
Abstract
Charophytes are a highly endangered group of algae. In the Baltic Sea, the number of species, distribution area and biomass of charophytes have significantly decreased in recent decades. Although eutrophication triggers their initial decline, the mechanism of the final extinction of charophyte populations is not fully understood. An in situ experiment was performed to study the role of the mesoherbivores Idotea baltica, Gammarus oceanicus and Palaemon adspersus in the decline of charophytes in the north-eastern Baltic Sea. Invertebrate grazing showed a clear seasonality: grazing pressure was low in April, moderate in July, and high in October. Grazing on charophytes by P. adspersus was negligible, whereas I. baltica and G. oceanicus significantly reduced the biomass of charophytes in the field. Low photosynthetic activity (high decomposition rate) of the charophytes favoured grazing. The invertebrates studied preferred Chara tomentosa to C. connivens. Low consumption of C. connivens may reflect its non-native origin. The experiment suggests that, under moderately eutrophic conditions, grazers are not likely to control charophyte populations. However, grazers have the potential to eliminate charophytes in severely eutrophic systems under the stress of filamentous algae.
Similar content being viewed by others

Introduction
Charophytes are a highly developed and diverse group of algae. They are widely distributed in freshwater, brackish and marine habitats from tropical to polar regions (Wood and Imahori 1965). Of 314 species, only six have been reported from all continents. Recent species prefer freshwater environments while the majority of fossil species occurred in brackish waters: even marine species were quite common. Charophytes often reach deeper than submerged angiosperms yet can also prevail in shallow waters (Chambers and Kalff 1985). In the Baltic Sea this group of algae inhabits sheltered coastal areas where their distribution pattern is primarily controlled by exposure, sediment type and salinity regime (Schubert and Yousef 2001a; Torn et al. 2004).
In recent decades, number of species, distribution area and biomass of charophytes have significantly declined in virtually the whole Baltic Sea (Dekere 2001; Koistinen and Munsterhjelm 2001; Martin 2001; Schubert and Yousef 2001b; Sinkevièienë and Jurgilaitë 2001). This decline has been attributed to increased nutrient loads resulting in higher productivity of phytoplankton, epiphytic algae and angiosperms. The reduction in available light intensity leads to an upward shift of the depth limit of charophytes and the replacement of the charophyte-dominated community by phanerogams and filamentous algae (Moss 1989; Scheffer at al. 1992; Yousef et al. 1997; Schubert and Yousef 2001b). The results, however, cannot explain the extinction of charophytes in shallower waters but this knowledge is essential for the effective protection of this highly endangered group of algae (HELCOM 1998).
Invertebrate herbivory is considered to be a major factor determining the structure and development of macroalgal assemblages (e.g. Paine 1974; Lubchenco 1978, 1982; Hawkins and Hartnoll 1983; Brawley 1992; Vadas and Elner 1992). As the mass development of epiphytic algae triggers an increase in grazer density (Kangas et al. 1982; Salemaa 1987; Malm 1999; Kotta 2000; Kotta et al. 2000), it is likely that the shading effect of filamentous algae, combined with an increased grazing rate, may be involved in the decline of charophytes in the Baltic Sea.
At present there are no experimental studies dealing with the effect of grazers on charophytes in the Baltic Sea. In our study, in situ grazing by prevailing mesoherbivores on two charophyte species was estimated in Kõiguste Bay, in the northern part of the Gulf of Riga. The main questions were: (1) Do selected invertebrates graze on Chara connivens and C. tomentosa?; (2) What quantities of alga are removed by grazing during different seasons?; and (3) Is the photosynthetic activity of the algae related to grazing pressure?
Methods
In situ grazing experiments were performed in Kõiguste Bay (58°22.10′N 22°58.69′E), north-eastern Baltic Sea in April, July and October 2001 (Table 1). Kõiguste Bay is typified by a wide coastal zone with a diverse bottom topography and macrophyte community. Boulders predominate in areas shallower than 3 m, pebbles at 3–8 m depth, and clay bottoms mixed with gravel and pebbles in deeper areas. Depending on the water exchange between the Gulf of Riga and the open Baltic Sea, salinity in Kõiguste Bay ranges from 6 to 7 psu. The region is ranked among the less eutrophic ones in the Estonian coastal sea (Suursaar 1995; Astok et al. 1999).
The globally spread C. tomentosa L. occurs in brackish waters only in the Baltic Sea (Olsen 1944; Björkman 1947) where it is distributed along virtually all coasts which provide wind- and wave-sheltered localities. The species is more frequent in the northern and central Baltic Sea (Pankow et al. 1990; Nielsen et al. 1995). In Europe, C. connivens Salzm. has been recorded from Scandinavia to Portugal and also in a few scattered localities in Britain and Ireland (Corrillion 1957; Moore 1986). In the Baltic Sea, C. connivens Salzm. is rare (Olsen 1944; Trei 1991; Nielsen at al. 1995) and considered as non-indigenous to the area (Luther 1979). According to recent data, the species is found only in Estonian and Swedish waters (Blindow 2000; Torn et al. 2004).
The prevailing grazers Idotea baltica (Pallas) and Gammarus oceanicus Segerstråle were collected from a shallow (1–3 m) area within the stands of Fucus vesiculosus L. by shaking the algae. Palaemon adspersus (Rathke) were caught by dredging the vegetated areas. Only adult specimens were used in the experiment.
I. baltica is omnivorous, feeding on benthic microalgae, filamentous algae, macroalgae, detritus, small invertebrates and even its conspecifics (Naylor 1955; Sywula 1964; Ravanko 1969; Nicotry 1980; Robertson and Mann 1980; Franke and Janke 1998). Gammarids are considered to be selective omnivores. Their diet consists of decaying organic matter with its microbial community, macroalgae, but also other animals such as other invertebrates, fish eggs, wounded fish (e.g. Macneil et al. 1997). Palaemon spp. are omnivorous, feeding on algae, moss, debris and small arthropods (Berglund 1980). Other studies have suggested the prevalence of carnivorous habits (Sitts and Knight 1979; Siegfried 1982).
Grazing was studied in 5×20 cm nylon net bags (~400 cm3) of 1 mm mesh size. To each macroalgal treatment was added either two specimens of I. baltica, two specimens of G. oceanicus or one specimen of P. adspersus. Net bags without grazers served as control. Three replicates of each treatment were run. The wet weight of algae was determined prior to the experiment to the nearest 0.01 g. Before weighing, the algae were gently dried on blotting paper until the paper did not become wet anymore. An additional three replicates of each macroalgal treatment served as controls to obtain the ratio of wet to dry weight. The algae were dried at 60°C for 48 h.
The net bags were placed at 2 m depth about 0.5 m above the bottom. Each series of the experiment lasted 10 days. In parallel to the grazing experiments, the in situ diurnal primary production of the studied macroalgal species was measured. Small tufts (ca. 0.05 g dry weight) with no macroepiphytes and grazers were placed in 600 ml glass bottles, filled with seawater and incubated horizontally on special trays at 0.5 m depth. Bottles which did not include any algae served as controls. There were five replicates per treatment and five controls (Kotta et al. 2000; Paalme et al. 2002).
At the end of the experiment the test animals were counted and the dry weights of invertebrates and macroalgae were determined for each net bag. The changes in the dry weight of algae relative to dry weight of invertebrate served as an estimate of invertebrate grazing in the field. The control treatment was used to compensate the grazing values with respect to algal production and/or decomposition.
Results
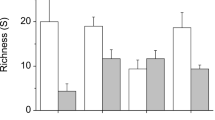
Invertebrate grazing on charophytes showed a clear seasonality (Table 2; Fig. 1). The effect depended on both algal and invertebrate species. The grazing of P. adspersus on charophytes was negligible, whereas I. baltica and G. oceanicus significantly reduced the biomass of charophytes in the field.

Seasonal variation in invertebrate grazing on charophyte species (means and SE)
Charophytes were not consumed in April. In July, grazing was moderate and only C. tomentosa was consumed. I. baltica had significantly higher grazing rates than G. salinus in July. Grazing was highest in October. The consumption of C. tomentosa by G. oceanicus was then about ten times as high as in July. Similarly, idoteid grazing on C. tomentosa was higher, though the difference was not statistically significant. In October, C. connivens was grazed by both I. baltica and G. oceanicus. The grazing pressure of the studied invertebrates on C. connivens was moderate and their values were not statistically different.
There was a negative linear relationship between the photosynthetic activity of C. tomentosa and invertebrate grazing, provided that water temperature was above 10°C (Figs. 2, 3). According to visual observations, the photosynthetic activity of the charophyte was inversely related to the state of algal decomposition.

Seasonal variation in algal production (means and SE)

Relationships between algal production and grazing rate. The zero grazing values are excluded from the analysis
Discussion
Grazing on charophytes in April was probably limited by low temperature. The significance of temperature for the activity of grazers has been demonstrated earlier in the field (Valiela et al. 1997; Lotze and Worm 2002). The activity of herbivores is negligible when water temperature is below 10°C in the north-eastern Baltic Sea (Orav-Kotta and Kotta 2003b). Hence, the recruitment of charophytes is likely to be affected more by climate (e.g. temperature, solar radiation) than by grazers.
The low grazing pressure in summer may be explained by the good condition of the charophytes as is indicated by their high photosynthetic activity. The protection against grazers may involve either morphological resistance (Steneck and Watling 1982; Hay et al. 1994) or chemical defence (Hay and Fenical 1988; Hay et al. 1994; Amsler 2001; Paul et al. 2001). The results of this experiment suggest that C. connivens is more resistant to herbivores than C. tomentosa. Besides, the mass development of filamentous alga which are the preferred food for the studied grazers in the area (Orav-Kotta and Kotta 2003a, 2003b) may explain why the grazing rate on charophytes is low in summer.
Our data suggest that herbivores have the highest impact on Chara stands in autumn. A similar seasonal pattern of herbivory has been reported for freshwater habitats, though waterbirds were considered in these experiments (Van den Berg 1999; Noordhuis et al. 2002). Visual observations indicate that strong grazing pressure is related to the onset of algal decomposition. This is also indicated by an inverse relationship between algal photosynthetic activity and invertebrate grazing.
By October, the density of filamentous algae had notably declined in the study area. The abundant population of herbivores which had relied on these algae were forced to switch to an alternative diet. As compared to other macroalgae in the area, the decomposing charophytes seemed to be the most rewarding food for the studied invertebrates, especially for G. oceanicus (Orav-Kotta and Kotta 2003b). The consumption of C. tomentosa by I. baltica was significantly higher than consumption of F. vesiculosus (Orav-Kotta and Kotta 2003b), even though the brown alga is ranked after the filamentous Pilayella littoralis as the second most important diet for this isopod in the northern Baltic Sea (Orav-Kotta and Kotta 2003a).
The utilisation of macrophytes is dependent on plant condition and nutritional quality (Hutchinson 1975; Wetzel 1983). As shown in this study, and also for other brackish (Paalme et al. 2002) and freshwater environments (Pöckl 1995), gammarids prefer decaying food to fresh algae. The low levels of herbivory are related to the unpalatability or resistance of the algae. According to Hunter (1976), fresh Chara are heavily calcified, which may greatly reduce their appeal to herbivores (but see Noordhuis et al. 2002). In the course of decomposition the cell walls of the algae become less resistant to herbivory (Birch et al. 1983) and the concentration of nutrients increases in the decomposing material as a result of increased microbial activity (Boyd 1970; Hunter 1976; Byren and Davies 1986; Buchsbaum et al. 1991). As a consequence, the algae become more attractive to benthic invertebrates (Mann 1988). The relationship between the rate of algal decomposition and grazing was not so straightforward for I. baltica, probably due to the different feeding mode of the isopod (e.g. Salemaa 1987). We observed that I. baltica was able to consume fresh alga. However, when provided with algae in different state of decomposition, I. baltica feeds selectively on more degraded parts of the algae (Salemaa 1987; Paalme et al. 2002). As for P. adspersus, this experiment demonstrated that charophytes (even when partly decomposed) do not belong to the diet of this species. The results support earlier findings of the prevalence of carnivorous habits in P. adspersus in the north-eastern Baltic Sea. In the absence of prey, P. adspersus may feed on filamentous algae such as Pilayella littoralis and Cladophora glomerata (L.) Kütz (Orav-Kotta and Kotta 2003b).
Why did the studied invertebrates prefer the coarse C. tomentosa over the fine C. connivens? It has been suggested that C. connivens has been introduced to western Europe and the Baltic Sea as an aquatic ballast plant. The known ranges coincide with port areas where, in earlier times, ballast sand was unloaded (Luther 1979). Hence, one possible explanation for low grazing pressure by I. baltica and G. oceanicus on C. connivens is that the native grazers may not be well adapted to feed on the introduced charophyte species. Only in October, when C. connivens had low photosynthetic activity and began to degrade, moderate grazing was observed. Often, the success of an introduction event is related to the absence of natural enemies (i.e. grazers or predators) in the recipient region (e.g. Moyle and Light 1996; Williamson and Fitter 1996). Consequently, the establishment of C. connivens in the Baltic Sea was probably favoured by an effective vegetative reproduction of the species (Luther 1979) and the absence of grazing during the productive season.
The relationship between photosynthetic activity of the charophytes and grazing may explain the disappearance of the charophyte populations. Under moderately eutrophic conditions, the charophyte populations are unlikely to be controlled by mesoherbivores. The grazers may partly reduce the vegetative production of the charophytes through an effective removal of algal biomass in autumn. However, in severely eutrophic conditions, when covered by filamentous algae, the photosynthetic activity of charophytes decreases and, hence, the algae become more attractive to herbivores. The high numbers of mesograzers in eutrophic areas (Kangas et al. 1982; Kotta et al. 2000) may then eliminate the charophytes. The grazers are likely to affect C. tomentosa more than C. connivens. We suggest that this difference reflects the non-native nature of C. connivens.
References
Amsler CD (2001) Induced defences in macroalgae: the herbivore makes difference. J Phycol 37:353–356
Astok V, Otsmann M, Suursaar Ü (1999) Water exchange as the main physical process in semi-enclosed marine systems: the Gulf of Riga case. Hydrobiologia 393:11–18
Berglund A (1980) Niche differentiation between two littoral prawns in Gullmar Fjord, Sweden: Palaemon adspersus and P. squilla. Holarctic Ecol 3:111–115
Birch PB, Gabrielson JO, Hamel KS (1983) Decomposition of Cladophora I. Field studies in the Peel-Harvey estuarine system, western Australia. Bot Mar 26:165–171
Björkman SO (1947) On the distribution of Chara tomentosa L. round the Baltic and some remarks on its specific epithet. Bot Not 2:157–170
Blindow I (2000) Distribution of charophytes along the Swedish coast in relation to salinity and eutrophication. Int Rev Hydrobiol 85:707–717
Boyd CE (1970) Losses of mineral nutrients during decomposition of Typha latifolia. Arch Hydrobiol 66:511–517
Brawley SH (1992) Mesoherbivores. In: John DM, Hawkins SJ, Price JH (eds) Plant-animal interactions in the marine benthos. The Systematics Association special vol No. 46. Clarendon Press, Oxford, pp 235–263
Buchsbaum R, Valiela I, Swain T, Dzierzeski M, Allen S (1991) Available and refractory nitrogen in detritus of coastal vascular plants and macroalgae. Mar Ecol Prog Ser 72:131–143
Byren BA, Davies BR (1986) The influence of invertebrates on the breakdown of Potamogeton pectinatus L. in a coastal marina (Zandvlei, South Africa). Hydrobiologia 137:141–151
Chambers PA, Kalff J (1985) Depth distribution and biomass of submerged aquatic macrophyte communities in relation to Secchi depth. Can J Fish Aquat Sci 42:701–709
Corrillion R (1957) Les Charophycées de France et d’Europe Occidentale. Bull Soc Sci Bretagne 11–12:1–499
Dekere Z (2001) Charophytes along the Lativan coast of the Baltic Sea. Schriftenr Landschaftspflege Naturschutz 72:19–21
Franke H-D, Janke M (1998) Mechanisms and consequences of intra- and interspecific interference competition in Idotea baltica (Pallas) and Idotea emarginata (Fabricius) (Crustacea: Isopoda): a laboratory study of possible proximate causes of habitat segregation. J Exp Mar Biol Ecol 227:1–21
Hawkins SJ, Hartnoll RG (1983) Grazing of intertidal algae by marine invertebrates. Oceanogr Mar Biol Annu Rev 21:195–282
Hay ME, Fenical W (1988) Marine plant-herbivore interactions: the ecology of chemical defence. Annu Rev Ecol Syst 19:11–145
Hay ME, Kappel QE, Fenical W (1994) Synergisms in plant defences against herbivores: interaction of chemistry, calcification and plant quality. Ecology 75:1714–1726
HELCOM (1998) Red list of marine and coastal biotopes and biotope complexes of the Baltic Sea, Belt Sea and Kattegat. Baltic Sea Environ Proc 75
Hunter RD (1976) Changes in carbon and nitrogen content during decomposition of three macrophytes in freshwater and marine environments. Hydrobiologia 51:119–128
Hutchinson GE (1975) A treatise on limnology. III. Limnological botany. Wiley Interscience, New York
Kangas P, Autio H, Hällfors G, Luther H, Niemi A, Salemaa H (1982) A general model of the decline of Fucus vesiculosus at Tvärminne, south coast of Finland in 1977–1981. Acta Bot Fenn 118:1–27
Koistinen M, Munsterhjelm R (2001) Charophytes of the Finnish coastal waters. Schriftenr Landschaftspflege Naturschutz 72:27–29
Kotta J (2000) Impact of eutrophication and biological invasions on the structure and functions of benthic macrofauna. Dissertationes Biologicae Universitatis Tartuensis, Tartu University Press 63:1–160
Kotta J, Paalme T, Martin G, Mäkinen A (2000) Major changes in macroalgae community composition affect the food and habitat preference of Idotea baltica. Int Rev Hydrobiol 85:693–701
Lotze HK, Worm B (2002) Complex interactions of climatic and ecological controls on macroalgal recruitment. Limnol Oceanogr 47:1734–1741
Lubchenco J (1978) Plant species diversity in a marine intertidal community: importance of herbivore food preferences and algal competitive abilities. Am Nat 112:23–39
Lubchenco J (1982) Effects of grazers and algal competitors on fucoid colonization in tide pools. J Phycol 18:544–550
Luther H (1979) Chara connivens in the Baltic Sea area. Ann Bot Fenn 16:141–150
Macneil C, Dick JTA, Elwood RW (1997) The trophic ecology of freshwater Gammarus spp. (Crustacea: Amphipoda): problems and perspectives concerning the functional feeding group concept. Biol Rev 72:349–364
Malm T (1999) Distribution patterns and ecology of Fucus serratus L. and Fucus vesiculosus L. in the Baltic Sea. Doctoral dissertation, University of Stockholm, Sweden
Mann KH (1988) Production and use of detritus in various freshwater, estuarine, and coastal marine ecosystems. Limnol Oceanogr 33:910–930
Martin G (2001) Distribution of charophytes in Estonian coastal waters. Schriftenr Landschaftspflege Naturschutz 72:31–36
Moore JA (1986) Charophytes of Great Britain and Ireland. Handbook No. 5. Botanical Society of the British Isles, London
Moss B (1989) Water pollution and the management of ecosystems: A case study of science and scientist. In: Grubb PJ, Whittaker JB (eds) Towards a more exact ecology. Blackwell, Oxford, pp 401–422
Moyle PB, Light T (1996) Biological invasions of fresh water: empirical rules and assembly theory. Biol Conserv 78:149–161
Naylor E (1955) The ecological distribution of British species of Idotea (Isopoda). J Anim Ecol 24:255–269
Nicotry ME (1980) Factors involved in herbivore food preference. J Exp Mar Biol Ecol 42:13–26
Nielsen R, Kristiansen A, Mathiesen L, Mathiesen H (eds) (1995) Distributional index of the benthic macroalgae of the Baltic Sea area. Acta Bot Fenn 155:1–51
Noordhuis R, van der Molen DT, van den Berg MS (2002) Response of herbivorous water-birds to the return of Chara in Lake Veluwemeer, The Netherlands. Aquat Bot 72:349–367
Olsen S (1944) Danish Charophyta. Kong Dansk Vidensk Selsk Skr 3:1–240
Orav-Kotta H, Kotta J (2003a) Food and habitat choice of the isopod Idotea baltica in the northeastern Baltic Sea. Hydrobiologia (in press)
Orav-Kotta H, Kotta J (2003b) Seasonal variations in the grazing of Gammarus oceanicus, Idotea baltica and Palaemon adspersus on benthic macroalgae. Proc Estonian Acad Sci Biol Ecol 52: 141–148
Paalme T, Kukk H, Kotta J, Orav H (2002) “In vitro” and “in situ” decomposition of nuisance macroalgae Cladophora glomerata and Pilayella littoralis. Hydrobiologia 475/476:469–476
Paine RT (1974) Intertidal community structure. Experimental studies on the relationship between a dominant competitor and its principal predator. Oecologia 15:93–120
Pankow H, Kell V, Wasmund N, Zander B (1990) Ostsee-Algenflora. Fischer, Jena
Paul VJ, Cruz-Rivera E, Thacker RW (2001) Chemical mediation of macroalgal-herbivore interactions: ecological and evolutionary perspectives. In: McClintock JB, Baker BJ (eds) Marine chemical ecology. CRC Press, Boca Raton, USA, pp 227–265
Pöckl M (1995) Laboratory studies on growth, feeding, moulting and mortality in the freshwater amphipods Gammarus fossarum and G. roeseli. Arch Hydrobiol 134:223–253
Ravanko O (1969) Benthic algae as food for some invertebrates in the inner part of the Baltic. Limnologica 7:203–205
Robertson AI, Mann KH (1980) The role of isopods and amphipods in the initial fragmentation of eelgrass detritus in Nova Scotia, Canada. Mar Biol 59:63–69
Salemaa H (1987) Herbivory and microhabitat preferences of Idotea spp. (Isopoda) in the northern Baltic Sea. Ophelia 27:1–15
Scheffer M, de Redelijkheid MR, Noppert F (1992) Distribution and dynamics of submerged vegetation in a chain of shallow eutrophic lakes. Aquat Bot 42:199–216
Schubert H, Yousef MAM (2001a) Charophytes in the Baltic Sea—threats and conservation. Schriftenr Landschaftspflege Naturschutz 72:7–8
Schubert H, Yousef MAM (2001b) Assessment of the occurrence of charophytes in shallow coastal waters of Mecklenburg-Vorpommern, Germany. Schriftenr Landschaftspflege Naturschutz 72:9–16
Siegfried CA (1982) Trophic relations of Crangon franciscorum Stimpson and Palaemon macrodactylus Rathbun: predation on the opossum shrimp, Neomysis mercedis Holmes. Hydrobiologia 89:129–139
Sinkevièienë Z, Jurgilaitë D (2001) Historical and new data on charophytes in the Lithuanian territorial waters of the Baltic Sea. Schriftenr Landschaftspflege Naturschutz 72:17–18
Sitts RM, Knight AW (1979) Predation by the estuarine shrimps Crangon franciscorum Stimpson and Palaemon macrodactylus Rathbun. Biol Bull 156:356–368
Steneck RS, Watling L (1982) Feeding capabilities and limitations of herbivorous molluscs: a functional approach. Mar Biol 68:299–312
Suursaar Ü (1995) Nutrients in the Gulf of Riga. In: Ojaveer E (ed) Ecosystem of the Gulf of Riga between 1920 and 1990. Academia 5:41–50
Sywula T (1964) A study on the taxonomy, ecology and geographical distribution of species of genus Idotea Fabricus (Isopoda, Crustacea) in Polish Baltic. 1 and 2. Bull Soc Amis Sci Lett Poznan Ser D 4:141–200
Torn K, Martin G, Kukk H, Trei T (2004) Distribution of charophyte species in Estonian coastal water (NE Baltic Sea). Sci Mar (in press)
Trei T (1991) Taimed Läänemere põhjal. Valgus, Tallinn
Vadas RL, Elner RW (1992) Plant-animal interactions in the north-west Atlantic. In: John DM, Hawkins SJ, Price JH (eds) Plant-animal interactions in the marine benthos. The Systematics Association special vol No. 46. Clarendon Press, Oxford, pp 33–36
Valiela I, McClelland J, Hauxwell J, Behr PJ, Hersh D, Foreman K (1997) Macroalgal blooms in shallow estuaries: controls and ecophysiological and ecosystem consequences. Limnol Oceanogr 42:1105–1118
Van den Berg MS (1999) Charophyte colonization in shallow lakes. Processes, ecological effects and implications for lake management. Thesis, Vrije Universiteit, Amsterdam
Wetzel RG (1983) Limnology, 2nd edn. Saunders College Publishing, Philadelphia
Williamson MH, Fitter A (1996) The characters of successful invaders. Biol Conserv 78:163–170
Wood RD, Imahori K (1965) A revision of the Characeae. First part. Monograph of the Characeae. Cramer, Weinheim
Yousef MAM, von Nordheim H, Küster A, Schubert H (1997) Eignung der Armleuchteralgen (Characeae) als Indikator für den Gewässerzustand der Flachwasserbereiche der Ostseeküste. Aktuel Prob Meeresumwelt Suppl 7:173–182
Acknowledgements
The study was financed by the Estonian Governmental Program No. 0182578s03 and Estonian Science Foundation grant No. 5103. The experiments comply with the current laws of the country in which they were performed.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by H.D. Franke
Rights and permissions
About this article
Cite this article
Kotta, J., Torn, K., Martin, G. et al. Seasonal variation in invertebrate grazing on Chara connivens and C. tomentosa in Kõiguste Bay, NE Baltic Sea. Helgol Mar Res 58, 71–76 (2004). https://doi.org/10.1007/s10152-003-0170-2
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10152-003-0170-2