
Abstract
Western scrub jays (Aphelocoma californica) hide food and rely on spatial memory to recover their caches at a later date. To do this cache-and-recovery, they can use both spatial and site-specific cues. I examined these cues in an experimental setting. The experiment established that scrub jays, like other food storers, prefer to rely on the location of the caching tray rather than tray-specific cues. They could modify their preference for spatial cues through training in which spatial cues were made irrelevant. Even after such training, the spatial cues controlled the jays’ behaviour when the spatial and site-specific cues gave competitive information about the cached sites. Thus, the global spatial cues have priority but the jays use the local site-specific cues when the spatial cues do not give enough information about the cached site.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
A number of studies have demonstrated that food-storing birds show impressive feats of spatial learning and memory. There is evidence from both the field and the laboratory that food-caching species encode the spatial location of their caches during a single, brief visit when the item is hidden and that they then rely, at least in part, on this memory when recovering their caches (see Shettleworth 1995 for a review). In corvids such as the Clark’s nutcracker (Nucifraga columbiana), observations of cache recovery in the wild suggest that an individual bird will recover about 72% of its caches during one field season (Tomback 1980). In the laboratory, the accuracy of cache recovery is impressive. Bednekoff and Balda (1997) found that Clark’s nutcrackers directed 82% of their ‘bill plunges’ at cache sites during recovery trials. Furthermore, when these birds were presented with a 2×3 array of potential caches sites, only one of which contained a food cache, they made an average of less than one ‘error’ (visit to a site that did not contain a food cache) before a successful recovery. Laboratory studies also demonstrate the longevity of these memories. Indeed some food-storing species show accurate retention of the spatial locations of a number of cache sites over long time periods, ranging from several weeks in some chickadees and titmice (Hitchcock and Sherry 1990; Healy and Suhonen 1996) to at least 285 days in Clark’s nutcrackers, N. columbiana, (Balda and Kamil 1992).
To achieve high accuracy of cache recovery using spatial memory, information about hundreds of individual cache sites has to be remembered over long time periods. As a consequence, the information may have to be remembered over different seasons, in which local colour, pattern, and shape cues in the environment may be altered dramatically. One might expect birds to rely on distal cues about the position of various prominent landmarks such as trees, rather than on local cues at the vicinity of the cache site (e.g. colour and shape of leaves, pattern and colour of the substrate) because the former are less likely to change over time and are more conspicuous from afar. Indeed, experiments with food-caching European jays (Garrulus glandarius) show that these birds rely on large vertical landmarks to provide cache-recovery cues (Bennett 1993). Clark’s nutcrackers also rely on large, prominent objects as landmarks to relocate food caches they have buried in the ground, whereas local cues provided by the soil substrate appear to be relatively unimportant (Vander Wall 1982). A predisposition to preferentially remember large vertical landmarks near caches, such as trees, has the advantage that these cues are unlikely to change from season to season, or to be covered by snow during the winter months (e.g. Bennett 1993).
In the field, the distinction between local and global cues is less clear because the extent to which a cue is site specific or global (spatial) is relative to the size of the area in which the animal is searching. Thus a particular object may provide both spatial and site-specific cues depending on the scale of the area searched. When a food-storing marsh tit, for example, flies across the woodland, a given oak tree may be viewed as a spatial cue. Yet the same tree might be viewed as a site-specific cue once the bird has perched on the tree and is searching for the location of a particular cache hidden under the bark. To cite Herz et al. (1994, p. 350), “It remains an open question whether the same mechanisms of spatial orientation are used on different spatial scales, or whether different mechanisms are employed depending on the scale of the area searched and the movements involved”.
Functionally, there may be good reason to attend to multiple landmarks, namely, because it increases the precision of searching at the target location, particularly when the goal and landmark are some distance apart. Kamil and Cheng (2001) proposed the “multiple-bearing hypothesis”: that food caching birds such as Clark’s nutcrackers may use multiple landmarks to work out their position and thus help them to relocate their caches of hidden food. The birds could do this by taking compass bearings to individual landmarks or by using the relative geometrical positions of the landmarks. Some evidence for the latter comes from a study by Kamil and Jones (1997) who tested Clark’s nutcrackers that had been trained to find a seed at the midpoint between two landmarks. The birds encoded the landmark orientation as middle on tests with novel distances between the landmarks, demonstrating their ability to relocate a point defined not by the relationship between the goal and the landmark, but by the relationship between landmarks.
In the laboratory, one can test whether animals might encode multiple landmarks using both spatial and site-specific cues by removing, adding, and rotating some of the cues after caching and prior to recovery. For example, Balda and Turek (1984) found that removal of prominent landmarks disrupts accurate cache recovery in Clark’s nutcrackers, and Bunch and Tomback (1986) found that the addition of prominent landmarks increased cache recovery accuracy in gray jays (Perisoreus canadensis). Herz et al. (1994) studied which spatial cues black-capped chickadees used for cache recovery by selectively removing or rotating various cues during recovery in the arena in which the birds had cached seeds. Removal of site-specific cues prior to cache recovery had no significant effect on the birds’ recovery accuracy, whereas removal of spatial cues significantly decreased recovery accuracy. The birds were then tested in a second experiment in which only the spatial cues were present and these were sometimes rotated by 90, 180, or 270° prior to recovery. When the spatial cues were rotated the birds searched at sites that were in the same position, relative to rotated distal objects (spatial cues), as the original cache sites had been. Taken together, the results of these experiments show that distal or spatial cues are more important than local cues for cache recovery in these species of food storer.
Here, I investigate the role of spatial and site-specific cues in spatial learning of western scrub jays. The western scrub jay is a food-storing bird like the Clark’s nutcracker and the pinyon jay. The Clark’s nutcracker has a sublingual pouch to hold seeds and the pinyon jay has an extended esophagus to hold seeds, whereas the western scrub jay does not have such morphological adaptations for food storing. Nonetheless, the western scrub jay stores around 6,000 pine seeds (Balda 1980). They also showed cache and recovery in a laboratory (Clayton and Dickinson 1998, 1999) and good spatial memory in a radial maze task (Gould-Beierle 2000). The purpose of the experiment was as follows: (a) to examine whether western scrub jays use predominantly spatial cues or site-specific cues to search for food stored by themselves, and (b) to examine the possibility of training birds to identify the tray in which they had cached using site-specific cues, independent of the position of the caching tray in the cage.
Materials and methods
Subjects
Six male and four female adult western scrub jays (Aphelecoma californica) were used in this study. These birds had experience of a cache-recovery accuracy test. In other words, they were adapted to cache and recovery in an experimental setting. Each bird was housed individually in a wire mesh cage (91×91×76 cm). The maintenance diet consisted of a variety of foods including powdered IAMS dog food and peanuts, a mixed seed mix, shelled sunflower seeds, pine nuts, mealworms, boiled egg, and grapes. Food and water were provided ad libitum when the birds were not being tested. During caching and recovery trials, which were conducted in the bird’s home cage, the maintenance diet was removed at 1730 hours on the previous day and returned to the bird’s home cage once the recovery trial was complete. Birds received these caching and recovery trials between 1030 hours and 1600 hours, and during these trials the birds could consume and cache wax worms, their most preferred food. The lights went off at 1800 hours and came on again at 0800 hours the following morning.
Apparatus
The birds cached and recovered food from plastic ice cube trays (6×22 cm) that contained a 2×8 array of 2.5-cm ice cube moulds, each of which was a potential cache site. The ice cube moulds were filled with corn kibble so that the birds could bury their caches. Each tray was secured to a wooden board (15×32 cm) and a heterogeneous arrangement of topographically and visually distinct Lego Duplo blocks was attached to one of the long sides of the tray to provide a distinct local, site-specific cue (see Clayton and Dickinson 1998). Birds therefore cached in visuo-spatially distinct trays on different days, and the tray location also varied from trial to trial, to ensure that each caching event was trial unique. The home cages were used as experimental chambers. In addition to perches inside the cages, all of the surrounding cages, walls, and a door of the animal room provided spatial cues.
Procedure
The jays received two habituation trials in which they could eat and cache wax worms and subsequently recover those they had cached in the caching tray approximately 4 h later. Each bird was tested individually. During the caching period, a caching tray was placed on the left or right side of the cage and a plastic bowl containing 50 wax worms was placed at the centre of the floor of the cage.
The subjects were allowed to eat and cache worms for 15 min, and then the tray was removed and the number of worms that had been cached in each ice cube mould was recorded. Prior to each recovery period the kibble was replaced and smoothed over to remove any local visual or olfactory cues about the location of cache. The tray was then returned to the original position in the cage after 4 h and the experimenter recorded by direct observation the number and location of searches made, and the number of caches recovered during the 5-min recovery period. After the first habituation trial, the next trial was carried out on the next day.
After 2 days of habituation trials, each bird received a competitive test (test 1) on the next day, which differed from the habituation trials during the recovery session in two respects. (1) All food was removed from the tray prior to recovery and fresh corn kibble was placed in the tray. This procedure was used to test the bird’s memory for the location of the previously hidden caches in the absence of any cues emanating directly from the food. (2) Two trays were present during recovery, such that the original tray in which the bird had cached (‘caching tray’) was placed in a new position in the cage and a new tray (‘new tray’) with a new site-specific cue (Lego Duplo) was placed in the location in which the bird had originally cached during the caching period. As the location and site-specific cues were contradictory, the test trial dissociated position and site-specific cues.
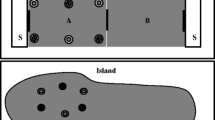
Following the first test trial (competitive test 1), all subjects received a block of six training trials (competitive training) in which the birds had the opportunity to learn to use the site-specific cues. One training trial was given per day. Thus, competitive training consisted of 6 days. During caching, the caching tray was placed at one of six different locations as shown in Fig. 1. The location of the tray was selected at random during the training trials, with the provision that each bird received a new tray in a different position during caching so that each bird cached in all six locations during the six training trials. The subjects were allowed to eat and cache worms for 15 min, then after 4 h they received a recovery period. During the 5-min recovery period, the birds received two trays. The original tray containing the worms they had cached there 4 h previously was now placed in a different position in the cage. A new tray was positioned where the original tray had been placed during the caching period. To recover their caches, the birds had to learn to avoid searching in the place where their caching tray had been located during the caching phase and to rely instead on the tray-specific Lego Duplo surround to guide their choice of searches during recovery.

The six possible locations for the caching tray. During a block of six training trials, the caching tray was placed in a new location on every caching trial. During recovery, the tray in which the bird had cached was moved to a second location and a new tray was placed in the caching tray’s previous location (competitive training) or a new position (non-competitive training)
After six training trials, the subjects received a second test trial (competitive test 2) that was identical to the first one, that is, in which no food was present at the time of recovery. The second block of six training trials (non-competitive training) began on the day after the second test trial. These trials were similar to the first block of training trials, in that birds cached in one tray and were then given the choice of two trays in which they could search during recovery. At recovery, the original caching tray was moved to a different position in the cage, as before. However, rather than placing the new tray in the location of the caching tray, for the second training block the new tray was placed in a new position that had not been occupied during caching. Twelve different trays were used (i.e. six caching trays and six additional trays for use during the recovery sessions), to ensure that on every trial the tray-specific Lego Duplo cues were novel. The trays were placed in the same six locations shown in Fig. 1, following the same procedures described for the previous block of training trials.
Birds received a third test on the day after the second block of training trials. This test trial (non-competitive test) differed from the previous test trials in that both trays were placed in new locations during recovery. So in this test trial, unlike in previous ones, the information about tray location and local Lego Duplo cues specific to the caching tray were not contradictory because neither tray was located in the position where the caching tray had been placed during the caching period. The purpose of this test trial was to establish whether the birds found it easier to use site-specific cues when no cache site was available in the original position so that the bird no longer had to choose between position and local site-specific cues. To maintain cache-recovery behaviour, the birds were given two non-competitive training trials before receiving a final test trial (competitive test 3) that was identical to the first two test trials.
Behavioural measures and analysis
The number and location of the food caches were recorded by determining which ice cube moulds contained caches at the end of each caching period. During training trials in which food was present during recovery I recorded the number of caches that were recovered and then calculated the number of caches recovered as a proportion of the number cached to take into account the fact that different individuals cache different amounts and on different trials. Across all trials, birds searched for food during recovery by probing the kibble and by repetitively swiping at the kibble with their bills in an apparent attempt to clear the kibble from the cache site. The total number and location of bill probes and bill swipes were recorded by direct observation, and specific ice-cube-mould sites inspected by the birds during recovery were then verified by noting the disturbances in the kibble in each cube mould. The first bill probe or swipe to a cache site counted as a search and a further search was recorded whenever a bird returned to that site after searching in another cache site.
Cache recovery efficiency was measured in two ways: first, by calculating the number of searches to sites in which the bird had cached as a proportion of the total number of searches during recovery, and second, by re-calculating these proportions based on only the first ten searches during recovery. Shettleworth and Krebs (1982) argued that the proportion of caches recovered during the first ten searches is considered to be a more accurate measure than using the total number of visits per trial. The reason is a motivational one: at the start of recovery, the hungry bird searches for the caches that it has hidden previously, whereas the bird is less hungry towards the end of the trial, having already eaten some of its recovered caches, and may therefore be more likely to explore new sites than to recover the remainder of its caches.
A one-sample t test was used to compare cache recovery accuracy and tray searching preferences with that predicted by random search. One-way analyses of variance (ANOVAs) with trial as the repeated measure were conducted on the number of items cached, the proportion of caches recovered, the proportion of searches directed to cache sites during training trials, and the proportion of searches in the caching tray during test trials. A binomial test was used to compare the number of birds making their first search to the caching and new trays.
Results
Caching behaviour
Birds readily cached and recovered from the trays, caching a mean of 12.3±5.47 (SD) items per trial during the first six trials of training and 6.05±2.95 (SD) during the second six trials of training. There was no significant effect of trial on the amount cached during the first six trials of training, F(5,59)=0.229, P=0.95, but there was a significant effect of trial during the second six trials of training, F(5,59)=2.55, P=0.04, and the difference between the first and second six trials was also significant (paired t test based on individual mean, t(9)=3.11, P=0.012). As shown in Fig. 2a this difference arose because there was a decrease in the number of items cached during the second six trials of training, and to some extent, on subsequent test trials.

Results of the two blocks of training trials. a Mean ± SE of the number of worms cached per trial. b Mean ± SE per trial of the number of worms that were recovered as a proportion of the number of worms cached. c Mean ± SE per trial for the number of searches at sites in which the bird had cached as a proportion of the total number of sites visited by the bird during recovery
Figure 3a shows the mean results of the four test trials. There was no significant effect of training on the number of items cached during the four test trials, F(3,39)=1.25, P=0.31, although the number of items cached across the non-competitive tests to competitive test 3 was consistently low.

Results of the test trials. a Mean ± SE of the number of worms cached per trial on test. b Total number of searches to the tray in which the bird had cached, expressed as a proportion of the total number of searches to both trays (solid bars) and during the first ten visits of a trial (hatched bars) for each of the four test trials in which birds cached as usual but no food was present during recovery. A score of 0.5 would therefore indicate no bias towards one of the two trays, and the higher the score the greater the preference for searching during recovery in the caching tray that had been placed in a new location. Competitive test 1 (C-TEST 1) was carried out prior to training, whereas competitive test 2 (C-TEST 2) was given after the six competitive training trials, and non-competitive test (NC-TEST) and competitive test 3 (C-TEST 3) after six non-competitive training trials
Recovery behaviour during training trials
During training, the birds recovered a mean of 5.73±2.74 (SD) items per trial, but it is clear from Fig. 2b and c that the proportion of items recovered increased across the two blocks of training, as did the proportion of searches to cache sites. Although both indices increased slightly during the first six training trials, a one-way ANOVA showed no significant effect of training on the proportion of items recovered, F(5,59)=0.19, P=0.97, and the proportion of searches to cache sites, F(5,59)=0.58, P=0.71. When the second block of six training trials (non-competitive training) was analysed, there was no significant effect of training on the proportion of items recovered, F(5,59)=0.623, P=0.683, but there was a significant effect of training on the proportion of searches to cache sites, F(5,59)=2.489, P=0.042. Generally, the birds recovered a greater proportion of their caches during the non-competitive training (paired t test, t(9)=2.68, P=0.025), and they were also more efficient at cache recovery, making fewer searches to find the caches (paired t test, t(9)=2.84, P=0.019).
Competitive tests
In Fig. 3b, data are presented in terms of the number of searches made to the caching tray during the recovery test. Data are presented in terms of the number of searches made to the caching tray as during the recovery test, and in terms of the first ten searches during recovery, expressed as a proportion of the total number of searches made to both trays. In the first competitive test, prior to any training, nine birds searched first in the new tray that had been placed in the original location of the caching tray, and only one bird searched in the caching tray that had been placed in a new location. This result suggests that these birds have a strong initial preference for using position cues over local tray-specific cues when searching for their caches (binominal test, P<0.02). A one-sample t test showed a significant difference from chance in the proportion of searches to the caching tray using both the total number of searches, t(9)=−3.64, P=0.005, and the first ten visits, t= −3.91, P=0.004.
As shown in Fig. 3b, the preference for tray location was reduced in subsequent test trials (competitive tests 2 and 3). In contrast to the first test where only one bird visited the caching tray placed in the new location (“correct tray”) first, six of ten birds visited the correct tray first in the competitive test 2, and five of ten birds visited the correct tray first in the competitive test 3. So in contrast to the first test, the number of birds visiting the caching tray did not differ from that expected by random in either of the two subsequent trial tests (binomial test, P=0.67 for competitive test 2 and P=0.62 for competitive test 3). A paired t test gave a significant difference between the competitive tests 1 and 2 on the proportion of total searches (t=3.317, df=9, P=0.009) and the first ten searches (t=3.96, P=0.003). However, the birds did not show a significant preference for relying on site-specific cues. Indeed, the proportion of searches to the correct tray did not differ significantly from chance, t(9)=0.715, P=0.49 and t(9)=1.168, P=0.272, based on the total number of searches and the first ten searches, respectively.
Similarly in the final competitive test, there was no significant preference for using the site-specific cues and the proportion of searches to the correct tray did not differ from chance (one group t test, t(9)=0.276, P=0.789 and t=0.509, P=0.623 for total and first ten searches, respectively). It is also clear from Fig. 3b that the results of competitive tests 2 and 3 are similar and a paired t test gave no significant difference in the proportion of searches to the correct tray using either the total number of searches, t(9)=0.57, P=0.58, or the first ten searches, t(9)=0.82, P=0.43. Like the results of competitive test 2, the proportion of searches in competitive test 3 was significantly different from that for competitive test 1, t(9)=2.679, P=0.025 and t(9)=2.682, P=0.025 for total and first ten searches, respectively.
Non-competitive test
In the non-competitive test trial, where the new tray was not placed in the caching tray’s original position, seven birds visited the correct tray first, although a binomial test did not give a significant difference from chance (binomial test, P=0.17). There was a significant difference from chance in the proportion of searches to the correct tray, however (one group t test, t=2.744, df=9, P=0.023 for the total number of searches and t=3.50, P=0.007 for the first ten searches). There was also a significant difference between the competitive test 1 and non-competitive test (t=4.769, df=9, P=0.001 and t=5.722, P=0.0003 for total and first ten searches, respectively). There was no significant difference between the competitive test 2 and the non-competitive test, t(9)=0.25 and 0.82, P=0.81 and 0.43 for total and first ten searches, respectively.
Discussion
In summary, the jays showed a strong initial preference to use spatial cues in competitive tests, although this strategy could be modified through training, and there was no statistically robust preference for relying on site-specific cues in any of the competitive test trials. When there was no alternative tray placed at the original location (non-competitive test), however, then the jays could clearly identify the tray based on its site-specific cues. Thus the preference for relying on site-specific cues was manifest only in the absence of conflicting spatial cues. The birds in this study showed a strong preference to rely on spatial cues when recovering their caches, but they were also able to use site-specific cues such as the Lego Duplo blocks surrounding each caching tray, provided no spatial cues were present to provide conflicting information.
Although the birds did not prefer the site-specific cues to identify the cached tray, they might use the site-specific cues to identify position of the cached moulds on the tray. Thus, their recovery behaviour has two processes, namely, identification of the cached tray by spatial cue, then identification of the cached cells in the tray by site-specific (tray-specific) cue.
Other studies have shown that food storers show a strong preference to rely on spatial cues about the position of various landmarks and cache sites rather than non-spatial (e.g. colour or pattern) cues to identify a particular cache site (Bennett 1993). Some researchers have used a one-trial associative memory procedure to address this issue (Brodbeck 1994; Clayton and Krebs 1994). The procedure consisted of two phases. Birds had to return in phase II of a trial to sites where they had been allowed to eat some, but not all, of a piece of peanut in phase I. If the correct choice in phase I is a compound stimulus of spatial location and non-spatial (e.g. colour or pattern) cues surrounding the baited site, then food-storing species respond primarily to the spatial information, whereas non-storers respond equally to spatial and non-spatial cues. This result has been shown in comparative studies of food-storing and non-storing species of parids and corvids (Brodbeck 1994; Clayton and Krebs 1994). These comparative studies demonstrated that food-storing birds preferentially visited the correct site on the search in phase II, whereas non-storers choose both position and site equally in the first and second phases.
Gould-Beirle and Kamil (1998) compared hidden-food searching behaviour of Clark’s nutcracker, pinyon jay, and western scrub jay and found that all species use local (landmark) cues to search for the hidden food. The birds were trained to find food hidden by an experimenter, not stored by the subjects. The size of the experimental arena and a tray for search were large (room 440×270 cm and tray 120×120 cm). In fact, the size of the tray was larger than the experimental chamber in the present experiment. If the size of the searching area is taken into acount, the landmark in Gould-Beirle and Kamil’s experiment would be a spatial cue in the present experiment. Thus, the present results were not contradictory to those found by Gould-Beirle and Kamil.
These observations and the current study confirm spatial-cue-based memory in the western scrub jays. Even though they have a strong trait for spatial-cue-based learning, they could modify their learning strategy under some conditions. In other words, their spatial learning is not completely determined by their natural trait but is also controlled by experience to some degree.
References
Balda RP (1980) Are seed caching systems co-evolved? Acta Congr Int Ornithol 2:1185–1191
Balda RP, Kamil AC (1992) Long-term spatial memeory in Clark’s nutcrackers, Nucifraga columbiana. Anim Behav 44:761–769
Balda RP, Turek RJ (1984) The cache-recovery system as an example of memory capabilities in the Clark’s nutcracker. In: Roitblat HL, Bever TG, Terrace HS (eds) Animal cognition. Erlbaum, Hillsdale, NJ, pp 513–532
Bednekoff PA, Balda RP (1997) Clark’s nutcracker spatial memory: many errors might not be due to forgetting. Anim Behav 54:691–698
Bennett ATD (1993) Spatial memory in a food storing corvid I. Near tall landmarks are primarily used. J Comp Physiol A 173:193–207
Brodbeck DR (1994) Memory for spatial and local cues: a comparison of a storing and a non-storing species. Anim Learn Behav 22:119–133
Bunch KG, Tomback DF (1986) Bolus recovery by gray jays: an experimental analysis. Anim Behav 34:754–762
Clayton NS, Dickinson A (1998) Episodic-like memory during cache recovery by scrub jays. Nature 395:272–278
Clayton NS, Dickinson A (1999) Scrub jays (Aphelocoma coerulescens) remember the relative time of caching as well as the location and content of their caches. J Comp Psychol 113:403–416
Clayton NS, Krebs JR (1994) Memory for spatial and object-specific cues in food-storing and non-storing birds. J Comp Physiol A 174:371–379
Gould-Beierle KL (2000) A comparison of four corvid species in a working and reference memory task using a radial maze. J Comp Psychol 114:347–356
Gould-Beierle KL, Kamil AC (1998) Use of landmarks in three species of food-storing corvids. Ethology 104:361–378
Healy SD, Suhonen J (1996) Memory for locations of stored food in willow tits and marsh tits. Behaviour 133:71–80
Herz RS, Zanette L, Sherry DF (1994) Spatial cues for cache retrieval by black-capped chickadees. Anim Behav 48:343–351
Hitchcock CL, Sherry DF (1990) Long-term memory for cache sites in the black-capped chickadee. Anim Behav 40:701–712
Kamil AC, Cheng K (2001) Way-finding and landmarks: the multiple-bearings hypothesis. J Exp Biol 204: 101–113
Kamil AC, Jones JE (1997) The seed-storing corvid Clark’s nutcracker learns geometric relationships among landmarks. Nature 390:276–279
Shettleworth SJ (1995) Memory in food-storing birds: from the field to the Skinner box. In: Alleva E, Fasolo A, Lipp H-P, Nadel L (eds) Proceedings of NATO advanced study institute series Maratea, Italy. Kluwer, The Hague, pp 158–179
Shettleworth SJ, Krebs JR (1982) How marsh tits find their hoards: roles of site preferences and spatial memory. J Exp Psychol Anim Behav Proc 8: 354–375
Tomback DF (1980) How nutcrackers find their stores. Condor 82:10–19
Vander Wall SB (1982) An experimental analysis of cache recovery in Clark’s nutcracker. Anim Behav 30:84–94
Acknowledgements
The experiment was carried out in the department of experimental psychology, University of Cambridge. The author wants to express his gratitude to Dr. N. Clayton and Dr. T. Dickenson for their advice and comments on the experiments and a draft of this paper. This research was supported by The 21st Century COE Program (D-1) in Japan.
Author information
Authors and Affiliations
Corresponding author
About this article
Cite this article
Watanabe, S. Strategies of spatial learning for food storing in scrub jays. J Ethol 23, 181–187 (2005). https://doi.org/10.1007/s10164-005-0150-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10164-005-0150-x