0.001). Although undeveloped areas were ranked significantly higher than other land types, urban land was used more often, even though availability was much higher. The third-order selection of land types by the coyotes in the less-developed group also differed from home range availability (Λ = 0.222, df = 3, P < 0.001). Forest preserves were ranked the highest for this group and were selected in greater proportion than their availability; urban land was ranked second but was selected less than available. The third-order selection of land types by coyotes in the mixed group also differed from availability (Λ = 0.149, df = 3, P < 0.001), but selection was not dominated by a single category as seen in the other groups. Use of land types during diurnal and nocturnal periods (Fig. 5) was similar to the overall land type selection patterns (Fig. 4). An exception was coyotes in the developed group, which selected undeveloped land types above other land types during diurnal periods (Λ = 0.193, df = 3, P < 0.001) and random use of land types during nocturnal periods (Λ = 0.845, df = 3, P = 0.470).

Compositional analysis rankings (left to right = highest to lowest) of 3rd-order land type selection by coyotes in 3 groups, Chicago metropolitan area, IL, USA, 2000–2002. Underlined land types indicate classes not significantly different in their rankings. Land types: Ag agriculture, FP forest preserves, Und undeveloped, UrLand urban land. Error bars represent 1 SE

Compositional analysis rankings (left to right = highest to lowest) of 3rd-order diurnal and nocturnal land type selection by coyotes in 3 groups, Chicago metropolitan area, IL, USA, 2000–2002. Underlined land types indicate classes not significantly different in their rankings. Land types: Ag agriculture, FP forest preserves, Und undeveloped, UrLand urban land. Error bars represent 1 SE
Selection of habitat types
For second-order selection, habitat use by each group differed from study area availability (developed: Λ = 0.018, df = 5, P < 0.001; less-developed: Λ = 0.055, df = 5, P < 0.001; mixed: Λ = 0.157, df = 5, P < 0.001). Home ranges for coyotes in developed areas contained more impervious surfaces and manicured grass than was available in the study area, and coyotes in both less-developed and mixed areas contained more nonurban habitats, with riparian and natural grass ranking at the top (Fig. 6). The home ranges of coyotes in both less-developed and mixed areas contained less impervious surface and manicured grass than occurred in the study area. Both adults and subadults had significant habitat selection (adults: Λ = 0.232, df = 5, P < 0.001; subadults: Λ = 0.108, df = 5, P < 0.001), but lacked major selection differences between them. The only major difference was that adults had more impervious surface areas than subadults, and subadults had more wooded areas than adults in their home ranges (Fig. 7). Habitat selection differences between sexes were less obvious. Both males and females had more riparian and natural grass in their home ranges than in the study area, and both contained less impervious surfaces and manicured grass (male: Λ = 0.206, df = 5, P < 0.001; female: Λ = 0.291, df = 5, P < 0.001).

Compositional analysis rankings (left to right = highest to lowest) of 2nd-order habitat type selection by coyotes in 3 groups, Chicago metropolitan area, IL, USA, 2000–2002. Underlined habitat types indicate classes not significantly different in their rankings. Habitat types: Crop crop, ImpSurf impervious surface, NaGrass natural grass, Rip riparian, MnGrass manicured grass, Woods woods. Error bars represent 1 SE

Compositional analysis rankings (left to right = highest to lowest) of 2nd-order habitat type selection by adult and subadult coyotes, Chicago metropolitan area, IL, USA, 2000–2002. Underlined habitat types indicate classes not significantly different in their rankings. Habitat types: Crop crop, ImpSurf impervious surface, NaGrass natural grass, Rip riparian, MnGrass manicured grass, Woods woods. Error bars represent 1 SE
All groups had significant rankings of habitats (developed: Λ = 0.061, df = 5, P < 0.001; less-developed: Λ = 0.205, df = 5, P < 0.001; mixed: Λ = 0.056, df = 5, P < 0.001), although coyotes in the less-developed areas utilized habitats in similar proportions to availability (Fig. 8). Even though rankings differed for each group, natural grass was utilized the most by all groups. Coyotes in both less-developed and mixed areas used natural grass in close proportion to its availability, while coyotes in developed areas used it >2 times its availability. Both impervious surfaces and manicured grass areas were utilized less than its availability for all groups. Both adults and subadults showed significant habitat rankings (adults: Λ = 0.132, df = 5, P < 0.001; subadults: Λ = 0.221, df = 5, P < 0.001). Although habitat rankings between age classes differed, there was little difference between utilization and availability of each habitat type. There were also significant differences between seasonal habitat rankings (breeding: Λ = 0.100, df = 5, P < 0.001; pup-rearing: Λ = 0.125, df = 5, P < 0.001; dispersal: Λ = 0.141, df = 5, P < 0.001). Similar to age class rankings, there were few differences between seasons (Fig. 9). Differences included: riparian areas were selected more often during the breeding season than other seasons, and crops were selected more during pup-rearing and dispersal than during breeding. We found that habitat use between diel periods by coyotes in less-developed areas was significantly different (day: Λ = 0.252, df = 5, P < 0.001; night: Λ = 0.324, df = 5, P < 0.001), although use and availability percentages were not different biologically between diel periods for this group (Fig. 10).

Compositional analysis rankings (left to right = highest to lowest) of 3rd-order habitat type selection by coyotes in 3 groups, Chicago metropolitan area, IL, USA, 2000–2002. Underlined habitat types indicate classes not significantly different in their rankings. Habitat types: Crop crop, ImpSurf impervious surface, NaGrass natural grass, Rip riparian, MnGrass manicured grass, Woods woods. Error bars represent 1 SE

Compositional analysis rankings (left to right = highest to lowest) of 3rd-order seasonal habitat type selection by coyotes, Chicago metropolitan area, IL, USA, 2000–2002. Underlined habitat types indicate classes not significantly different in their rankings. Habitat types: Crop crop, ImpSurf impervious surface, NaGrass natural grass, Rip riparian, MnGrass manicured grass, Woods woods. Error bars represent 1 SE

Compositional analysis rankings (left to right = highest to lowest) of 3rd-order habitat type selection by coyotes in the less-developed group, Chicago metropolitan area, IL, USA, 2000–2002. Underlined habitat types indicate classes not significantly different in their rankings. Habitat types: Crop crop, ImpSurf impervious surface, NaGrass natural grass, Rip riparian, MnGrass manicured grass, Woods woods. Error bars represent 1 SE
Discussion
We found seasonal differences among home-range sizes of coyotes, with the pup-rearing period having the smallest home-range size. The home-range size of coyotes in both developed and less-developed areas fluctuated slightly by season, but was highly variable for coyotes in the mixed group. This high seasonal fluctuation could have been explained if coyotes in mixed areas were overrepresented by coyotes that were semitransient with unstable home ranges. However, we did not find that coyotes in mixed areas contained more of these animals than other groups. There are, however, reasons why coyote populations have seasonal fluctuations. Small home ranges have been documented for coyotes in summer or pup-rearing months, possibly due to den-site fidelity of pack members (Person and Hirth 1991), small home-range sizes of juveniles (Andelt 1985), or cyclic prey (McNab 1963).
Coyotes in developed areas had home ranges 2× the size of coyotes in less-developed areas. The resource dispersion hypothesis (MacDonald 1983; Carr and MacDonald 1986) suggests that areas with dispersed habitat patches require animals to have larger home ranges to cover enough patches to meet their basic needs. Studies have found that coyotes prefer habitats with the most prey (Litvaitis and Shaw 1980; Andelt and Andelt 1981), home-range sizes increase at low prey densities (McNab 1963; Meia and Weber 1995), and home ranges containing habitat with the lowest prey density are the largest (Gese et al. 1988). In an urban area in southern California, Riley et al. (2002) found a positive correlation between home-range size and urban-associated areas. Developed urban areas may provide either lower-quality habitat or a matrix of dispersed habitat patches. Coyotes in these areas may find it necessary to occupy and defend larger areas to meet their feeding, resting, and denning needs. We found that coyotes in developed areas had larger home ranges than animals in less-developed areas because developed areas may contain dispersed resources or lower quality of food resources.
The difference between resident and transient home ranges was expected since our classification of a coyote as a transient was determined by its widely fluctuating home-range size. Canids, particularly subadults, disperse from their natal territory and will often cover large areas (Gese et al. 1988). This may be due to their occupancy of inferior-quality habitat or widely dispersed resources, or their search for available territories.
Coyotes in our study area occupied home ranges with varying degrees of human activity and use. We found that some home ranges consisted almost exclusively of residential and commercial areas, while other coyotes in adjacent territories occupied almost exclusively less-developed areas. Coyotes preferred less-developed areas with low levels of human activity over urban land which had high levels of human activity. The higher selection of less-developed areas may be due to higher quality or quantity of habitat for resting, denning, foraging, and escape cover to avoid humans. However, some coyotes occupied large areas of developed areas with high levels of human use. Although coyotes may prefer less-developed areas, some coyotes may be forced to occupy developed areas due to territoriality. In order for coyotes to occupy these areas, there must be landscape components enabling them to exist. We found that human-associated food sources made up 2–11 % of the diet of coyotes in the study area, indicating that native foods dominated (89–98 %; Morey et al. 2007). Since the coyotes were mostly feeding on native prey, we concluded they did not prefer developed areas but would rather occupy areas with less development. The compositional analysis also showed a preference for native habitats, as well as less-developed areas, when coyotes are given a choice of developed land types versus less-developed land types in their home range.
Coyotes have been shown to adapt to urbanized areas by restricting activity to less-developed areas (Quinn 1997a; Gehrt et al. 2009). The higher use of less-developed land types and the lower use of urban land by all coyotes in our study area at the third order of selection showed the importance of less-developed areas. Even though coyotes in developed areas used urban land more than coyotes in less-developed areas, it was much less than was available in their home ranges and in the study area. This is further evidence that less-developed areas offered resources that were limited in urban land, although the high use of urban land by coyotes in developed areas also showed they used urban land at some point during their activities. This suggests that, because urban land is the dominant land-use type in the home ranges of coyotes in developed areas, less-developed land-use types may occur in patches and may be dispersed throughout their home ranges, thus requiring coyotes to travel through urban land to access these areas.
Urban coyotes have been found to be crepuscular or nocturnal (Quinn 1997a; Grinder and Krausman 2001a), and less active during diurnal periods (Shargo 1988; McClennen et al. 2001). Diurnal locations have also been associated with resting behavior, and nocturnal locations with traveling or hunting behavior (Andelt and Andelt 1981; Tigas et al. 2002). If landscape selection was different for each of these activities, we should have seen a different land-use selection between diel periods. We did not see a significant difference in land-use selection between diel periods for any group. The same land-use types that our coyotes used during diurnal times appear to also provide resources for their nocturnal activities. Also, the unchanged temporal selection for coyotes in less-developed areas showed that these coyotes were staying within boundaries of less-developed areas. Most of the less-developed areas were forest preserves having defined borders with very few fragments of developed land within. The lack of utilization of urban land during diel periods by these coyotes suggested they may have less contact with humans through both periods compared with coyotes in developed areas.
Coyotes have been found to use more developed or urban land at night (Quinn 1997a; Riley et al. 2002). Therefore, we thought the use of urban land by coyotes in developed areas would have increased at night when human activity decreased. The lack of change in utilization of urban land between diel periods for coyotes in our study area suggested either human activity continued to suppress coyote activity at night, or coyotes used urban land irrespective of human activity during both diel periods.
Coyotes in each of our groups had different access to nonurban habitats. By definition, coyotes in less-developed areas had higher percentages of nonurban habitats and lower percentages of urban habitats in their home ranges compared with developed areas. Nonurban habitats of shrubs, tall natural grasses, riparian vegetation, and woods can provide cover for coyotes. These areas also provided coyotes with their main food items including rodents, lagomorphs, deer, and vegetation (Morey et al. 2007). These resources were reduced in impervious surface habitats consisting of roads, parking lots, and buildings, and also in manicured grass habitats consisting of golf course fairways, and lawns in city parks and residential areas, suggesting less-developed areas might be superior to developed areas. In addition, in order for coyotes in developed areas to utilize needed nonurban habitats, they must have larger home ranges, travel through urban habitats, and potentially increase their exposure to humans.
Nonurban habitats (riparian, natural grass, crops, woods) were important for all coyotes in the study area. All 3 groups used nonurban habitats near or in greater proportions than their availability. This is particularly true for coyotes in developed areas that used riparian and natural grass areas in twice the proportion of their availability. Similar to land use, coyotes in developed areas switched their top-ranked habitats from urban habitats at the second order of selection to nonurban habitats at the third order. This again showed that, although their home ranges may contain high degrees of urbanization, coyotes in developed areas need some nonurban areas in their home ranges.
All coyotes showed strong avoidance of urban habitats. All coyotes used impervious surfaces and manicured grass in lower percentages than was available in their home range. This was particularly true for coyotes in developed areas, whose home ranges consisted of 48 % impervious surfaces, yet they only utilized 19 % of these areas. Some impervious surfaces may not be available to coyotes (i.e., although paved areas are available for use, buildings are not). Based on the lack of resources and cover on impervious surfaces, it was understandable why coyotes did not select these areas. Coyotes avoided manicured grass which lacked horizontal cover. Even impervious surface areas had buildings, fences, and other manmade barriers to provide some level of cover. However, manicured grass areas may have provided sources of food, as coyotes were observed stalking or chasing domestic cats, rabbits, and deer in these areas.
Natural areas consisting of grasslands, woods, and riparian communities were the most important habitats for coyotes in the study area. Grass and open areas have been found to be important habitats for coyotes (Andelt and Andelt 1981; Person and Hirth 1991; Holzman et al. 1992b). Natural grasslands, woods, and riparian habitats provide coyotes with cover and food. Rodents and lagomorphs were important food sources for coyotes in both developed and less-developed areas (Morey et al. 2007). The high utilization of these prey items by all coyotes in our study explains why they used natural grass areas in higher proportion than available in their home range. Analysis of food habits also showed that the availability of rodents and lagomorphs were similar in nonurban habitats in developed and less-developed areas (Morey et al. 2007).
Wooded areas have been used by coyotes for denning (Person and Hirth 1991) and cover (Andelt and Andelt 1981). Although we did not observe seasonal differences of wooded habitat utilization, wooded areas were important during denning activities; 15 of 19 coyote den sites found in our study area were within wooded habitats. One den was in a 0.5-ha patch of woods within a residential area. In addition to cover, food resources may have attracted coyotes to wooded areas. Rodents were the dominant food item found in scats in the study area (Morey et al. 2007).
The spatial and temporal variation of landscape utilization among coyotes suggested that different potentials for human conflict exist between developed and less-developed areas (Gehrt et al. 2009). Coyotes in developed areas had large home ranges, consisting of mostly urban land, yet preferred nonurban habitats for resting and foraging. Because of the fragmented landscape inherent to many urban areas, many of these nonurban habitats were dispersed throughout their home ranges. When urban land was avoided, coyotes in developed areas were forced to travel through these areas to access nonurban habitats. The ensuing use of urban land as travel paths could put coyotes in potential contact with humans and pets. However, coyotes in developed areas reduced their activity levels during nocturnal periods when human activity was high. Because of the use of urban habitats primarily as travel paths and the temporal reduction of movements during times with high human activity, coyotes appeared to minimize contact with humans. Additionally, coyotes in less-developed areas may have reduced potential for contact with humans, as these coyotes rarely went into urban land. Less-developed areas contained little urban land and more vertical cover than nonurban habitats, minimizing their exposure to humans. The high use of nonurban habitats and low use of urban habitats suggested these animals may behaviorally reduce potential contact with humans.
The spatial and temporal variation of landscape utilization among coyotes in our area showing a partitioning of subpopulations may help to understand coyote behavior in an urban landscape. We believe the approach of making a general inference about the population as a whole may mask important variations in patterns of landscape use. Our ability to classify coyotes based on their degree of association with urban land showed that coyotes in developed areas had larger home ranges and less access to natural habitats, and may have more exposure to humans, as compared with animals in less-developed areas. The interpretation of how coyotes use the urban landscape and the potential for contact with humans will depend on their classification into one of these landscape groups. Managers must recognize that a “one hat fits all” approach may be insufficient to properly manage urban coyote populations.
References
Aebischer NJ, Robertson PA (1992) Practical aspects of compositional analysis as applied to pheasant habitat utilization. In: Priede IG, Swift SM (eds) Wildlife telemetry: remote monitoring and tracking of animals. Ellis Horwood Limited, London, pp 285–293
Aebischer NJ, Robertson PA, Kenward RE (1993) Compositional analysis of habitat use from animal radio-tracking data. Ecology 74:1313–1325
Andelt WF (1985) Behavioral ecology of coyotes in south Texas. Wildl Monogr 94:1–45
Andelt WF, Andelt SH (1981) Habitat use by coyotes in southeastern Nebraska. J Wildl Manag 45:1001–1005
Andelt WF, Manan BR (1980) Behavior of an urban coyote. Am Midl Nat 103:399–400
Baker RO, Timm RM (1998) Management of conflicts between urban coyotes and humans in southern California. Vertebr Pest Conf 18:288–312
Carbyn LN (1989) Coyote attacks on children in western North America. Wildl Soc Bull 17:444–446
Carr GM, MacDonald DW (1986) The sociality of solitary foragers: a model based on resource dispersion. Anim Behav 35:1540–1549
Clark KA, Neill SU, Smith JS, Wilson PJ, Whadford VW, McKirahan GW (1994) Epizootic canine rabies transmitted by coyotes in south Texas. J Am Vet Med Assoc 204:536–540
Gehrt SD (2004) Ecology and management of striped skunks, raccoons, and coyotes in urban landscapes. In: Fascione N, Delach A, Smith M (eds) Predators and people: from conflict to conservation. Island, Washington, pp 81–104
Gehrt SD, Anchor C, White LA (2009) Home range and landscape use of coyotes in a metropolitan landscape: conflict or coexistence? J Mammal 90:1045–1057
Gese EM, Andersen DE, Rongstad OJ (1990) Determining home-range size of resident coyotes from point and sequential locations. J Wildl Manag 54:501–506
Gese EM, Rongstad OJ, Mytton WR (1988) Home range and habitat use of coyotes in southeastern Colorado. J Wildl Manag 52:640–646
Gier HT (1968) Coyotes in Kansas (revised). Kansas State College Agricultural Experiment Station Bulletin 393, Manhattan, Kansas
Grinder M (1999) Ecology of coyotes in Tucson, Arizona. Ph.D. dissertation, University of Arizona, Tucson
Grinder M, Krausman PR (2001a) Home range, habitat use, and nocturnal activity of coyotes in an urban environment. J Wildl Manag 65:887–898
Grinder M, Krausman PR (2001b) Morbidity–mortality factors and survival of an urban coyote population in Arizona. J Wildl Dis 37:312–317
Holzman S, Conroy MJ, Pickering J (1992a) Diseases, parasites and survival of coyotes in south-central Georgia. J Wildl Dis 28:572–580
Holzman S, Conroy MJ, Pickering J (1992b) Home range, movements, and habitat use of coyotes in southcentral Georgia. J Wildl Manag 56:139–146
Howell RG (1982) The urban coyote problem in Los Angeles County. Vertebrate Pest Conf 10:21–23
Johnson DH (1980) The comparison of usage and availability measurements for evaluation of resource preference. Ecology 61:65–71
Laundrè JW, Keller BL (1981) Home range use by coyotes in Idaho. Anim Behav 29:449–461
Leban F (1999) Resource selection for Windows, version 1.0. University of Idaho, Moscow
Litvaitis JA, Shaw JH (1980) Coyote movements, habitat use, and food habits in southwestern Oklahoma. J Wildl Manag 44:62–68
MacDonald DW (1983) The ecology of carnivore social behaviour. Nature 301:379–384
Martin D (1999) Wild (and unleashed) coyote is captured in Central Park. New York Times 148:51480, New York, pp B1
McClennen N, Wigglesworth RR, Anderson SH (2001) The effect of suburban and agricultural development on the activity patterns of coyotes (Canis latrans). Am Midl Nat 146:27–36
McClure MF, Smith NR, Shaw WW (1995) Diets of coyotes near the boundary of Saguaro National Monument and Tuscon, Arizona. Southwest Nat 40:101–125
McNab NK (1963) Bioenergetics and the determination of home range size. Am Nat 97:133–140
Meia JS, Weber JM (1995) Home ranges and movements of red foxes in central Europe: stability despite environmental changes. Can J Zool 73:1960–1966
Morey PS, Gese EM, Gehrt S (2007) Spatial and temporal variation in the diet of coyotes in the Chicago Metropolitan Area. Am Midl Nat 158:147–161
Odum EP, Kuenzler EJ (1955) Measurement of territory and home range size in birds. Auk 72:128–137
Person DK, Hirth DH (1991) Home range and habitat use of coyotes in farm region of Vermont. J Wildl Manag 55:433–441
Quinn T (1997a) Coyote (Canis latrans) habitat selection in urban areas of western Washington via analysis of routine movements. Northwest Sci 71:289–297
Quinn T (1997b) Coyote (Canis latrans) food habits in three urban habitats of western Washington. Northwest Sci 71:1–5
Reynolds TD, Laundrè JW (1990) Time intervals for estimating pronghorn and coyote home ranges and daily movements. J Wildl Manag 54:316–322
Riley SD, Sauvajot RM, Fuller TK, York EC, Kamradt DA, Bromley C, Wayne RK (2002) Effects of urbanization and habitat fragmentation on bobcats and coyotes in southern California. Conserv Biol 17:566–576
Rodgers AR, Carr AP (2002) ESRI ArcView home range extension. Ontario Ministry of Natural Resources, Toronto
Shargo ES (1988) Home range, movements, and activity patterns of coyotes (Canis latrans) in Los Angeles suburbs. Ph.D. dissertation, University of California, Los Angeles
Tigas LA, Van Vuren DH, Sauvajot RM (2002) Behavioral responses of bobcats and coyotes to habitat fragmentation and corridors in an urban environment. Biol Conserv 108:299–306
Timm RM, Baker RO, Bennett JR, Coolahan CC (2004) Coyote attacks: an increasing suburban problem. Trans North Am Wildl Nat Resour Conf 69:67–88
US Census Bureau (2000) State and metropolitan data book. Census Bureau, Washington
White GC, Garrott RA (1990) Analysis of wildlife radio-tracking data. Academic, New York
Worton BJ (1989) Kernal methods for estimating the utilization distribution in home range studie. Ecology 70:164–168
Acknowledgments
Funding and logistical support for this project were provided by Cook County Animal and Rabies Control, Max McGraw Wildlife Foundation, Forest Preserve District of Cook County, US Department of Agriculture’s National Wildlife Research Center-Logan Field Station, and Utah State University. We thank J.A. Bissonette, P. Box, and J. Symanzik for reviews of the manuscript. For assistance and support we thank B. Montgomery and C. Paine of the Max McGraw Wildlife Foundation, C. Anchor of the Forest Preserve District of Cook County, P.A. Terletzky and S. Durham of Utah State University, and D. Brooks of the Schaumburg Park District. We extend many thanks to technicians J. Rieb, C. King, C. Mullens, M. Heiser, D. Bogan, C. Diel, B. Fredrick, K. Freeman, M. Neri, T. Pruess, and G. Turcshak for field assistance.
Author information
Authors and Affiliations
Corresponding author
About this article
Cite this article
Gese, E.M., Morey, P.S. & Gehrt, S.D. Influence of the urban matrix on space use of coyotes in the Chicago metropolitan area. J Ethol 30, 413–425 (2012). https://doi.org/10.1007/s10164-012-0339-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10164-012-0339-8