
Abstract
Morphological variation among natural populations is a phenomenon commonly observed in marine invertebrates and well studied, particularly, in shelled gastropods. The nassariid Buccinanops globulosus is interesting to study shell shape variation because it exhibits strong interpopulation differences in life history features, including maximum size, fecundity and growth rate. In this study, we examined the pattern of variation in size and shell shape among populations and between sexes of B. globulosus (Bahía San Antonio 40°29′S 63°01′W, Playa Villarino 40°45′S 64°40′W and Bahía Nueva 42°46′S 65°02′W). In particular, we used geometric morphometric techniques to test: (1) whether the two components of shell morphology (size and shape) are independent and (2) whether shape differences between sexes are consistently found among populations, regardless of their body sizes. Our results show shell shape variation between the populations of B. globulosus of northern Patagonia. Intra-specific shell shape variation is affected by body size, indicating allometry. Regardless of the size differences, individuals from Playa Villarino have high-spired shells, and shorter apertures and wider columellar area than individuals from the other populations. Also, sex-related shape differences were consistently found at each population, thus suggesting a common sexual dimorphism in shell morphology for this species. The functional significance of the variability found is discussed in terms of the flexibility of developmental programmes for morphology as well as the evolution of phenotypic plasticity.
Similar content being viewed by others


Introduction
Morphological variation among natural populations is a phenomenon frequently observed in marine invertebrates and well studied, particularly, in shelled gastropods. The study of shell shape is important for the identification of species and for the quantification of morphological variation within species (e.g. Grahame and Mill 1989; De Wolf et al. 1998; Rolán et al. 2004; Conde-Padín et al. 2007; Sánchez et al. 2011). Because a shell contains a complete record of ontogeny for its owner, differences in shell shape may represent variations in shared developmental programmes (Stone 1998). Shelled gastropods provide an ideal system for the study of potential trade-offs associated with induced changes in shell shape because they must live within the shell they construct (Trussell 2000). Also, the collection of morphometric data may offer a cost-effective alternative to infer key biological parameters for individual populations because body form is a product of ontogeny (Saunders and Mayfield 2008; Walker and Grahame 2011).
Shell shape variation within species may occur both at a microgeographical scale (i.e. over distances of metres or between different shore levels at the same site) and at a macrogeographical scale (i.e. between distant regions). The ecological process of natural selection may lead, among others, to differences in shell morphology between populations. The factors in the environment that exert selection, both the biological (predators, competitors and parasites) and the non-biological ones (physical factors, for example, temperature regimes), may vary along the geographical distribution of a species. When the characteristics under selection show heredity, selection will lead to evolutionary change in these characteristics (Kingsolver and Pfenning 2007). Intra-specific shell shape variation, for example, among littorinids, has been well documented (Reid 1996; Johannesson 2003; Rolán-Alvarez 2007). In Littorina saxatilis, two distinct morphological forms or ecotypes occur along Galician, British and Swedish shores (Johannesson et al. 1993; Hull et al. 1996; Hollander et al. 2005, 2006; Conde-Padín et al. 2007). This polymorphism, at least in Galician shores, is explained mainly by natural selection acting directly on the genes affecting the morphological traits (Conde-Padín et al. 2009). Another way of modifying phenotype to increase adaptation induced by the environment may be mediated by phenotypic plasticity, that is, differential expression of genotype in different environments. This has been reported in many species, for example, the dogwhelk Nucella lapillus in the presence of crab effluents develops thicker lip and teeth inside the aperture (Appleton and Palmer 1988; Palmer 1990). Other source of variability in shell morphology may be related to the presence of allometric patterns. Allometry, the variation of traits associated with variation of the overall size of organisms (Huxley and Teissier 1936) is interesting not only because of its implications for form–function relationships but also because of the insight into growth and development (Zelditch et al. 2004).
The purpose of morphometrics is to quantify, analyse and describe variation in biological forms (Roth and Mercer 2000). Many techniques have been used for the morphological study of gastropod shells: classic measurements analysed in a multivariate framework, mathematical modelling (e.g. Raup 1966; Stone 1995; Urdy et al. 2010a) and, more recently, geometric morphometric techniques (Bookstein et al. 1985). Geometric morphometric techniques present many advantages, including the possibility to separate the size and shape components of morphology, the graphic display of results so they can be interpreted easily, the maintenance of geometric relationships throughout the analysis and a remarkable statistical power (Bookstein et al. 1985; Rohlf and Slice 1990; Klingenberg 2002; Adams et al. 2004). There is a variety of geometric morphometric methods that may be applied according to the material under study. Regarding shells, it is possible to recognize discrete anatomical loci, named landmarks, which can be found repeatedly and reliably in all specimens (Zelditch et al. 2004). In several studies dealing with marine gastropods, landmark-based geometric morphometrics has been recently used to describe shape differences between ecotypes (e.g. Carvajal-Rodríguez et al. 2005, 2006; Hollander et al. 2006; Conde-Padín et al. 2007, 2009; Guerra-Varela et al. 2009), to identify species (Urra et al. 2007) and to assess pollution effects (Márquez et al. 2011).
Buccinanops globulosus is interesting to study shell shape variation because it exhibits strong interpopulation variation in life history features, including maximum size, fecundity and growth rate (Narvarte et al. 2008; Avaca et al. 2013). This intertidal species occurs in soft substrates (mud or sand), along the Atlantic coast of South America (34°S to 48°S) (Scarabino 1977). It feeds on carrion, mainly carcasses of crabs (Daleo et al. 2005) or bivalves (e.g. Amiantis purpurata, Avaca et al. 2010). Development is direct; females enclose their eggs and nurse eggs in capsules attached to their callus shell zone. Juveniles of about 5 mm hatch from the capsules after 2–3 months of intracapsular development (Narvarte 2006; Avaca et al. 2012). Thus, the potential for dispersal is thought to be low as in other species with similar life history.
Adult shells of B. globulosus are longer than wide, while those of juveniles that recently emerged from the egg capsules are almost isodiametric (pers. obs.). Hence, the post-hatching growth rates of different parts of the shell must differ (i.e. allometric growth) to give place to adult proportions. However, allometric growth patterns have been described for only one species of the family Nassariidae, Nassarius vibex (Yokoyama and Amaral 2011). Allometric growth patterns coupled with interpopulation size variation can result in morphological variation (Zelditch et al. 2004). Furthermore, recent theoretical models have suggested that variation in growth rates can interact with allometric growth patterns to produce different shell shapes (Urdy et al. 2010b). Several studies have shown a relationship between shell shape and growth rates in snails (e.g. Kemp and Bertness 1984; Chapman 1997; Urdy et al. 2010b; Saura et al. 2012); even slightly allometric growth patterns can result in demonstrably different adult shell shapes if growth rates are very dissimilar (Tamburi and Martín 2009). Interpopulation variation in the shell growth rate in B. globulosus is substantial (Narvarte et al. 2008; Avaca et al. 2013) and is therefore a feasible explanation for the different shell shapes found in North Patagonia. Females of B. globulosus also grow faster and attain larger sizes than males (Narvarte et al. 2008; Avaca et al. 2013), and this could add another component to the morphological variation in these populations.
In this study, we examine the pattern of variation in size and shell shape among populations and between sexes of a small intertidal gastropod. An important reason for choosing B. globulosus is that the differences in growth rates both between populations and between sexes may lead to allometric variation. In particular, we used geometric morphometric techniques to test: (1) whether there is independence between the two components of shell morphology, that is, size and shape and (2) whether shape differences between sexes are consistently found among populations regardless of their body sizes.
Materials and methods
Study sites and sample collection
The study was conducted in San Matías Gulf and Nuevo Gulf (northern Patagonia, Argentina). Specimens of B. globulosus were monthly collected during 2006–2007 from three populations for which other life history features are known (Narvarte 2006; Narvarte et al. 2008; Avaca et al. 2013). Two of the sampling sites were located in San Matías Gulf (Bahía San Antonio [BSA] 40°29′S 63°01′W and Playa Villarino [PV]: 40°45′S 64°40′W) and one in Nuevo Gulf (Bahía Nueva [BN] 42°46′S 65°02′W) (Fig. 1). Sampling sites are separated from each other along the South Atlantic Ocean coast by at least 50 km, and the two most distant sites, PV and BN, are 300 km apart (Fig. 1). These populations were chosen because they represent the near extremes in shell size documented for B. globulosus in Patagonia (Narvarte et al. 2008).

Location of the sampling sites in San Matías Gulf (SMG) and Nuevo Gulf (NG) in northern Patagonia. 1: Playa Villarino (PV), 2: Bahía San Antonio (BSA) and 3: Bahía Nueva (BN)
Biological bait, mainly composed of discarded fish, was used to attract whelks on sandy bottoms on the lower intertidal and upper infralittoral zone (Narvarte 2006). Bait was left for at least 20 min on each sampling occasion; attracted snails were collected, stored and carried to the laboratory. In the laboratory, individuals were frozen in order to be sexed, when defrosted, without damaging the shell. Due to the occurrence of imposex in two of the populations studied, BSA and BN (Bigatti et al. 2009), sex was determined on the basis of the presence of an albumen gland in females and by its absence and the presence of a penis in males. Total shell length was measured with a digital calliper (±0.01 mm). To reduce the effects of ontogenetic allometry, only adult individuals were used. Adult size was established separately for each population, according to female size at maturity, that is, size of the smallest female with egg capsules on the callus, although male maturity may be attained at a smaller size than that of females (BSA 15.92 mm, PV 45.2 mm and BN 23.17 mm; Narvarte et al. 2008; Avaca et al. 2012).
Geometric morphometrics
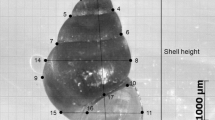
Shells were digitally imaged in apertural view and were always placed in a consistent orientation, that is, with the shell axis parallel to the horizontal plane, and following the recommendations of Zelditch et al. (2004). We used the tpsUtil and tpsDig software (Rohlf 2006a) to place landmarks on shell images. A total of 13 landmarks (LM) along the perimeter of the shell were placed to capture the shell form (Fig. 2). LM1 was placed at the apex of the shell; LM 2 was on the right edge of the outline of the shell at the end of the lower suture of the penultimate whorl; LM 3 was at the suture of the last whorl; LM 4 was at the beginning of the aperture, LM 5 was at the midpoint of the outer lip; LM 6 was at the end of the outer lip; LM 7 was at the end of the siphonal canal; LM 8 was at beginning of the carina of the siphonal canal; LM 9 was at the most external point of the body whorl; LM 10 was opposite to LM 2; LM 11 was the beginning of the siphonal canal; LM 12 and LM 13 were, respectively, on the internal and external edge of the columellar callus on a perpendicular line to the axis of LM 5.

Shells of B. globulosus from different sites in northern Patagonia: Playa Villarino (left), Bahía Nueva (centre) and Bahía San Antonio (right), showing the 13 landmarks selected for this study. Scale bar: 1 cm
For each shell, shape and size variables were obtained by using the tpsRelw software (Rohlf 2006b). Shape variables were estimated by the alignment of the raw coordinates of the specimens by the Generalized Procrustes Analysis (Rohlf and Slice 1990; Bookstein 1991; Rohlf and Marcus 1993). In geometric morphometrics, shape is defined as all the geometric information that remains when location, scale and rotational effects are removed from an object (Kendall 1977). Aligned configurations were used to calculate partial warps and uniform component scores, and then to perform a relative warp analysis. Partial warp scores represent the non-uniform part of the deformation, describing changes that have a location and spatial extent in the organism (Zelditch et al. 2004). The combination of the uniform and non-uniform components describes any shape change. The relative warp (RW) analysis consists in fitting an interpolating function (thin-plate spline) to all homologous landmarks or each specimen in a sample (Rohlf 1993). The RWs were computed with the scaling option α = 0. Each RW explains a percentage of the overall local variation in decreasing order, as with principal components (Rohlf 1993). The centroid size was computed as the square root of the sum of squared distances of the series of measured landmarks to their common centroid (Bookstein 1991). Centroid size is an estimator of size uncorrelated with shape in the absence of allometry (Zelditch et al. 2004).
Firstly, we analysed differences in size and shape between populations, and then, each population was analysed separately to investigate shape differences between sexes. Size variation was assessed with two-way ANOVA with population (BSA, BN and PV) and sex (male and female) as random and fixed factors, respectively. Data were Ln-transformed to fit for assumptions of normality. To assess for allometric effects, both between and within populations, we performed a multivariate regression of shape on size using Procrustes coordinates and centroid size (Ln), as dependent and independent variables, respectively (Klingenberg 2011). Owing to the presence of allometry (see results), the residual values of the multivariate regression were used as new shape variables independent of the size effect, that is, the non-allometric shape component. To test for differences in the non-allometric shape component, both between populations and between sexes, we used multivariate ANOVA (MANOVA: Zelditch et al. 2004). We also performed a canonical variate analysis (CVA) coupled with discriminant analysis to ordinate the samples by maximizing the separation of the specified groups (populations or sexes) (Zelditch et al. 2004). All multivariate analyses, except MANOVA, were computed with the software MorphoJ version 1.05a (Klingenberg 2011). Shape differences along the first CV axis both between and within populations were described using thin-plate spline deformation grids by using the TPSRegr software (Rohlf 2007a, b).
Results
A total of 607 shells from the three populations were analysed. The size range (mm) and the number (n) of shells per population were as follows: BSA 16–28 mm, n = 194 (n M 98, n F 96) PV 45–62 mm, n = 177 (n M 68, n F 109) and BN 25–42 mm, n = 236 (n M 114, n F 122).
Size and shape variation between populations
Both population and sex effects on size were found to be significant (two-way ANOVA, Table 1). The mean size of the PV population was larger than that of BN and BSA (LSD Fisher; P < 0.05). Except for BSA, females were significantly larger than males of each population (Tukey’s HSD, P < 0.05) (Fig. 3), although the overall interaction between site and sex was not significant.

Mean centroid size of B. globulosus shells from northern Patagonia, compared by sex (M males, F females) and population (PV Playa Villarino, BN Bahía Nueva and BSA Bahía San Antonio). Different letters indicate that the means are different under the post hoc comparisons (Tukey’s HSD test, P < 0.05)
The first eight RWs explained 87.91 % of the total variance. Since significant differences in size were found between populations and the scatter plot of RW 1 (42.51 % of the total variation) versus Ln (CS) suggested a linear relationship between size and shape (Fig. 4), we computed the pooled-within-group multivariate regression of shape (Procrustes coordinates) on size. This regression was highly significant thus indicating the presence of allometry (P < 0.0001) (Table 2). The percentage of shape variation explained by size was 10.54 %.

Scatter plot of centroid size (Ln-transformed) and first relative warp (RW1) estimate of shell shape for B. globulosus populations from northern Patagonia. PV Playa Villarino, BN Bahía Nueva and BSA Bahía San Antonio
The MANOVA on the residual values of the multivariate regression (non-allometric component of shape variation) comparing populations was highly significant (Wilk’s λ = 0.12; F 44, 1116 = 50.89; P < 0.0001). This indicates that differences in shell shape between populations persisted when size effects were removed.
The canonical variate analysis on the residuals resulted in a clear separation of the individuals in the morphospace (Fig. 5). The first canonical axis explained 89.02 % of between-population shell shape variation. This axis depicts mostly a contrasting shape pattern between Playa Villarino (negative scores) and the other two sites (positive scores). The Mahalanobis distances showed that the highest similarity was between Bahía San Antonio and Bahía Nueva; nevertheless, each site presented a typical shell shape (Mahalanobis distance pairwise comparisons: BSA-PV 4.93, BSA-BN 1.91 and PV-BN 4.32; for each permutation test P < 0.0001). The percentage of individuals correctly allocated into their population group varied between 74.12 and 100 according to the cross-validation method.

Canonical variate analysis of the non-allometric shell shape variation between populations of B. globulosus populations from northern Patagonia (PV Playa Villarino, BN Bahía Nueva and BSA Bahía San Antonio)
Differences between populations represented in the CV1 by the thin-plate spline deformation grids involved the height of the spire, the size of the aperture and the columellar zone (Fig. 6). Individuals from Playa Villarino showed relatively higher spires, shorter apertures and a wider columellar area, which describe a more elongate shell shape than that of individuals from the other populations. In contrast, individuals from Bahía San Antonio presented lower spires, larger apertures and narrower collumelar area, resulting in a more globose shell shape.

Thin-plate spline representation along the first CV axis of the non-allometric component of shape variation for each population of B. globulosus from northern Patagonia (PV Playa Villarino, BN Bahía Nueva and BSA Bahía San Antonio), showing the mean canonical score obtained. Some landmarks are connected by lines to facilitate the interpretation of differences between populations
Shape variation within populations
The multivariate pooled-within-group regression of Procrustes coordinates on centroid size (Ln) was found to be highly significant for each population, thus indicating allometry (Table 2). The percentage of shape variation explained by size was 8.90, 5.15 and 28.39 % for BSA, PV and BN, respectively.
The MANOVA on the residual values of the multivariate regression (non-allometric component of shape variation) using sex as factor was highly significant for each population (BSA: Wilk’s λ = 0.47, F 22, 171 = 8.68; PV: Wilk’s λ = 0.40, F 22, 154 = 10.42 and BN: Wilk’s λ = 0.46, F 22, 213 = 11.29, P < 0.0001), indicating sexual dimorphism in shell shape.
The canonical variate analysis on the residuals showed a distinction between sexes within each population (Fig. 7). Differences in shell shape between males and females were significant within each population (Mahalanobis pairwise distance; BSA 2.12, PV 2.49, BN 2.15; for each permutation test P < 0.0001). The percentage of individuals correctly allocated into their sex group varied between 79.60 and 89.30 according to the cross-validation method. The efficiency of discrimination between sexes was similar between populations (Fig. 7). Differences between sexes represented in the CV1 depicted by the thin-plate spline deformation grids are related to the compression of the columellar axis and involve total width and size of the aperture (Fig. 8). Males have more elongated shells and more slender apertures than females.

Canonical variate analysis between sexes within each population of B. globulosus from northern Patagonia (PV Playa Villarino, BN Bahía Nueva and BSA: Bahía San Antonio). The numbers indicate the percentage of individuals correctly allocated into their sex group according to the discriminant function obtained

Thin-plate spline representation showing the most extreme canonical scores obtained along the first CV of the non-allometric component of shape variation for males and females from each population. PV Playa Villarino, BN Bahía Nueva and BSA Bahía San Antonio
Discussion
Shell shape variation was found between the populations of B. globulosus of northern Patagonia. Our results show that intra-specific shell shape variation is affected by body size, indicating allometry. Regardless of the size differences, the individuals from Playa Villarino showed high-spired shells, and shorter apertures and wider columellar area than individuals from the other populations. Also, sex-related shape differences were consistently found at each population, thus suggesting a common sexual dimorphism in shell morphology.
Geometric morphometrics was useful both to describe and to quantify the shell shape variation observed between populations of B. globulosus. This method was reliable for distinguishing individuals from different locations based solely on their shell shape (successful diagnosis of 89 % on average). Centroid size was effective to characterize populations, being this the first report of its use in this species. This estimate of size may be used as a size variable in any study; for example, the combined use of centroid size and the age of individuals can be used to assess the individual growth pattern (Vioarsdóttir et al. 2002). However, this use is not common among invertebrate species.
The size-related variation in shape observed, although small, means that growth in size is accompanied by changes in shell shape. This is the first report for the genus Buccinanops about the presence of an allometric growth pattern and the second in a nassariid species (Yokoyama and Amaral 2011). Allometric changes are usually related to the need of maintaining the functionality of the organisms not only for biomechanical reasons but also for facing the different ecological demands that vary over the individual lifespan (Zelditch et al. 2004). For example, allometric growth at the size of maturity in the E-morph of L. saxatilis in Swedish shores is consistent with promoting increased fecundity by expanding spire volume and giving room to brood more embryos (Hollander et al. 2006). Thus, shell shape variation may be related to fitness variables, for example, reproductive effort (Hollander et al. 2006; Walker and Grahame 2011). In this sense, females of B. globulosus from Playa Villarino, which show larger shells and wider columellar area, are also more fecund in terms of number of egg capsules per egg mass than females from the other two study sites (Narvarte et al. 2008; Avaca et al. 2012). However, to what extent shell shape and reproductive output are related should be further established.
Sexual dimorphism in the shell shape of B. globulosus involves the aperture area and total width. The aperture has been widely mentioned as the most labile feature of whelk shells, being the area where differences may be related to sexual function (ten Hallers-Tjabbes 1979). Males of B. globulosus have more slender apertures than females, being the first report of shell dimorphism for this species. In other species of marine gastropods, males have larger apertures than females (e.g. Buccinum undatum: ten Hallers-Tjabbes 1979; Gendron 1992; Kideys et al. 1993). Among females, the response to organotin compounds may also cause differences in shell morphology (size and/or shape) between females affected and unaffected by imposex (e.g. Son and Hughes 2000; Bigatti and Carranza 2007; Márquez et al. 2011). However, in this study, shell shape differences between sexes were similar between organotin-polluted (Bahía San Antonio and Bahía Nueva) and organotin-unpolluted sites (Playa Villarino), thus suggesting little or negligible effects of this phenomenon in this species at these sites and at low intensity imposex level (e.g. females affected by imposex normally carry egg capsules, Avaca et al. 2012).
The intertidal zone not only represents a highly variable environment with clines of food availability, desiccation and presence of predators but it is also affected by human impact (Raffaelli and Hawkins 1999; Chapman 2000). Intra-specific variation in shell morphology may be driven by several ecological factors other than body size, such as predation cues, wave exposure or temperature regime (e.g. Trussell and Etter 2001; Dalziel and Boulding 2005; Doyle et al. 2010). A paradigm of rocky intertidal ecology is that gastropods from wave-exposed shores tend to be small with thin shells and larger apertures, whereas conspecifics from wave-sheltered shores tend to be large with thick shells and smaller apertures (Raffaelli and Hawkins 1999). Larger apertures allow keeping a larger foot, which in turn allow decreasing the risk of dislodgment (Grahame and Mill 1986; Trussell 1997). The large size and thick shell with its small aperture of sheltered shore gastropods are thought to be adaptations to the high abundance of highly motile predators, to crushing by stones, and to desiccation and heat stress (Palmer 1979; Edgell et al. 2008). In this study, which was performed on sandy beaches, the fact that individuals from both bays (Bahía San Antonio and Bahía Nueva) were more similar in shape than individuals from Playa Villarino, a long dissipative beach, suggests that factors leading to the variation observed may be related to environmental conditions. The lack of a planktonic larval stage and the burrowing habit of this scavenger snail (Avaca et al. 2012) indicates that the dispersal rate is probably low, and hence the probability of adaptation to local conditions is high, at least at a mesogeographical scale. On the other hand, size-corrected shape differences between Playa Villarino and the other two populations may be the result of the mechanical interplay between allometric growth patterns and shell growth rates (Kemp and Bertness 1984; Chapman 1997; Urdy et al. 2010b; Saura et al. 2012), since the overall growth performance of this population is higher than that of the other two populations of B. globulosus (Narvarte 2006; Narvarte et al. 2008; Avaca et al. 2013). Unfortunately, ecological studies on B. globulosus are scarce (Daleo et al. 2005, 2012; Narvarte et al. 2008) and there are many gaps regarding trophic relationships or survival rate at different environments. Probably the factors affecting size and shell shape variation in B. globulosus are multiple and not mutually exclusive. For example, temperature and predator cues explain differences in shell form and body mass in Littorina obtusata (Trussell 2000). Experimental studies are needed to address the role of ecological and physical factors related to the variation observed as well as to test whether this variation has an adaptive value.
The morphological variation found in the radula (Avaca et al. 2010) was different from that found in the shell (this study) of B. globulosus. The intra-specific variation in radular characteristics is related mainly to body size differences between populations. The radula of small body-sized individuals is essentially a scaled-down replica of the radula of large body-sized individuals. Regarding the shell, size differences explain only a small amount of the variation observed. The shell of small body-sized individuals presented larger apertures and shorter spires than the shell of large body-sized individuals. These differences between the morphological patterns of the shell and the radula may be related to the fact that shells are related to many biological functions (protection, growth and defence), whereas the radula is related only with an alimentary function. Therefore, it is likely that the number of factors related to the shell development is higher than that related to the radular development. Although it is not known whether variability in the traits examined has functional significance, it is still important to document such morphological variability. Some species are very stereotypic, while others are more “developmental noise” in their morphological construction. Understanding which species and under which conditions species fall into either camp will help us to understand the flexibility of developmental programmes for morphology as well as the evolution of phenotypic plasticity.
References
Adams DC, Rohlf FJ, Slice D (2004) Geometric morphometrics: ten years of progress following the ‘revolution’. Ital J Zool 71:5–16
Appleton RD, Palmer AR (1988) Water-borne stimuli released by predatory crabs and damaged prey induce more predator-resistant shells in a marine gastropod. Proc Natl Acad Sci USA 85:4387–4391
Avaca MS, Narvarte MA, Martín PR (2010) Description of the radula of Buccinanops globulosus (Neogastropoda: Nassariidae) and an assessment of its variability among Northern Patagonian Gulfs. Malacologia 53(1):175–184
Avaca MS, Narvarte MA, Martín PR (2012) Size assortative mating and effect of female size on reproductive output of the nassariid Buccinanops globulosus. J Sea Res 69:16–22
Avaca MS, Narvarte MA, Martín PR (2013) Age, growth and mortality in Buccinanops globulosus (Gastropoda: Nassariidae) from Golfo Nuevo (Argentina). Mar Biol Res 9:208–219
Bigatti G, Carranza A (2007) Phenotypic variability associated with the occurrence of imposex in Odontocymbiola magellanica from Golfo Nuevo, Patagonia. J Mar Biol Assoc UK 87:755–759
Bigatti G, Primost MA, Cledón M, Averbuj A, Theobald N, Gerwinski W, Arntz W, Morriconi E, Penchaszadeh PE (2009) Contamination by TBT and imposex biomonitoring along 4700 km of Argentinean shoreline (SW Atlantic: from 38°S to 54°S). Mar Poll Bull 58:695–701
Bookstein FL (1991) Morphometric tools for landmark data. Cambridge University Press, New York
Bookstein FL, Chernoff B, Elder RL, Humphries JM, Smith GR, Strauss RE (1985) Morphometrics in evolutionary biology. Academy of Natural Sciences Press, Philadelphia
Carvajal-Rodríguez A, Conde-Padín P, Rolán-Alvarez E (2005) Decomposing shell form into size and shape by geometric morphometric methods in two sympatric ecotypes of Littorina saxatilis. J Moll Stud 71:313–318
Carvajal-Rodríguez A, Guerra-Varela J, Fernández B, Rolán-Alvarez E (2006) An example of the application of geometric morphometric tools to the morphological diagnosis of two sibling species in Nassarius (Mollusca, Prosobranchia). Iberus 24:81–88
Chapman MG (1997) Relationships between shell shape, water reserves, survival and growth of highshore littorinids under experimental conditions in New South Wales, Australia. J Moll Stud 63:511–529
Chapman MG (2000) Variability of foraging in highshore habitats: dealing with unpredictability. Hydrobiologia 426:75–87
Conde-Padín P, Grahame JW, Rolán-Alvarez E (2007) Detecting shape differences in species of the Littorina saxatilis complex by morphometric analysis. J Moll Stud 73:147–154
Conde-Padín P, Caballero A, Rolán-Alvarez E (2009) Relative role of genetic determination and plastic response during ontogeny for shell-shape traits subjected to diversifying selection. Evolution 63:1356–1363
Daleo P, Escapa M, Isacch JP, Ribeiro P, Iribarne O (2005) Trophic facilitation by the oystercatcher Haematopus palliates Temminick on the scavenger snail Buccinanops globulosum Kiener in a Patagonian bay. J Exp Mar Biol Ecol 325:27–34
Daleo P, Alberti J, Avaca MS, Narvarte MA, Martinetto P, Iribarne O (2012) Avoidance of feeding opportunities by the whelk Buccinanops globulosum in the presence of damaged conspecifics. Mar Biol 159:2359–2365
Dalziel B, Boulding EG (2005) Water-borne cues from a shell-crushing predator induce a more massive shell in experimental populations of an intertidal snail. J Exp Mar Biol Ecol 317:25–35
De Wolf H, Backeljau T, Van Dongen S, Verhagen R (1998) Large-scale patterns of shell variation in Littorina striata, a planktonic developing periwinkle from Macaronesia (Mollusca: Prosobranchia). Mar Biol 131:309–317
Doyle S, Mcdonald BM, Rochette R (2010) Is water temperature responsible for geographic variation in shell mass of Littorina obtusata (L.) snails in the Gulf of Maine? J Exp Mar Biol Ecol 394:98–104
Edgell TC, Brazeau C, Grahame JW, Rochette R (2008) Simultaneous defense against shell entry and shell crushing in a snail faced with the predatory shorecrab Carcinus maenas. Mar Ecol Prog Ser 317:191–198
Gendron L (1992) Determination of the size at sexual maturity of the waved whelk Buccinum undatum Linnaeus, 1758, in the Gulf of St. Lawrence, as a basis for the establishment of a minimum catchable size. J Shell Res 11:1–7
Grahame J, Mill PJ (1986) Relative size of the foot of two species of Littorina on a rocky shore in Wales. J Zool 208:229–236
Grahame JW, Mill PJ (1989) Shell shape variation in Littorina saxatilis and L. arcana: a case of character displacement? J Mar Biol Assoc UK 69:837–855
Guerra-Varela J, Colson I, Backeljau T, Breugelmans K, Hughes RN, Rolán-Alvarez E (2009) The evolutionary mechanism maintaining shell shape and molecular differentiation between two ecotypes of the dogwhelk Nucella lapillus. Evol Ecol 23:261–280
Hollander J, Lindegarth M, Johannesson K (2005) Local adaptation but not geographical separation promotes assortative mating in a snail. Anim Behav 70:1029–1219
Hollander J, Adams DC, Johannesson K (2006) Evolution of adaptation through allometric shifts in a marine snail. Evolution 60:2490–2497
Hull SL, Grahame J, Mill PJ (1996) Morphological divergence and evidence for reproductive isolation in Littorina saxatilis (Olivi) in northeast England. J Moll Stud 62:89–99
Huxley JS, Teissier G (1936) Terminology of relative growth. Nature 137:780–781
Johannesson K (2003) Evolution in Littorina: ecology matters. J Sea Res 49:107–117
Johannesson K, Johannesson B, Rolán-Alvarez E (1993) Morphological differentiation and genetic cohesiveness over a microenvironmental gradient in the marine snail, Littorina saxatilis. Evolution 47:1770–1787
Kemp P, Bertness MD (1984) Snail shape and growth rates: evidence for plastic shell allometry in Littorina littorea. Proc Natl Acad Scienc USA 81:811–813
Kendall D (1977) The diffusion of shape. Adv Appl Probab 9:428–430
Kideys AE, Nash RDM, Hartnoll RG (1993) Reproductive cycle and energetic cost of reproduction of the neogastropod Buccinum undatum in the Irish Sea. J Mar Biol Assoc UK 73:391–403
Kingsolver JG, Pfenning DW (2007) Patterns and power of phenotypic selection in nature. Bioscience 57:561–572
Klingenberg CP (2002) Morphometrics and the role of the phenotype in studies of the evolution of developmental mechanisms. Gene 287:3–10
Klingenberg CP (2011) MORPHOJ: an integrated software package for geometric morphometrics. Mol Ecol 11:353–357
Márquez F, González-José R, Bigatti G (2011) Combined methods to detect pollution effects on shell shape and structure in Neogastropods. Ecol Indic 11:248–254
Narvarte MA (2006) Biology and fishery of the whelk Buccinanops globulosum (Kiener, 1834) in northern coastal waters of the San Matías Gulf (Patagonia, Argentina). Fish Res 77:131–137
Narvarte MA, Willers V, Avaca MS, Echave ME (2008) Population structure of the snail Buccinanops globulosum (Prosobranchia, Nassariidae) in San Matías Gulf, Patagonia Argentina: isolated enclaves? J Sea Res 60:144–150
Palmer AR (1979) Fish predation and the evolution of gastropod shell sculpture: experimental and geographical evidence. Evolution 33:697–713
Palmer AR (1990) Effect of crab effluent and scent of damaged conspecifics on feeding, growth, and shell morphology of the Atlantic dogwhelk Nucella lapillus (L.). Hydrobiologia 193:155–182
Raffaelli D, Hawkins S (1999) Intertidal ecology. Kluwer, The Netherlands
Raup DM (1966) Geometric analysis of shell coiling: general problems. J Paleontol 40:1178–1190
Reid DG (1996) Systematics and evolution of Littorina. The Ray Society, London
Rohlf FJ (1993) Relative warp analysis and an example of its application to mosquito wings. In: Marcus LF, Bello E, García Valdecasas A (eds) Contributions to morphometrics. Museo Nacional de Ciencias Naturales, Madrid, pp 131-159
Rohlf FJ (2006a) TPSDig2, version 2.1. Department of Ecology and Evolution, State University of New York at Stony Brook, New York
Rohlf FJ (2006b) TPSUtil, file utility program, version 1.38. Department of Ecology and Evolution, State University of New York at Stony Brook, New York
Rohlf FJ (2007a) TPSRegr, shape regression, version 1.34. Department of Ecology and Evolution, State University of New York at Stony Brook, New York
Rohlf FJ (2007b) TPSRelw, relative warp analysis, version 1.45. Department of Ecology and Evolution, State University of New York at Stony Brook, New York
Rohlf FJ, Marcus LF (1993) A revolution in morphometrics. Trends Ecol Evol 8:129–132
Rohlf FJ, Slice D (1990) Extensions of the Procrustes method for the optimal superimposition of landmarks. Syst Zool 39:40–59
Rolán E, Guerra-Varela J, Colson I, Hughes RN, Rolán-Alvarez E (2004) Morphological and genetic analysis of two sympatric morphs of the dogwhelk Nucella lapillus (Gastropoda: Muricidae) from Galicia (northwestern Spain). J Moll Stud 70:179–185
Rolán-Alvarez E (2007) Sympatric speciation as a by-product of ecological adaptation in the galician Lttorina saxatilis hybrid zone. J Moll Stud 73:1–10
Roth VL, Mercer JM (2000) Morphometrics in development and evolution. Am Zool 40:801–810
Sánchez R, Sepúlveda RD, Antonio B, Cárdenas L (2011) Spatial pattern of genetic and morphological diversity in the direct developer Acanthina monodon (Gastropoda: Mollusca). Mar Ecol Prog Ser 434:121–131
Saunders TM, Mayfield S (2008) Predicting biological variation using a simple morphometric marker in the sedentary marine invertebrate Haliotis rubra. Mar Ecol Prog Ser 366:75–89
Saura M, Rivas MJ, Diz AP, Caballero A, Rolán-Alvarez E (2012) Dietary effects on shell growth and shape in an intertidal marine snail, Littorina saxatilis. J Moll Stud. doi:10.1093/mollus/eys004
Scarabino V (1977) Moluscos del Golfo San Matías (Provincia de Río Negro, República Argentina). Inventario y claves para su identificación. Comun Soc Malacol Urug IV(31–32):177–297
Son MH, Hughes RN (2000) Relationship between imposex and morphological variation of the shell in Nucella lapillus (Gastropoda: Thaididae). Estuar Coast Shelf Sci 50:599–606
Stone JR (1995) CerioShell: a computer program designed to simulate variation in shell form. Paleobiology 21:509–519
Stone JR (1998) Landmark based thin plate spline relative warp analysis of gastropod shells. Syst Biol 47:254–263
Tamburi N, Martín PR (2009) Effect of food availability on morphometric and somatic indices of the apple snail Pomacea canaliculata (Caenogastropoda: Ampullaridae). Malacologia 51(2):221–232
ten Hallers-Tjabbes CC (1979) Sexual dimorphism in Buccinum undatum L. Malacologia 18:13–17
Trussell GC (1997) Phenotypic plasticity in the foot size of an intertidal snail. Ecology 78:1033–1048
Trussell GC (2000) Phenotypic clines, plasticity, and morphological trade-offs in an intertidal snail. Evolution 54:151–166
Trussell GC, Etter RJ (2001) Integrating genetic and environmental forces that shape the evolution of geographic variation in a marine snail. Genetica 112–113:321–337
Urdy S, Goudemand N, Bucher H, Chirat R (2010a) Allometries and the morphogenesis of the molluscan shell: a quantitative and theoretical model. J Exp Zool (Mol Dev Evol) 314B:280–302
Urdy S, Goudemand N, Bucher H, Chirat R (2010b) Growth-dependent phenotypic variation of molluscan shells: implications for allometric data interpretation. J Exp Zool (Mol Dev Evol) 314B:303–326
Urra A, Oliva D, Sepúlveda M (2007) Use of a morphometric analysis to differentiate Adelomelon ancilla and Odontocymbiola magellanica (Caenogastropoda: Volutidae) of Southern Chile. Zool Stud 46:253–261
Vioarsdóttir US, O’Higgins P, Stringer C (2002) A geometric morphometric study of regional differences in the ontogeny of the modern human facial skeleton. J Anat 201:211–229
Walker TN, Grahame JW (2011) Shell shape variation and fitness variables in the gastropod Littorina saxatilis. Mar Ecol Prog Ser 430:103–111
Yokoyama LQ, Amaral ACZ (2011) Allometric growth of a common Nassariidae (Gastropoda) in south-east Brazil. J Mar Biol Assoc UK 91:1095–1105
Zelditch M, Swiderski DL, Sheets HD, Fink W (2004) Morphometrics for biologists. A primer. Elsevier, London
Acknowledgments
We would like to thank “Kelo” Camarero for the technical support in the field. M.S.A is a postdoctoral fellow in CONICET. This work was partially supported by Project PID #371 ANPCYT to R. González, M. Narvarte and A. Gagliardini.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Franke.
Rights and permissions
About this article
Cite this article
Avaca, M.S., Narvarte, M., Martín, P. et al. Shell shape variation in the Nassariid Buccinanops globulosus in northern Patagonia. Helgol Mar Res 67, 567–577 (2013). https://doi.org/10.1007/s10152-013-0344-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10152-013-0344-5