
Abstract
A common characteristic observed in many biological invasions is the existence of a lag between the time of arrival by the alien population and the time when established populations are noticed. Considerable advances have been made in modeling the expansion of invading species, and there is often remarkable congruence between the behavior of these models with spread of actual populations. While these models have been used to characterize expansion of very newly founded colonies, there have been few attempts to compare the behavior predicted from theory with spread in actual newly founded populations, largely due to the difficulty of sampling sparse populations. Models predict that time lags in the radial expansion of newly invaded populations may be due to time requirements for the population to grow from founding to detectable levels. Models also indicate that these time lags can be predicted based upon population parameters such as the intrinsic rate of population growth and diffusion coefficient. In this paper, we compared the behavior of these models with historical data on gypsy moth, Lymantria dispar, establishment and spread to show similarities between model predictions and observed population spread, both of which exhibited temporal lags of expansion. However, actual populations exhibited certain behaviors that were not predicted, and this could be due, in part, to the existence of Allee effects and stochasticity. Further work that incorporates these effects is needed to more fully understand the growth of incipient colonies of invading species. Ultimately, this information can be of critical importance in the selection of effective strategies for their detection and eradication.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Biological invasions can cause profound ecological changes and consequently are considered one of the most serious environmental problems of the century (Vitousek et al. 1996; Mack et al. 2000). Understanding the population processes operating during biological invasions has emerged as a major challenge, but this subject has developed as an area where mathematical ecology has made important contributions (Simberloff 1988; Hastings 1996; Sakai et al. 2001). Three phases are generally recognized in all biological invasions: arrival, establishment, and spread (Shigesada and Kawasaki 1997; Sakai et al. 2001; National Research Council 2002). Considerable recent literature has addressed the population biology of spread and to a lesser extent the establishment phase, but here we consider a specialized topic, namely the very early period of spread of a newly established population. This topic has received little attention, in part due to the difficulty of sampling very low density, newly established populations.
A common characteristic observed in many invasions is a period of time between arrival of the alien population and its eventual growth to sufficient levels such that it is noticed (Sakai et al. 2001). Several hypotheses have been advanced to explain these time lags. One explanation is the alteration of the habitat by the invader to make it more favorable over a long time period (e.g., Rilov et al. 2004). Another hypothesis is the necessity for local adaptation by an invader, which occurs over a prolonged period (Lee 2002; Parker et al. 2003). In some systems, the delay in population expansion is thought to be caused by the accumulation of multiple introductions that provide sufficient genetic diversity for population adaptation and growth (Ellstrand and Schierenbeck 2000; Kolbe et al. 2004). In other cases, the delay may be merely the result of the slow growth of a population originating from a very small number of founding individuals (Memmott et al. 2005). Along these lines, simple population models demonstrate that lags in population growth can arise during the early stages of an invasion as a result of net population losses caused by emigration that are not compensated by reproduction or immigration (Shigesada and Kawasaki 1997; Kean and Barlow 2000).
We were motivated in part by the relatively few studies that have directly studied the growth of newly founded populations (e.g., Memmott et al. 2005). In this study, we analyzed data on newly founded gypsy moth, Lymantria dispar, populations in North America to quantify temporal lags in population growth. This polyphagous, foliage-feeding Lepidoptera species is native to most of temperate Europe and Asia and was accidentally introduced to North America in 1868 or 1869 (Liebhold et al. 1989, 1992). The species has been extensively studied and, therefore, there are ample studies documenting its life history and population biology, which facilitates the estimation of population parameters. Thus, we were able to use population parameter estimates to model lags in population growth. In addition, because the gypsy moth invasion in North America is so widely documented, we were able to utilize the wealth of existing data to reconstruct historical growth of newly formed populations.
The existence of time lags in growth of newly established gypsy moth colonies, while never formally quantified before, are generally known to occur and their existence is implicitly assumed in strategies to manage these infestations. Isolated colonies of gypsy moth, which is univoltine, are typically detected from pheromone trap captures in geographically extensive surveys; continual captures at a given location through time typically triggers more extensive trapping to delimit a population, which is then followed by management intervention to eradicate the population (Liebhold and Bascompte 2003). These steps usually require several years during which colonies do not expand rapidly. Thus, understanding the dynamics of colony expansion during the early phase of colony growth could provide useful information for increasing the efficiency of management strategies.
Modeling the growth of isolated gypsy moth populations
Probably the simplest mathematical representation of range expansion by an alien population is that of Skellam (1951) who modeled spread as a combination of exponential growth and random (diffusive) movement. Skellam used this model to show that the rate of radial range expansion asymptotically reaches a constant value, c, that can be estimated as
where r is the intrinsic rate of increase and D is the diffusion coefficient. This equation holds true in other situations (e.g., logistic growth) though spread rates may be quite different under other conditions, such as in the presence of Allee effects (Lewis and Kareiva 1993) or more complex forms of dispersal (Shigesada et al. 1995; Clark et al. 2001). Nevertheless, Eq. 1 has performed remarkably well at predicting rates of spread of many invading species (Andow et al. 1990; Shigesada and Kawasaki 1997).
Shigesada and Kawasaki (1997) used Skellam’s (1951) model to derive an equation for the radius, x t , of an invading colony defined by threshold (of detection) density, n*
where n 0 is the initial number of individuals. This equation can be used to model radial range expansion for a variety of threshold densities (e.g., Fig. 1). One interesting aspect of this equation is that for many values of r, D and n*, there is a lag between the time of initial colony seeding and the time at which colony radius, defined in terms of n*, starts increasing. Shigesada and Kawasaki note that this delay is affected by r, D, n 0 and n* and that all four parameters can be combined as
They derive an expression that relates t e , the time to expansion (the lag between arrival and growth of the colony measured by n*), to γ:
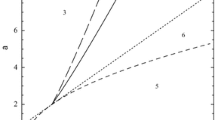
Using this equation, the relationship between γ and rt e can be graphed (Fig. 2). For large values of γ (i.e., r and n 0 are large and/or D and n* are small) the time to expansion is near zero; that is range expansion begins very quickly following arrival. However when γ exceeds 4π/e ≅ 6.223, t e abruptly becomes zero.

Radial spread of a hypothetical isolated population predicted using Eq. 2 and parameters realistic for North American gypsy moth, Lymantria dispar, populations (n 0=0.1, r=2, D=0.003)

Relationship between γ (defined in Eq. 3) and the product of time to expansion, t e , by r
These relationships thus provide a demographic explanation to the time lags so frequently observed in the early phases of invasion by alien species. We sought to determine whether these demographic processes could explain the delays observed in the spread of gypsy moth populations in North America. Therefore, we used ‘realistic’ parameters in Eq. 2 to predict radial expansion for a series of abundance thresholds n*=1, 10, 100, 1,000 and 10,000 (Fig. 1). In these calculations, we used r=2 which assumes an average ∼7.3 fold increase in gypsy moth density per generation. While gypsy moth fecundity can be high (e.g., 700 eggs per female), populations are subjected to a variety of sources of mortality and generational replacement rates rarely exceeds tenfold. We assumed that n 0=0.1, and a diffusion coefficient of D=0.003 km2/generation. Females from North American gypsy moth populations are incapable of flight and the primary active dispersal mechanism is windborne movement of first instars hanging on silken threads. Our value of D was calculated by Shigesada and Kawasaki (1997) from an experimental release-recapture experiment (cf. Mason and McManus 1981).
Simulations based upon Eq. 2 indicated that at very low threshold densities (e.g., n*=1, 10), spread can be expected to commence immediately following arrival. In contrast, for higher density levels, spread is delayed. For values of n*=1, 10, 100, 1,000, 10,000 corresponding values of γ are 666.7, 66.7, 6.7, 0.67 and 0.067 and the corresponding values of t e are 0, 0, 0, 2.2 and 3.6 years.
Historical growth of isolated gypsy moth populations
Since the time of its initial release in Medford, Mass., USA, the gypsy moth has been gradually expanding its range. With the exception of a disjunct established population that appeared in Michigan in the 1980s, spread has proceeded through expansion from the primary focus in Medford. In many years, populations are discovered in outlying areas (e.g., California), but they are generally quickly eradicated.
Medford, Massachusetts population, 1868–1890
The accidental release of the gypsy moth in North America is known to have been carried out by E. Leopold Trouvelot, an amateur naturalist, in either 1868 or 1869 (Liebhold et al. 1989). Precise details are not known but it is believed that the release occurred from populations that he was maintaining under culture on trees in the backyard of his home on 27 Myrtle Street, Medford, Mass., USA. A volume by Forbush and Fernald (1896) provides extensive details on the subsequent spread of gypsy moth through his neighborhood. This description includes accounts by various residents of the first date that they noticed large numbers of gypsy moth caterpillars and the locations of their residences are noted on a map. Based upon these narrations and map, we were able to reconstruct the historical spread of outbreak levels of the gypsy moth from 1870 to 1890. Extensive, but unsuccessful, efforts to eradicate the population did not commence until 1890, thus we may consider the spread during this interval as generally unimpeded and ‘natural’.
Figure 3 depicts the historical spread of gypsy moth from 1870 to 1890 as documented by Forbush and Fernald (1896). The first observation of outbreak levels was made in 1880 by William Taylor who lived at 27 Myrtle Street, formerly the house occupied by Trouvelot. The next observation of populations was by William Belcher, who lived at 29 Myrtle Street, the house immediately next door to the former Trouvelot residence. The graph is indicative of an acceleration of radial range expansion, possibly reaching a constant rate of spread of ca. 2 km/year which was approximately the rate of radial range expansion observed from 1900 to 1912 by Liebhold et al. (1992) from more extensive data.

Graph of distance from the point of introduction (Trouvelot’s house) as a function of year of first record of gypsy moth infestation recorded in Forbush and Fernald (1896)
We do not know exactly how many gypsy moth individuals Trouvelot introduced though an approximation of 0.1–1 egg masses seems reasonable (presumably they escaped as larvae). We also do not know what the threshold was for detection of gypsy moths by residents in Medford, but we can approximate that this was somewhere between 100 and 1,000 egg masses/ha, which is roughly the range of densities when numerous caterpillars become evident (Gottschalk 1993). Thus, the detection density (n*) might be about 100–1,000 times the inoculum density (n 0). We can see from Fig. 3 that there was an 11- to 12-year delay between the time of arrival and the date of first spread at the detection threshold (1890, when Mr. Taylor noticed large numbers of caterpillars). However, a detection threshold ca. 1,000 times larger than the inoculum should only exhibit a delay before expansion of 3–4 years according to theory (Fig. 1). Also, the Medford population exhibited a distinctly accelerating pattern of radial spread (Fig. 3), but theory (Fig. 1) does not predict such behavior.
Compton, Arkansas population, 1982–1994
Over the last 40 years, the gypsy moth has been gradually expanding its range in North America. Well ahead of this gradual expansion, isolated colonies sometimes form when humans accidentally transport gypsy moth life stages (e.g., egg masses on automobiles or recreational vehicles) from the infested area to an uninfested region (Liebhold et al. 1992; Sharov and Liebhold 1998; Whitmire and Tobin 2006). Most of the uninfested area is currently surveyed annually using pheromone traps to detect such incipient populations. Repeated captures of moths in traps at the same location or the discovery of egg masses or other life stages usually indicates the existence of a reproducing population. When such populations are discovered, they are usually delimited (using grids of pheromone traps) and then eradicated using aerial applications of microbial or chemical pesticides (Liebhold and Bascompte 2003).
Prior to ca. 1995, some portions of the uninfested portion of the USA were not surveyed for gypsy moth presence on a regular basis using pheromone traps. Consequently, there have been a handful of populations that were not detected until they grew to such a size and intensity that citizens “stumbled upon” an outbreak. One such population had its core just west of Compton, Arkansas, but the population was ultimately discovered to extend over a 17,604-ha area in northern Arkansas and southern Missouri (Table 1). The population was believed to have been founded in 1982 or 1983 when a recreational vehicle was transported from an area with high-density gypsy moth populations in New Jersey. No precise information is available, but the vehicle may have had 10–100 egg masses located on various surfaces and larvae from these masses apparently founded the population. This colony was not discovered until 1992 when ≈1 ha was noticed to be completely defoliated and some trees were found to have thousands of egg masses. The colony was not fully delimited until 1993 and 1994; and this more intensive trapping indicated that populations extended over an area of 17,604 ha. Pesticides were applied in 1993, 1994 and 1995, and, by 1996, trapping data indicated that eradication had been successful.
While data on this infestation are relatively sparse, there is a remarkable similarity to the Medford population in that it took 9–10 years for populations in the core area to increase to high densities such that it was noticed (Table 1).
Isolated populations in North Carolina and Virginia, 1985–2005
As part of an integrated effort to slow gypsy moth spread, traps are deployed annually in a 2-km grid along a 100-km band following the entire gypsy moth population front. Similar to the more distant uninfested area described above, these traps are used to detect isolated colonies so that they can subsequently be delimited and treated (Sharov et al. 2002; Tobin et al. 2004). This program adopts a strategy in which isolated colonies along the expanding front are retarded in order to reduce spread (Sharov and Liebhold 1998).
We used historical data collected as part of this barrier zone project to quantify the growth of four isolated gypsy moth populations located in Virginia and North Carolina (Table 2). These populations were selected for this analysis because they arose well ahead of the continuously infested area and can therefore be assumed to be growing independently of propagule pressure from the principle infested area.
Annual pheromone trap catch data were used to compute annual rates of colony expansion for a range of threshold trap count values. For each population, we interpolated trap catch data using median indicator kriging (Isaaks and Srivastava 1989) to generate a 500×500 m2 raster matrix of interpolated trap counts (e.g., Fig. 4a). We then imported these interpolated raster matrices into a GIS and calculated the area of each colony above a series of threshold trap count values in each year and then used these areas to compute annual average colony radii for each threshold (e.g., Fig. 4b).

Historical data on the growth of an isolated gypsy moth population near Hiawasee (Caroll, Pulaski and Floyd counties), Virginia, 1988–1991. a Interpolated pheromone trap count surfaces. b Time series of mean colony radii defined by five trap capture thresholds derived from interpolated surfaces
Hiwasee, Virginia population, 1988–1991
The Hiwasee, Virginia, population was located on the border of Carroll, Pulaski and Floyd counties in Virginia, and was first detected in 1989. It was delimited with a higher trap density in 1990 and 1991, after which it was eradicated. Graphs of radial expansion based on empirical data (Fig. 4b) bear some functional resemblance to theoretical expectations (Fig. 1) in that, for larger thresholds, there was a delay in the timing of initial expansion. It is also interesting to note that, for the three smallest thresholds (0, 1, 3), radial expansion proceeded quickly, then slowed and actually reversed (i.e., populations recede). While this particular behavior is not seen in Fig. 1, it is predicted by theory to occur in the very early stages of colony growth (Shigesada and Kawasaki 1997; Kean and Barlow 2000) when net emigration exceeds net growth with the consequence of a reduction in the core population size.
Henderson County, North Carolina population, 2003–2005
The isolated population in Henderson County, North Carolina, was first apparent in 2003 (Fig. 5). At the time of this writing, eradication treatments had never been applied. Radial spread of the Henderson population behaved slightly different from that seen in the Hiwasee population (Fig. 4b) and substantially different from that predicted by the model (Fig. 1). Expansion occurred at all thresholds in 2003, but then receded at all thresholds in 2004 and then all thresholds expanded again in 2005 (Fig. 5).

Historical data on the growth of an isolated gypsy moth population near Henderson County, North Carolina, 2002–2005. a Interpolated pheromone trap count surfaces. b Time series of mean colony radii defined by five trap capture thresholds derived from interpolated surfaces
Eastern Warren County, North Carolina population, 1985–1988
The isolated population in eastern Warren County, North Carolina, was first apparent in 1985 and gradually expanded (Fig. 6). Its colony radius steadily increased except for the zero threshold radius which stayed constant (this may have reflected very low levels of randomly dispersing males) and the >1 threshold which temporarily receded in 1986 (Fig. 6b).

Historical data on the growth of an isolated gypsy moth population in eastern Warren County, Virginia, 1984–1988. a Interpolated pheromone trap count surfaces. b Time series of mean colony radii defined by five trap capture thresholds derived from interpolated surfaces
Western Warren County, North Carolina population, 1986–1988
An isolated population in western Warren County, North Carolina, was first apparent in 1986 and gradually expanded (Fig. 7). Its colony radius steadily increased except for the zero threshold radius which stayed constant (this may have reflected very low levels of randomly dispersing males) (Fig. 7b).

Historical data on the growth of an isolated gypsy moth population in western Warren County, Virginia, 1985–1988. a Interpolated pheromone trap count surfaces. b Time series of mean colony radii defined by five trap capture thresholds derived from interpolated surfaces
Discussion
Time lags between the arrival of an alien population and the commencement of range expansion has been observed for many species (Sakai et al. 2001). Several theories have been advanced to explain these time lags (Ellstrand and Schierenbeck 2000; Lee 2002; Parker et al. 2003; Kolbe et al. 2004; Rilov et al. 2004). However, application of Skellam’s (1951) simple reaction-diffusion model predicts that such delays in expansion may occur for purely demographic reasons (Shigesada and Kawasaki 1997; Kean and Barlow 2000). Despite the simplicity of this model it does capture many critical characteristics of the growth of newly founded populations. Unfortunately, there have been few previous attempts to compare the behavior of this model to the growth of actual invading populations.
The gypsy moth invasion in North America, coupled with a spatially- and temporally-extensive sampling program to detect the presence of isolated colonies, provides a unique opportunity to describe the growth of newly found populations during the early phase of invasion. In our analysis of historical records of the growth of isolated gypsy moth populations, we observed that time lags between initial population founding and range expansion were common. Moreover, application of Skellem’s (1951) model using parameter values realistic for gypsy moth, predicted time lags of 2–5 years for some larger population thresholds (Fig. 1). However, it took 11–12 years before gypsy moth populations were observed spreading into the vicinity directly surrounding the site of initial introduction in Medford, Massachusetts (Fig. 3). Similarly, the Arkansas population did not reach noticeable high densities until 9–10 years following introduction (Table 1). More extensive characterizations of populations were available from pheromone traps placed in grids (Figs. 4, 5, 6, 7) but, for those populations, information was lacking on the date of the initial founding event. It is quite likely that these colonies were founded many years before they grew to sufficient size such that they were detected in trap grids.
Data from these trapping grids (Figs. 4, 5, 6, 7) indicate that it is not uncommon for populations to exhibit rapid initial spread, followed by a deceleration of expansion or, in some cases, populations receded. Several authors have pointed out that during the period directly following arrival of founding population, net emigration may exceed net immigration and this can lead to a latency in the apparent growth of an invading population (Shigesada and Kawasaki 1997; Kean and Barlow 2000). However, it seems unlikely that the colony recessions seen in the trap data were examples of this type of phenomenon. The period of population latency, as predicted by theoretical immigration/emigration deficits, should directly follow the first arrival of founding individuals. While we do not know precisely when the founding individuals arrived, we contend that they arrived well before they were first detected in pheromone traps. Thus, the observed declines in male moth abundance as measured by traps are probably not examples of the theoretical demographic latency described above. Rather, we suspect that many of the declines seen from the time series of colony expansion (Figs. 4, 5, 6, 7) are more likely due to inter-annual stochasticity in population growth rates. Stochasticity, while not included in Skallam’s (1951) model (Fig. 1), appears to play an important role in the growth of newly founded populations. In addition to stochasticity in growth rates, dispersal may be stochastic. Yamamura (2004) showed that stochasticity in dispersal distances can affect spread rates and can also delay the initial expansion of newly invaded populations, much as we observed in the Massachusetts and Arkansas populations.
In addition to stochasticity, another key population process missing from the model described by Eq. 2 and illustrated in Fig. 1 is Allee dynamics. The Allee effect describes any relationship in which net reproductive growth declines with decreasing density (Courchamp et al. 1999; Stephens et al. 1999). Low-density gypsy moth populations are known to be affected by Allee effects, most likely due to difficulty in mate finding (Sharov et al. 1995), which can lead to extinction of very low density, isolated populations (Liebhold and Bascompte 2003; Whitmire and Tobin 2006). Furthermore, Allee effects can cause a reduction in the rate of range expansion (Lewis and Kareiva 1993; Hastings et al. 2005). It is quite likely that Allee effects may have contributed to the longer than expected time lags in spread that we observed in the Medford and Arkansas populations (Fig. 3, Table 1).
Gypsy moth range expansion is thought to follow a type of stratified dispersal in which short-range movement is due to the windborne dispersal of first instars while long range movement occurs through accidental movement of life stages (Liebhold et al. 1992; Sharov and Liebhold 1998). However, it seems unlikely that this process of stratified dispersal would be important in explaining observed patterns of expansion of very small isolated gypsy moth colonies because long-range movement of gypsy moth life stages would most likely be expected only when populations reach high enough densities such that there is an appreciable probability of accidental movement. Thus, when population levels are low, short-range movement of first instars would be expected to be the dominant mode of population dispersal.
Understanding the dynamics and spread of newly founded alien populations is critical to development of effective strategies to minimize invasions. The detection and eradication of isolated gypsy moth populations, including those well beyond the expanding population front, consumes considerable resources in North America as well as elsewhere where this species is not native (e.g., New Zealand). The finding of ≈10-year time lags between the time of colony initiation and the development of high density gypsy moth populations suggests that isolated colonies need not be treated immediately upon detection. Instead, there is sufficient time to deploy grids of traps in successive years to delimit the geographical extent of the population and hence maximize the efficiency of a site-specific eradication effort. Presumably, once populations reach high densities, Allee effects become less of a factor while the accidental movement of life stages becomes a more common occurrence, which in turn would lead to an increase in colony growth rate and thus possibly render eradication efforts more difficult.
References
Andow DA, Kareiva PM, Levin SA, Okubo A (1990) Spread of invading organisms. Landsc Ecol 4:177–188
Clark JS, Lewis MA, Horvath L (2001) Invasion by extremes: population spread with variation in dispersal and reproduction. Am Nat 157:537–554
Courchamp F, Clutton-Brock T, Grenfell B (1999) Inverse density dependence and the Allee effect. Trends Ecol Evol 14:405–410
Ellstrand NC, Schierenbeck KA (2000) Hybridization as a stimulus for the evolution of invasiveness in plants? Proc Nat Acad Sci USA 97:7043–7050
Forbush EH, Fernald CH (1896) The gypsy moth, Porthetria dispar (Linn). Wright and Potter, Boston
Gottschalk KW (1993) Silvicultural guidelines for forest stands threatened by the gypsy moth. US. Department of Agriculture Forest Service, Northeastern Forest Experiment Station, general technical report NE-171:49
Hastings A (1996) Models of spatial spread: a synthesis. Biol Conserv 78:143–148
Hastings A, Cuddington K, Davies KF, Dugaw CJ, Elmendorf S, Freestone A, Harrison S, Holland M, Lambrinos J, Malvadkar U, Melbourne BA, Moore K, Taylor C, Thomson D (2005) The spatial spread of invasions: new developments in theory and evidence. Ecol Lett 8:91–101
Isaaks EH, Srivastava RM (1989) An introduction to applied geostatistics. Oxford University Press, Oxford
Kean JM, Barlow ND (2000) Effects of dispersal on local population increase. Ecol Lett 3:479–482
Kolbe JJ, Glor RE, Schettino LR, Lara AC, Larson A, Losos JB (2004) Genetic variation increases during biological invasion by a Cuban lizard. Nature 431:177–181
Lee CE (2002) Evolutionary genetics of invasive species. Trends Ecol Evol 17:386–391
Lewis MA, Kareiva P (1993) Allee dynamics and thje spread of invading organisms. Theor Popul Biol 43:141–158
Liebhold AM, Bascompte J (2003) The Allee effect, stochastic dynamics and the eradication of alien species. Ecol Lett 6:133–140
Liebhold A, Mastro V, Schaefer PW (1989) Learning from the legacy of Leopold Trouvelot. Bull Entomol Soc Am 35:20–22
Liebhold AM, Halverson JA, Elmes GA (1992) Gypsy moth invasion in North America: a quantitative analysis. J Biogeogr 19:513–520
Mack RN, Simberloff DS, Lonsdale WM, Evans HF, Clout M, Bazzaz FA (2000) Biotic invasions: causes, epidemiology, global consequences and control. Ecol Appl 10:689–710
Mason CJ, McManus ML (1981) Larval dispersal o f the gypsy moth. In: Doane CC, McManus ML (eds) The gypsy moth: research toward integrated pest management. Technical bulletin, vol 1584. US Department of Agriculture, Washington
Memmott J, Craze PG, Harman HM, Syrett P, Fowler SV (2005) The effect of propagule size on the invasion of an alien insect. J Anim Ecol 74:50–62
National Research Council (2002) Predicting invasions of nonindigenous plants and plant pests. National Academy Press, Washington, pp 194
Parker IM, Rodriguez J, Loik ME (2003) An evolutionary approach to understanding the biology of invasions: local adaptation and general-purpose genotypes in the weed Verbascum thapsus. Conserv Biol 17:1–59
Rilov G, Benayahu Y, Gasith A (2004) Prolonged lag in population outbreak of an invasive mussel: a shifting-habitat model. Biol Invasions 6:347–364
Sakai AK, Allendorf FW, Holt JS, Lodge DM, Molofsky J, With KA, Baughman S, Cabin RJ, Cohen JE, Ellstrand NC, McCauley DE, O’Neil P, Parker IM, Thompson JN, Weller SG (2001) The population biology of invasive species. Annu Rev Ecol Syst 32:305–332
Sharov AA, Liebhold AM (1998) Model of slowing the spread of gypsy moth (Lepidoptera: Lymantriidae) with a barrier zone. Ecol Appl 8:1170–1179
Sharov AA, Liebhold AM, Ravlin FW (1995) Prediction of gypsy moth (Lepidoptera: Lymantriidae) mating success from pheromone trap counts. Environ Entomol 24:1239–1244
Sharov A, Leonard D, Liebhold A, Roberts A, Dickerson W (2002) “Slow the spread”: a national program to contain the gypsy moth. J Forest 100:30–36
Shigesada N, Kawasaki K (1997) Biological invasions: theory and practice. Oxford University Press, New York
Shigesada N, Kawasaki K, Takeda Y (1995) Modeling stratified diffusion in biological invasions. Am Nat 146:229–251
Simberloff D (1988) The contribution of population and community biology to conservation science. Annu Rev Ecol Syst 19:473–511
Skellam JG (1951) Random dispersal in theoretical populations. Biometrika 38:196–218
Stephens PA, Sutherland WJ, Freckleton RP (1999) What is the Allee effect? Oikos 87:185–190
Tobin PC, Sharov AA, Liebhold AM, Leonard DS, Roberts EA, Learn MR (2004) Management of the gypsy moth through a decision algorithm under the STS Project. Am Entomol 50:200–209
Vitousek PM, D’Antonio CM, Loope LL, Westbrooks R (1996) Biological invasions as global environmental change. Am Sci 84:468–478
Whitmire SL, Tobin PC (2006) Perisistance of invading gypsy moth populations in the United States. Oecologia 147:230–237
Yamamura K (2004) Dispersal distance of corn pollen under fluctuating diffusion coefficient. Popul Ecol 46:87–101
Acknowledgments
We thank Donna Leonard, David Blackburn, John Ghent and Bobbe Fitzgibbon for providing historical information on isolated gypsy moth colonies. We also thank Laura Blackburn for technical assistance.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Liebhold, A.M., Tobin, P.C. Growth of newly established alien populations: comparison of North American gypsy moth colonies with invasion theory. Popul Ecol 48, 253–262 (2006). https://doi.org/10.1007/s10144-006-0014-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10144-006-0014-4