
Abstract
An experimental microdiet prepared using an internal gelation method was used to partially replace the traditional live feed (Artemia) for larval Atlantic halibut, Hippoglossus hippoglossus L. Three trials were conducted with microdiet introduced at 20, 32, and 43 days post first feeding and larvae were sampled at approximately 2, 13, 23, and 33 days after microdiet introduction in each trial. The success of feeding was assessed by morphometrics and histological analysis of gut contents. Microdiet particles were readily consumed after a period of adaptation and provided an adequate source of nutrients with no significant increase in mortality in the microdiet-fed group compared to the control group. However, growth was limited and there was an increased incidence of malpigmentation of the eye and skin. Subtle changes in underlying digestive and developmental physiology were revealed by microarray analysis of RNA from control and experimental fish given microdiet from day 20 post first feeding. Fifty-eight genes were differentially expressed over the four sampling times in the course of the trial and the 28 genes with annotated functions fell into five major categories: metabolism and biosynthesis, cell division and proliferation, protein trafficking, cell structure, and stress. Interestingly, several of these genes were involved in pigmentation and eye development, in agreement with the phenotypic abnormalities seen in the larvae.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Atlantic halibut (Hippoglossus hippoglossus L.) shows good potential for cold-water aquaculture (Berg 1997; Mangor-Jenson et al. 1998) and is highly valued for food (Blanchard and Scarratt 2002). High mortality during the early stages of feeding remains as one of the major bottlenecks for the development of the halibut aquaculture industry. Low survival of larvae is often linked to nutrient deficiencies and their poor performance on start feeding diets. At the early stages of development, halibut larvae are fed live food organisms such as copepods, rotifers, and Artemia (Hamre et al. 2002; Evjemo et al. 2003; Lie and Lambertsen 1985; Solbakken et al. 2002; Berg 1997; Mangor-Jenson et al. 1998; Ronnestad et al. 1998). These organisms show wide variations in food quality and nutrient composition. Generally, Artemia and rotifers are enriched with emulsions based on formulations that contain highly unsaturated fatty acids and specific micronutrient supplements to improve the nutritional quality of live feed organisms. In the past three decades, significant research efforts have been directed towards replacing live food organisms with dry microdiets; however, they have achieved limited success in improving the survival and growth of several cold-water marine fish larvae. Microencapsulated diets have recently been studied as alternatives to live organism feeds for sea bream (Yúfera et al. 2000, 2003, 2005), Dover sole (Applebaum 1985), and Senegalese sole (Yúfera et al. 2003, 2005). However, little information is available for early weaning of larval Atlantic halibut (Hamre et al. 2001; Naess et al. 2001).
Microencapsulation, a technique whereby stable particles with minimal nutrient leaching are produced, has been used for the production of larval diets. These diets improve the delivery of essential nutrients to the larvae, minimize nutrient leaching, and prevent subsequent water quality problems (Jones et al. 1974; Langdon 2003). Digestibility of the capsules, ability to retain nutrients, nutrient load in the encapsulated particle, durability, and minimal use of toxic substances are key qualities in the design of microencapsulated diets (Yúfera et al. 2005). Previous studies with microencapsulated diets showed low growth of fish larvae due to inadequate diet formulation and inappropriate particle structure. In many cases, good larval growth is only achieved with microbound diets when cofeeding with live prey takes place (Langdon 2003). It is possible that adding feeding stimulants, enzymes, or nutrients in live feed could improve the utilization of artificial diets (Kolkovski et al. 1997). For Atlantic halibut, a microbound diet was only effective if introduced 20 days post hatching (Hamre et al. 2001). However, a microencapsulated diet was able to substitute for live food during the early stages of larval rearing in gilthead sea bream larvae, although again only limited growth was achieved (Yúfera et al. 1999). The need for properly formulated feed with correct nutrient balance is necessary to increase feed utilization and improve larval growth.
Different forms of microencapsulated diet for larval fish have been developed in past years (Langdon 2003; Yúfera et al. 1999; Önal and Langdon 2005). Each one has advantages that must be considered in finding equilibrium between stability to prevent excessive micronutrient leaching before being ingested and sufficient digestibility for larval fish with immature digestive tracts. Recently, Yúfera et al. (2005) developed a method for producing microparticles by internal gelation whereby sodium alginate reacted with a calcium solution to produce calcium alginate. This method allows the use of practical ingredients and environmentally friendly reagents during the preparation. The microdiets elaborated using this method are able to support growth in larval fish (Yúfera et al. 2005).
Eye development, eye migration, skeletal development, successful metamorphosis, and pigmentation are crucial aspects in the growth of halibut larvae and are all affected by diet (Vilhelmsson et al. 2004; Hamre et al. 2003; Naess and Lie 1998; Gara et al. 1998; Lewis et al. 2004; Lewis-McCrea and Lall 2007; Hamre et al. 2005; Solbakken et al. 2002; Hamre et al. 2002; Bell et al. 2003). Few studies have employed microarray analysis to investigate the growth of fish in response to dietary changes and none have yet been performed on flatfish. A small-scale nutrigenomic study investigated changes in gene expression in Atlantic salmon liver resulting from partial replacement of fish oil with rapeseed oil (O.-Jordal et al. 2005) and a more comprehensive microarray analysis of the effect of ration on growth hormone transgenic Atlantic salmon has been performed (Rise et al. 2006). Three studies have investigated changes in gene expression during larval development of sea bass (Darias et al. 2008), sea bream (Sarrapoulou et al. 2005), and Atlantic halibut (Douglas et al. 2008).
The present study seeks to determine whether a microencapsulated diet produced by internal gelation (referred to as the microdiet) can be successfully used to partially replace the traditional live feed (Artemia) for larval Atlantic halibut, H. hippoglossus L. The success of feeding was assessed by morphometrics and histological analysis of gut contents. A custom Atlantic halibut microarray representing over 9,000 genes (Douglas et al. 2008) was used to assess whether the expression of genes involved in digestion and absorption of macronutrients was altered in the larvae as a result of the introduction of the microdiet.
Methods
Experimental Feed
The microencapsulated particles were prepared by a modified method of internal gelation, as described by Yúfera et al. (2005). The mixed dietary ingredients (10% w/v; Table 1) and calcium citrate tetrahydrate 1% w/v (Aldrich, Oakville, ON, Canada) were blended into a 1.5% w/v sodium alginate solution of medium viscosity (MP Biochemicals, Santa Ana, CA, USA). Two parts of the homogenized solution were emulsified in five parts of a soy lecithin (LV Lomas, Brampton, ON, Canada) and sunflower oil solution (2% w/v) for 10 min at 800 rpm in a Stir-pak laboratory mixer (Cole-Palmer Instrument; Model 4554-10) at ambient temperature. A glacial acetic acid (Caledon, Georgetown, ON, Canada) and sunflower oil solution (1/1) was added to the emulsion (4% v/v of total oil volume) and dispersed for 10 min. The microparticles settled and supernatant oil was decanted and further separated by centrifugation at 1,000 rpm for 2 min under ambient temperature (Centra CL3R; Model 120; Thermo EIC). The microparticles were stirred into an anhydrous calcium chloride solution (0.5% w/v; 0.035 mM; Panreac, Barcelona, Spain) for 8 min and settled before decanting the upper portion and then separated in a 200-µm sieve. The recovered particles were dispersed into a Tween 80 (Anachemia, Mississauga, ON, Canada) solution (1%) for 5 min, sieved, and then washed with fresh water to remove debris. The capsules were freeze-dried to produce the final microdiet.
Fish Feeding and Rearing
All fish rearing was conducted as part of a production run at Scotian Halibut Ltd., Clark's Harbour, Nova Scotia, Canada. Experimental fish were housed in parallel cylindrical tanks; each measured approximately 40 cm in width and 100 cm in depth. All halibut larvae used for the experiment were transferred from a single 7-m3 tank with flow-through salt water (32 ppt) maintained at 11 ± 0.2°C using a heat exchanger. The larvae were transferred at three ages: 20 days (trial 1), 32 days (trial 2), and 43 days (trial 3; 492, 628, and 706 degree days, respectively) post-Artemia (Artemia salina; Gulf Breeze Aquaculture & Seafood, Fresno, CA, USA) introduction. Approximately 200 fish were transferred to each of three replicate treatment tanks for each trial. Due to space limitations, the 7-m3 production tank was considered the control treatment and fish were sampled from this tank at the same times as the experimental fish.
All tanks were fed Artemia twice daily at approximately 7 a.m. with 3 × 105 cells and at 4 p.m. with 6 × 105 cells. The experimental larvae, in addition to the Artemia feed, were fed the microdiet every half hour to excess. Artemia feed amounts were adjusted in the control tank over time according to how quickly the Artemia were cleared from the tank after feeding; however, the experimental larvae were fed a constant amount twice daily of Artemia and microdiet to excess until day 40 when Artemia was discontinued and larvae were fed only microdiet to excess. The microdiet was then fed for an additional 10 days. Trial 3 larvae that were transferred on day 43 were cofed Artemia and microdiet on the day of transfer only and then fed exclusively microdiet until day 53. Larvae in all tanks were fed a commercial dry diet (Gemma micro, Skretting, Bayside, NB, Canada) to excess from day 53 onwards.
Oxygen and temperature were measured daily in the experimental tanks from when larvae were first transferred until the end of the trial. Mortalities for experimental tanks were removed daily and counted. Mortalities for the control tank were measured daily from day 20 until the end of the trial. Feeding times for both the microdiet and the Artemia were also recorded daily.
Sampling and Measurement of Larvae
The control group and each replicate from the three experimental groups were sampled throughout the trial to obtain material for microarray and histological analyses. Sampling occurred at days 2 and 10–13 after transfer for all three trials. Additional sampling occurred at days 21–23 after transfer for trials 1 and 2 and also at day 33 after transfer for trial 1.
For larvae up to 30 days post first feeding, 50 individuals were sampled for RNA work and for histology. For older larvae, 20 individuals were sampled. Larvae were euthanized with an overdose of tricaine methanesulfonate (MS-222; Argent Chemical Laboratories, Redmond, WA, USA) and placed in 50 mL tubes with twice their volume of RNALater (Ambion, Austin, TX, USA). Samples for histology were fixed overnight at 4°C by immersion in 4% v/v paraformaldehyde in Tris–HCl (pH 7.8) and processed for paraffin embedding (Murray et al. 2006). Ten fixed larvae at each sampling point were measured for weight, length, and width. All fish were photographed and measured using the Nikon AZ100 microscope (Nikon Canada, Mississauga, ON, Canada) with the NIS Elements Advanced Research package V.3.0. All fish were photographed against a white background and were held flat with a microscope slide. Lengths were measured from the tip of the snout/mouth to the terminus of the body muscle not including the caudal fin rays (standard length). The widths were measured by drawing a line perpendicular to the length measure, starting the line at a point near the anus. The width measures did not include ventral or dorsal fin areas.
Microscopy
Paraffin-embedded tissues were sectioned at 7 µm, mounted on uncoated glass slides, dried briefly, and then baked overnight at 60°C to enhance adherence to the slides. For general histological examination, sections were deparaffinized, rehydrated, and then stained with hematoxylin and eosin using standard procedures. Four slides were prepared from each fish and four serial sections were examined from each for liver morphology and gut contents. Feed intake was calculated as the percent of the gut filled by food for each of four to six individuals per sampling time.
Image Capture and Analysis
The microscopy images for control and experimental individuals were captured and analyzed for the “percentage of gut filled by food” by performing area measurements using the Simple PCI 6.0 software (Simple PCI, Hamamatsu 2008).
Statistics
A one-way analysis of variance (ANOVA) was performed using SAS Version 9.1 (SAS Institute, Cary, NC, USA) to analyze the feed consumption data and calculate the significant difference between the experimental and control fish for “percentage of gut filled by food.” Statistical analysis of comparative morphometric data was performed using one-way ANOVA in Excel (Microsoft, Redmond, WA, USA).
RNA Extraction
For larvae transferred at day 20 (trial 1), RNA was isolated from five whole individuals by homogenizing in 1.5 ml Trizol (Invitrogen, Burlington, ON, Canada) according to the manufacturer's recommendation. For control samples, fish were randomly taken from the three replicate samples at each of the sampling times and processed as above, giving a pooled control for each sampling time. After rough quantitation, 100 µg of each RNA sample was cleaned using the RNeasy kit (Qiagen, Mississauga, ON, Canada) following the RNA Cleanup protocol. A 2-µl aliquot of each eluted sample was quantified using a NanoDrop Spectrophotometer (Nanodrop Technologies, Wilmington, DE, USA) and RNA integrity was determined using a Bioanalyzer 2100 (Agilent Technologies, Mississauga, ON, Canada). The RNA integrity number generated for all RNA samples was over 8.5.
Microarray Experiments
Since early weaning was the focus of this study, microarray experiments were performed for larvae from trial 1 only. cDNA was prepared from 1 µg of total RNA from each of the three replicates at each time point (days 2, 13, 23, and 33 after sampling) using the Array 900 kit (Genisphere, Hatfield, PA, USA) and labeled with Alexa Fluor™ 647 dye (see Fig. 1). This was hybridized to the Atlantic halibut oligonucleotide array, which contains 9,277 unique features from Atlantic halibut printed in quadruplicate (Douglas et al. 2008), with control cDNA prepared from 1 µg of a pooled RNA from fish randomly sampled from the production tank at the same time points and labeled as above with the Alexa Fluor™ 546 dye. The formamide-based hybridization buffer was used and hybridization was performed overnight at 43°C. Capture reagent (2.5 µl) was added in 30 µl of hybridization buffer and the microarray incubated at 52°C for 4 h.

Experimental design for gene expression analysis of fish from trial group 1 fed microdiet at 20 days post first feeding. RNA from experimental fish sampled at D2, D13, D23, and D33 is shown in boxes on the left and RNA from control fish sampled at the same times is shown in boxes on the right. Replicates for experimental samples are designated E1, E2, and E3. Pooled control RNA samples are designated C
Microarrays were scanned at 543 and 633 nm using a ScanArray® 5000XL Microarray Acquisition scanner (Packard Bioscience, Billerica, MA, USA) at a resolution of 10 µm. Laser power was set at levels between 80% and 100% and photomultiplier tube settings were set at values ranging from 70% to 95% to adjust Alexa 555 and Alexa 647 channels on individual slides. Spot intensities were measured using the SpotReader version 1.3 (Niles Scientific, Portola Valley, CA, USA) software, and the CSV files loaded into the ArrayPipe (Hokamp et al. 2004) server at the Institute for Marine Biosciences.
Markers (blanks and those containing Arabidopsis controls) were flagged and the remaining spots were corrected for background using the “limma normexp BG correction” option. Background-corrected spots were normalized using the “limma loess (subgrid)” option and data from the quadruplicate spots were merged (using median log2 ratios). The “limma eBayes mod t-test (within group)” was applied to test for significant spots and the medians of the ratios across triplicate arrays from each time point were then calculated. Spots with p values <0.05 and fold changes less than −2 or greater than +2 were retained for further analysis. Significance analysis of microarrays (SAM; Tusher et al. 2001) was also performed for each set of triplicate arrays in order to select differentially expressed genes that were statistically significant.
Gene Ontology (GO) analysis (Ashburner et al. 2000) was performed using MatLab® (v. 7.0, The MathWorks, Natick, MA, USA) functions that emulate the functionality of GO::TermFinder (Boyle et al. 2004; Flight and Wentzell 2009). GO terms for 3,927 of the 9,277 unique genes on the microarray were obtained from http://www.pleurogene.ca. Two types of analysis were conducted. The first examined the genes at each time point (D2, D13, D23, and D33) that were found to be differentially expressed with a p value <0.05 in comparison to all the genes that passed filtering for the time point. The second analysis examined the low p value genes that passed filtering at three or more time points and compared them with all the genes that passed filtering at three or more time points. Multiple testing correction was implemented using the false discovery rate (FDR), and significant GO terms were those with an FDR <5%. The analysis was carried out using GO terms from biological process (P), molecular function (F), and cellular component (C).
Quantitative Real-Time PCR Experiments
First-strand cDNA was prepared from 1 µg of total RNA from each of the samples used for microarray analysis using the Superscript III First-Strand Synthesis Super Mix (Invitrogen). Primers for candidates to be validated: glutathione S-transferase (gstA1), DNA polymerase delta subunit 3 (polD3), replication protein A3 (rpa3), titin (ttn1), and peripherin (prph) and two housekeeping genes elongation factor 1A1 (ef1A1) and ribosomal protein S4 (rps4) were designed based on the Atlantic halibut expressed sequence tag (EST) corresponding to the microarray spot, using PrimerQuestSM software (http://www.idtdna.com/Scitools/Applications/Primerquest/) and synthesized by Integrated DNA Technologies (Toronto, ON, Canada). Amplicon sizes were approximately 140–160 bp and the optimum T m was 55°C (Table 2). All quantitative real-time polymerase chain reactions (qRT-PCR) were performed in a 20-µl total reaction volume (18 µl master mix and 2 µl PCR product/cDNA template). The master mix contained 6.4 µl H2O, 0.8 µl of each primer (0.4 µM final concentration), and 10.0 µl of the SYBR Green Mix (Roche Applied Science, Laval, PQ, Canada). The following cycling conditions were used: (1) denaturation, 5 min at 95°C; (2) amplification repeated 40 times, 10 s at 95°C, 10 s at 55°C, and 15 s at 72°C with ramp rate of 4.4°C/s, 2.2°C/s, and 4.4°C/s, respectively; (3) melting curve analysis, 1 min at 95°C and 1 min at 55°C with ramp rate of 4.4 and 2.2°C/s, respectively, then up to 95°C at a rate of 0.1°C/s; (4) cooling, 10 s at 40°C with ramp rate of 2.2°C/s. Each sample was analyzed in duplicate and reactions were performed in a Light Cycler (Roche Applied Science). Crossing point values were compared and converted to fold differences by the relative quantification method using the Relative Expression Software Tool (REST©) 384 v. 2 (Pfaffl et al. 2002) with ef1A1 and rps4 as the reference genes.
Results
Microdiet and Larval Performance
The microdiet consisted of particles of 50–200 µm that were of an irregular shape (Fig. 2). The diet sank relatively quickly to the bottom of the tanks and sometimes clouded the tanks even with regular cleaning. In a visual comparison of surface area of the fish at the conclusion of the trial, the control fish were 100% larger when compared to the fish fed the microdiet (Fig. 3). Measurements of weight, length, and width also showed significant (ANOVA; p < 0.05 or 0.001) differences in growth with the exception of the trial 3 fish and the first sampling point in trial 1 (Fig. 4). The experimental fish appeared to be developmentally delayed; by the end of the trial, eye migration in the control group was completed but fish in the experimental groups had either not yet started eye migration or had not completed it. Differences in pigmentation were also apparent; in the control tank, <25% of fish were malpigmented, whereas in the experimental tanks, 75% were malpigmented. Of these, 25% were nonpigmented and 50% were abnormally pigmented (“orange” fish). The bodies of these “orange” fish were light brown to orange and their eyes were a lighter hue than the normal black seen in the control fish. Mortalities varied between tanks; however, differences in mortalities between treatments were insignificant (ANOVA; p > 0.05). Mean mortalities were 32 (SD = 1.4), 46.5 (SD = 30.4), and 44 (SD = 0) for trials 1, 2, and 3, respectively. In the control tank, which contained many more fish than the experimental tanks, there were only 20 mortalities by the end of the trial.

Freeze-dried microencapsulated diet produced as described in the “Methods” section. Scale bar (dots) represents 500 µm

Phenotypic differences between experimental and control fish at the conclusion of the trial. a Trial group 2 transferred at 32 days post first feeding; b trial group 3 transferred at 43 days post first feeding; c control. Note the size difference and pigmentation. Larvae are not available from trial group 1 transferred at 20 days post first feeding as they were all required for RNA extraction due to their small size

Morphometrics of experimental and control fish from trials 1, 2, and 3 at different sampling times. a Weight, b length, c width. Mean values with standard errors are shown. *p < 0.05; **p < 0.001
Feed Consumption
The percentage of filled intestine as determined by image analysis of sections of gut from the larval samples is shown in Table 3. For all three trials, there was a significant difference between the control and experimental fish for the first 10 days with the former exhibiting a full gut and the latter only a partially filled gut (Table 3 and Fig. 5). However, for the larvae in trials 1 and 2, the experimental larvae appear to have adjusted to the microdiet after time as there was no significant difference in the amount of gut filled with food compared to control larvae at the later sampling times.

a Representative cross-section of the intestine of larval Atlantic halibut transferred from trial 1 and sampled 2 days after the introduction of the microdiet. b Representative cross-section of the intestine of larval Atlantic halibut from the production tank at the same time. Images are representatives of 16 sections analyzed for each of four to six individuals per sampling time. Scale bar is 200 µ. Note the difference in food contents of the gut
Gene Expression Changes
Of the 39,936 spots on the Atlantic halibut microarray, between 3,600 and 8,900 passed quality control for each array (Table 4). After calculating the median log2 ratios of the quadruplicate spots, approximately 2,000–4,000 data points were retained from each array. Following similar analysis of the triplicate arrays, approximately 1,500–2,300 spots were detected for each time point. In order to restrict our analysis to the highest quality data, only spots detected on two or more of the arrays were retained from each of the four time points. Of these, approximately 250–300 had a p value of <0.05 at each of the four time points and approximately two thirds were greater than twofold upregulated or downregulated. Fifty-eight genes were in common among the significantly differentially regulated genes in three or more time points (Tables 5 and 6). The gene showing similarity to trypsin was most closely related to a trypsin from sponge and the signal probably arose from either the Artemia live prey or, less likely, a component of the microdiet. For this reason, it was excluded from further analysis, as were those with no annotation.
SAM generated gene lists containing genes only present in all three of the replicate arrays. For microarrays corresponding to larvae sampled at D2, D13, D23, and D33 after transfer, there were 37, 54, 80, and 65 genes, respectively, that were significant by SAM. All of these genes were also present in the ArrayPipe results with the exception of three genes (23998—unknown, 22338—similar to ubiquinol–cytochrome c reductase complex 11-kDa protein, and 33342—similar to collagen type X, alpha 1) that were present only in the D23 SAM list and one gene (25531—unknown) that was present only in the D33 SAM list.
The differentially expressed genes that were present in gene lists from at least three out of the four time points could be grouped into five main classes according to their functions: metabolism and biosynthesis, protein trafficking, structural proteins, stress proteins, and those involved in cell division and proliferation (Tables 5 and 6). GO term analysis showed that the majority of these 26 annotated genes were significantly enriched in GO terms corresponding to metabolic processes when compared to all of the genes that passed filtering (Table 7). When the differentially expressed genes were compared at each of the four sampling times (Table 8), significant enrichment in GO terms associated with metabolic processes was evident at D2 after transfer, but at D23, the enrichments were associated with ion transport and muscle physiology. No significant enrichments were seen at D13 or D33.
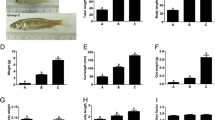
Five genes-of-interest were selected for verification by qRT-PCR: two genes that were upregulated at all time points (gstA1 and prph) and three genes that were downregulated at all time points (polD3, rpa3, and ttn1). The choice of housekeeping genes ef1A1 and rps4 as reference genes was based on their apparent stability throughout Atlantic halibut larval development (Infante et al. 2008). Although the magnitude of the changes in the expression of the genes-of-interest was not the same as in the microarray experiments, qRT-PCR analyses confirmed that glutathione S-transferase was significantly upregulated at D23 and peripherin at D2. DNA polymerase delta subunit 3 was not found to be significantly downregulated by qRT-PCR, whereas replication protein A3 and titin were significantly downregulated at D2 and D13.
Discussion
The microdiet appeared to be successfully consumed (assessed by percent of gut filled) after an initial period of adaptation. This occurred whether the diet was introduced at 20 or 32 days post first feeding. For the later introduction at 43 days post first feeding, the larvae did not consume the microdiet well (42.5% and 30% filled guts after 2 and 10 days, respectively). Since the larvae were switched to the commercial diet after 10 days, it was impossible to determine if they would have adapted to the microdiet.
The larvae appeared to prefer Artemia since the microdiet was not consumed initially as well. However, when Artemia was removed from the experiment at day 40 and the larvae were fed the microdiet only, the tanks were clear of uneaten particles. Histological examination of the gut of experimental larvae showed that the microdiet was physically digestible. Therefore, even with the exclusion of Artemia in their daily diet, the microparticles provided an alternate diet on which the larvae could survive. Fernández-Díaz et al. (1994), working with a protein microencapsulated diet, reported that fish larvae clearly prefer live prey to inert diets.
Mortalities of the experimental groups were somewhat higher than the control group, although this was not statistically significant. For example, trial 3, the last to be fed the experimental diet, had twice as many mortalities as the control group. This could be because they did not have time to adjust from the longer preceding period on the Artemia diet and were only given 1 day to adjust before they were switched to the microdiet. Trial 1 had a longer weaning period to switch from Artemia and adjust to the new microdiet and had the least mortalities. In fact, an early cofeeding period seems to be beneficial for accepting and processing inert diets, allowing better growth performance when weaning starts (Engrola et al. 2009; Curnow et al. 2006; Cañavate and Fernández-Díaz 1999; Rosenlund et al. 1997). In future studies, if the microdiet is administered, it may be beneficial to allow a few weeks for the fish to adjust to a new diet.
Lower feed consumption by larvae could have been due to the small particle size. The diet contained particles 50–200 µm in size; however, a more appropriate size range would have been 400 to 600 µm. The small particle size may have prevented the larvae from ingesting the diet due to its low visibility; it simply could have been ignored or not recognized as a food item. Alternatively, additional energy utilized to capture the small feed particles by larvae may have limited energy available for their growth and development.
Another explanation for the reduced growth in trial fish could be the density of the microdiet, which quickly sank to the bottom of the tank. Since halibut optimally feed within the water column, any particles that were not consumed in the brief time sinking through the water column collected on the bottom of the tank. Crucial micronutrients may also have leached from the microdiet prior to intake. If the microdiet had been suspended in the water column for a longer period of time, it may have been more available to the larvae. Changes to the physical properties of the microdiet to make it more buoyant should improve the results.
Although the diets were formulated to provide adequate level of nutrients, low bioavailability of certain nutrients as well as the leaching of nutrients may have limited growth of the experimental fish compared to fish fed live food organisms. As seen in Fig. 4, when fish were fed microdiet at early ages (trials 1 and 2), they exhibited reduced weight, length, and width compared to control fish. A similar observation was reported for sea bream larvae when their normal feed was substituted with the microdiet (Yúfera et al. 2005). The experimental halibut also appeared developmentally delayed, such as in eye migration and pigmentation, compared to the control group. This is likely due to small size, perhaps caused by nutrient deficiency or limited consumption of microdiet.
In order to study changes in larval gene expression, we compared larvae fed control diets with those fed microdiet. Since the control fish were reared in the production tank in optimal conditions throughout the trial, they were at a more advanced developmental stage than their experimental counterparts, especially in the later sampling times. This is evident in the morphometric data for trial 1 presented in Fig. 4. Although this may be seen as a limitation in our study of gene expression changes, most larval feeding trials are conducted in this manner since, unlike with juveniles, it is difficult to separate larval development changes from treatment effects. Rather than compare gene expression changes sequentially over time in the control fish and corresponding sequential changes in the experimental fish, we chose to compare gene expression changes in the control fish relative to the experimental fish at each sampling time with the caveat that developmental differences may preclude absolutely accurate comparisons. Every precaution was taken by the experienced hatchery staff to minimize handling stress.
Our previous microarray study of sequential development of Atlantic halibut larvae under control conditions indicated that the major gene expression changes seen between larvae at days 21 and 64 post hatching were related to digestion and muscle development (Douglas et al. 2008). Similar results were obtained in a comparison of sea bass larval development where genes involved in muscle development were expressed at later stages of development (31–43 days post hatching) and those involved in energy production were expressed early in development (Darias et al. 2008). In sea bream, genes involved in energy metabolism are also expressed early in development (Sarrapoulou et al. 2005). As discussed below, the changes we saw in response to the introduction of microdiet were mainly involved in metabolism, eye development, pigmentation, and tissue structure and gave an indication of subtle underlying physiological changes responsible for the growth and phenotypes observed.
Our analysis focused on significantly differentially expressed genes that we could track across at least three of the four time points. Of the 58 genes that satisfied these criteria, 31 were for unknown genes or ESTs that could not be annotated. Several of these unannotated genes showed very high upregulation (>20-fold) or were effectively shut off (<0.005-fold); their identification may reveal interesting processes that are sensitive to constituents of the microdiet, starvation, or stress. In fact, two of the genes that were similar to Platichthys flesus cDNA (Spot IDs 24743 and 26900) were from a liver cDNA library that had been selected for genes induced by chemical pollutants (Williams et al. 2006) and may be general stress proteins that are also induced by dietary stress.
Most of the genes involved in metabolism and biosynthesis were upregulated in the fish fed microdiet compared to control diet. GO analysis revealed an association of the genes with metabolic processes that were enriched particularly at D2 after transfer. This may reflect their attempt to adapt to marginal nutrient intake from the microdiet needed for optimum growth. Two glycolytic enzymes, aldolase A fructose-bisphosphate and glyceraldehyde-3-phosphate dehydrogenase, were upregulated 2 days after switching to the microdiet, indicating that the larvae were attempting to derive energy from glucose stores. Glutamine-fructose-6-phosphate transaminase 2 is an isozyme of glutamine-fructose-6-phosphate transaminase 1 (Gfpt1), the rate-limiting enzyme in glucosamine-6-phosphate biosynthesis, an important pathway for cellular glucose sensing. It is expressed throughout the central nervous system, especially in the spinal cord in humans and mice and has been implicated in type II diabetes (Zhang et al. 2004). Interestingly, the isozyme Gfpt1 promotes ontogenetic melanocyte darkening (Yang et al. 2007). In our study, Gfpt2 was moderately upregulated in fish fed the microdiet. A gene encoding transmembrane protein 33, which is similar to DB83 protein, a component of the melanosome proteome (Chi et al. 2006), was also highly upregulated at all three time points for which we have data. Since malpigmentation is such an issue in the aquaculture of Atlantic halibut, possible roles of these proteins in flatfish melanocyte development should be further examined. Two enzymes involved with lipid metabolism, 1-acylglycerol-3-phosphate O-acyltransferase and cGMP-inhibited phosphodiesterase 3A, were upregulated at all time points tested. cGMP-inhibited phosphodiesterase 3A plays various roles in vertebrates including procoagulant activity, lipid metabolism, and signal transduction mainly through the regulation of intracellular cAMP levels (Zhang and Colman 2007), whereas 1-acylglycerol-3-phosphate O-acyltransferase is involved in regulating phospholipid biosynthesis (Gale et al. 2006).
Two genes encoding proteins involved in trafficking through the Golgi and endoplasmic reticulum (ER) were highly upregulated (approximately 20-fold) in fish fed the microdiet. Golgi vesicular membrane trafficking protein p18 (Bet1) is involved in SNARE interactions in vesicular transport (Joglekar et al. 2003) and Golgi membrane protein SB140 (yip1) is involved in the regulation of ER–Golgi traffic at the level of ER exit sites (Barrowman et al. 2003). The increased expression of these two genes may reflect a disruption in the fishes' ability to traffic newly synthesized proteins efficiently through the ER.
Genes for a number of structural proteins were also differentially regulated. Two forms of collagen, Ia3, which is found in the skin, and IXa1, which may participate in electrostatic interactions with polyanionic glycosaminoglycans in cartilage, were both downregulated. The gene encoding the titin isoform N2-B was also downregulated. Titin is a scaffold for signaling proteins in muscle and is responsible for establishing and maintaining the structure and elasticity of sarcomeres in striated muscle (Steffen et al. 2007). Warm-temperature-acclimation-related 65-kDa protein contains hemopexin-like repeats which occur in vitronectin and some matrix metalloproteinase families such as collagenases, stromelysins, and other enzymes that metabolize the extracellular matrix. It may be important in the remodeling of the extracellular matrix and play an important role during early development (Nakaniwa et al. 2005). It was upregulated at D13 and D23 but had decreased by D33. Enrichment in GO terms associated with muscle physiology among the differentially expressed genes at D23 (Table 8) confirms the effect seen on structural proteins. Taken together, the downregulation of structural proteins would result in the lower growth seen in the experimental fish compared to the control fish.
Genes for three proteins involved in eye development were upregulated. Crystallin betaA2-2 participates in ocular development and contributes to the transparency and refractive properties of the eye lens (Wang et al. 2008). It was highly upregulated at D13 following the introduction of the microdiet. A gene for a protein similar to es1 protein was also upregulated at this time. Zebrafish es1 is expressed specifically in adult photoreceptor cells and appears to be important for maintaining normal retina structure and function (Chang and Gilbert 1997). Peripherin 1 (also known as plasticin) is a neuronal intermediate filament protein that constitutes part of the cytoskeleton and is important for retinal tissue regeneration (Cameron et al. 2005; Glasgow et al. 1992). It is a marker of anterior neural induction and is expressed most strongly in the brain of the tailbud stage Xenopus embryos (Sharpe et al. 1989). In our study, it was highly upregulated in larvae fed the microdiet throughout all four time points. As discussed above, transmembrane protein 33 (DB83 homolog) is found in melanosomes and may impact eye pigmentation. Also, lens epithelium-derived growth factor and basic transcription factor 3 (BTF3a) have been shown to be differentially expressed during retinal development in mice (see below). These genes associated with eye development may have contributed to some of the aberrations in the eyes of larvae fed the microdiet.
The gene for glutathione S-transferase A1 was highly upregulated in larvae fed microdiet. This protein is responsible for detoxification of reactive electrophilic compounds, including intracellular metabolites produced endogenously as a result of cellular oxidative processes, as well as exogenous compounds such as drugs, pollutants, and pesticides. Two similar gstA genes have been isolated from a related flatfish, Pleuronectes platessa (Leaver et al. 1997); it was suggested that these particular glutathione S-transferases are involved in the detoxification of fatty acid metabolites, particularly those arising from polyunsaturated fatty acid oxidation. It is possible that oxidation products or some of the artificial components of the microdiet induced the expression of this gene. Hepcidin was highly upregulated at D13 but downregulated at other time points. Hepcidin is a bifunctional peptide involved in both innate immunity and iron homeostasis and is highly expressed in response to bacterial infection and iron overload (Ganz 2006). Interestingly, its expression was reduced in response to growth hormone transgenesis in Atlantic salmon (Rise et al. 2006), which would result in increased availability of iron for hemoglobin biosynthesis but possibly compromised innate immunity. The dramatic upregulation of hepcidin at D13 may reflect the stresses affecting the larvae in response to the microdiet and subsequent adaptation to the microdiet (with concomitant decrease in hepcidin expression) at later time points.
Most of the genes involved in replication, translation, and cell proliferation that we identified as differentially expressed were downregulated in fish fed the microdiet relative to the controls. These included DNA polymerase delta subunit 3 and replication protein A3, 14 kDa, a nuclear single-stranded DNA binding protein that appears to be involved in all aspects of DNA metabolism including replication, recombination, and repair. Also downregulated were genes encoding transmembrane protein 85 (proliferation-inducing gene 17) and BTF3a transcription factor, which is essential for post implantation embryonic development in mice (Deng and Behringer 1995) and is also differentially expressed in the outer neuroblastic layer of the developing retina (Blackshaw et al. 2004). The gene encoding a protein similar to mitotic spindle assembly checkpoint protein MAD2A, which prevents progression of the cell cycle until all chromosomes are properly aligned at the metaphase plate (Howell et al. 2000), was also downregulated as was the gene for lens epithelium-derived growth factor, a chromatin-associated protein that has been implicated in transcriptional regulation and that provides cellular protection against stress by transactivating stress-associated genes. This protein also facilitates cellular protection against ethanol stress and plays a role in retinoic acid production (Fatma et al. 2004), which is required for the normal growth and maintenance of many cell types, including lens epithelial cells.
Two genes encoding regulatory proteins were upregulated in larvae fed microdiet. The “zinc finger, A20 domain containing 2 protein” bears the inhibitor of cell death-like zinc finger domains, which mediate the self-association of A20 and subsequent IL-1-induced NF-kappaB activation (Heyninck and Beyaert 1999). Its moderate upregulation (fourfold to sevenfold) could impact the expression of immune/stress-related genes that are affected by NF-kappaB. The gene encoding SFRS protein kinase 1, a serine/arginine (SR) protein kinase specific for the SR-rich domain family of splicing factors was massively upregulated (33-fold) at D13 and moderately upregulated at the other time points (fourfold to eightfold). It is thought to play a role in the regulation of both constitutive and alternative splicing by regulating phosphorylation and intracellular localization of splicing factors (Ma et al. 2008). The gene for eukaryotic translation elongation factor 1B2, which participates in the elongation cycle of protein biosynthesis, was only moderately affected.
Conclusion
Although not completely successful, the microdiet showed promise as a larval feed in Atlantic halibut. It was an improvement from previous experiments because the particles were readily consumed after a period of adaptation and provided sufficient nutrients at the maintenance level for larvae. Similar to other microdiets, growth was limited in the fish. Insights into the reasons for growth limitation and malpigmentation have been revealed through studies of gene expression changes. Adjustments to the method of feeding the microdiet, such as suspending the particles in the water column using aeration, may improve results. Changing the physical properties and formulation of the microdiet, such as a larger particle size and lower density, may show an improvement in the overall health and quality of larval halibut.
References
Applebaum S (1985) Rearing of Dover sole, Solea solea (L.), through its larval stages using artificial diets. Aquaculture 49:209–221
Ashburner M, Ball CA, Blake JA, Botstein D, Butler H, Cherry JM, Davis aP, Dolinski K, Dwight SS, Eppig JT, Harris MA, Hill DP, Issel-Tarver L, Kasarskis A, Lewis S, Matese JC, Richardson JE, Ringwald M, Rubin GM, Sherlock G (2000) Gene ontology: tool for the unification of biology. The Gene Ontology Consortium. Nat Genet 25:25–29
Barrowman J, Wang W, Zhang Y, Ferro-Novick S (2003) The Yip1p.Yif1p complex is required for the fusion competence of endoplasmic reticulum-derived vesicles. J Biol Chem 278:19878–19884
Bell JG, Mcevoy LA, Estevez A, Shields RJ, Sargent JR (2003) Optimising lipid nutrition in first-feeding flatfish larvae. Aquaculture 227:211–220
Berg L (1997) Commercial feasibility of semi-intensive larviculture of Atlantic halibut (Hippoglossus hippoglossus L.). Aquaculture 155:333–340
Blackshaw S, Harpavat S, Trimarchi J, Cai L, Huang H, Kuo WP, Weber G, Lee K, Fraioli RE, Cho SH, Yung R, Asch E, Ohno-Machado L, Wong WH, Cepko CL (2004) Genomic analysis of mouse retinal development. PLoS Biol 2:1411–1431
Blanchard B, Scarratt D (2002) Finely-tuned hatchery the key to halibut success. Hatch Int 3:9–13
Boyle EI, Weng S, Gollub J, Jin H, Botstein D, Cherry JM, Sherlock G (2004) GO::TermFinder—open source software for accessing Gene Ontology information and finding significantly enriched Gene Ontology terms associated with a list of genes. Bioinformatics 20:3710–3715
Cameron DA, Gentile KL, Middleton FA, Yurco P (2005) Gene expression profiles of intact and regenerating zebrafish retina. Mol Vis 11:775–191
Cañavate JP, Fernández-Díaz C (1999) Influence of co-feeding larvae with live and inert diets on weaning the sole Solea senegalensis onto commercial dry feeds. Aquaculture 17:255–263
Chang H, Gilbert W (1997) A novel zebrafish gene expressed specifically in the photoreceptor cells of the retina. Biochem Biophys Res Commun 237:84–89
Chi A, Valencia JC, Hu ZZ, Watabe H, Yamaguchi H, Mangini NJ, Huang H, Canfield VA, Cheng KC, Yang F, Abe R, Yamagishi S, Shabanowitz J, Hearing VJ, Wu C, Appella E, Hunt DF (2006) Proteomic and bioinformatic characterization of the biogenesis and function of melanosomes. J Proteome Res 5:3135–3144
Curnow J, King J, Bosmans J, Kolkovski S (2006) The effect of reduced Artemia and rotifer use facilitated by a new microdiet in the rearing of barramundi Lates calcarifer (BLOCH) larvae. Aquaculture 257:204–213
Darias MJ, Zambonino-Infante JL, Hugot K, Cahu CL, Mazurais D (2008) Gene expression patterns during the larval development of European sea bass (Dicentrarchus labrax) by microarray analysis. Mar Biotechnol (NY) 10:416–428
Deng JM, Behringer RR (1995) An insertional mutation in the BTF3 transcription factor gene leads to an early postimplantation lethality in mice. Transgenic Res 4:264–269
Douglas SE, Knickle LC, Williams J, Flight RM, Reith ME (2008) A first generation Atlantic halibut microarray: application to developmental studies. J Fish Biol 72:2393–2408
Engrola S, Figueira L, Conceiçao LEC, Gavaia PJ, Ribeiro L, Dinis MT (2009) Co-feeding in Senegalese sole larvae with inert diet from mouth opening promotes growth at weaning. Aquaculture 288:264–272
Evjemo JO, Reitan KI, Olsen Y (2003) Copepods as live food organisms in the larval rearing of halibut larvae (Hippoglossus hippoglossus L.) with special emphasis on the nutritional value. Aquaculture 227:191–210
Fatma N, Kubo E, Chylack LTJ, Shinohara T, Akagi Y, Singh DP (2004) LEDGF regulation of alcohol and aldehyde dehydrogenases in lens epithelial cells: stimulation of retinoic acid production and protection from ethanol toxicity. Am J Physiol Cell Physiol 287:C508–C516
Fernández-Díaz C, Pascual E, Yúfera M (1994) Feeding behavior and prey size selection of gilthead seabream, Sparus aurata, larvae fed on inert and live food. Mar Biol 118:323–328
Flight RM, Wentzell PD (2009) Potential bias in GO::TermFinder. Brief Bioinform 10:289–294. doi:10.1093/bib/bbn054
Gale SE, Frolov A, Han X, Bickel PE, Cao L, Bowcock A, Schaffer JE, Ory DS (2006) A regulatory role for 1-acylglycerol-3-phosphate-O-acyltransferase 2 in adipocyte differentiation. J Biol Chem 281:11082–11089
Ganz T (2006) Hepcidin—a peptide hormone at the interface of innate immunity and iron metabolism. Curr Top Microbiol Immunol 306:183–198
Gara B, Shields RJ, Mcevoy L (1998) Feeding strategies to achieve correct metamorphosis of Atlantic halibut, Hippoglossus hippoglossus L., using enriched Artemia. Aquac Res 29:935–948
Glasgow E, Druger RK, Levine EM, Fuchs C, Schechter N (1992) Plasticin, a novel type III neurofilament protein from goldfish retina: increased expression during optic nerve regeneration. Neuron 9:373–381
Hamre K, Næss T, Espe M, Holm JC, Lie Ø (2001) A formulated diet for Atlantic halibut (Hippoglossus hippoglossus, L.) larvae. Aquac Nutr 7:123–132
Hamre K, Opstad I, Espe M, Solbakken J, Hemre G-I, Pittman K (2002) Nutrient composition and metamorphosis sucess of Atlantic halibut (Hippoglossus hippoglossus, L.) larvae fed natural zooplankton or Artemia. Aquac Nutr 8:139–148
Hamre K, Øfsti A, Næss T, Nortvedt R, Holm JC (2003) Macronutrient composition of formulated diets for Atlantic halibut (Hippoglossus hippoglossus, L.) juveniles. Aquaculture 227:233–244
Hamre K, Moren M, Solbakken J, Opstad I, Pittman K (2005) The impact of nutrition on metamorphosis in Atlantic halibut (Hippoglossus hippoglossus L.). Aquaculture 250:555–565
Heyninck K, Beyaert R (1999) The cytokine-inducible zinc finger protein A20 inhibits IL-1-induced NF-kappaB activation at the level of TRAF6. FEBS Lett 442:147–150
Hokamp K, Roche FM, Acab M, Rousseau M-E, Kuo B, Goode D, Aeschliman D, Bryan J, Babiuk LA, Hancock REW, Brinkman FSL (2004) ArrayPipe: a flexible processing pipeline for microarray data. Nucleic Acids Res 32:W457–459
Howell BJ, Hoffman DB, Fang G, Murray AW, Salmon ED (2000) Visualization of Mad2 dynamics at kinetochores, along spindle fibers, and at spindle poles in living cells. J Cell Biol 150:1233–1250
Infante C, Matsuoka MP, Asensio E, Cañavate JP, Reith M, Manchado M (2008) Selection of housekeeping genes for gene expression studies in larvae from flatfish using real-time PCR. BMC Mol Biol 9:28
Joglekar aP, Xu D, Rigotti DJ, Fairman R, Hay JC (2003) The SNARE motif contributes to rbet1 intracellular targeting and dynamics independently of SNARE interactions. J Biol Chem 278:14121–14133
Jones D, Munford J, Gabbott P (1974) Microcapsules as artificial food particles for aquatic filter feeders. Nature 247:233–235
Kolkovski S, Tandler A, Izquierdo MS (1997) Effects of live food and dietary digestive enzymes on the efficiency of microdiets for seabass (Dicentrarchus labrax) larvae. Aquaculture 148:313–322
Langdon C (2003) Microparticle types for delivering nutrients to marine fish larvae. Aquaculture 227:259–275
Leaver MJ, Wright J, George SG (1997) Structure and expression of a cluster of glutathione S-transferase genes from a marine fish, the plaice (Pleuronectes platessa). Biochem J 321:405–412
Lewis LM, Lall SP, Witten PE (2004) Morphological descriptions of the early stages of spine and vertebral development in hatchery-reared larval and juvenile Atlantic halibut (Hippoglossus hippoglossus). Aquaculture 241:47–59
Lewis-Mccrea LM, Lall SP (2007) Effects of moderately oxidized dietary lipid and the role of vitamin E on the development of skeletal abnormalities in juvenile Atlantic halibut (Hippoglossus hippoglossus). Aquaculture 262:142–155
Lie Ø, Lambertsen G (1985) Digestive lipolytic enzymes in cod (Gadus morhua): fatty acid specificity. Comp Biochem Physiol 80B:447–450
Ma CT, Velazquez-Dones A, Hagopian JC, Ghosh G, Fu XD, Adams JA (2008) Ordered multi-site phosphorylation of the splicing factor ASF/SF2 by SRPK1. J Mol Biol 376:55–68
Mangor-Jenson A, Harboe T, Shields RJ, Gara B, Naas KE (1998) Atlantic halibut, Hippoglossus hippoglossus L., larvae cultivation literature, including a bibliography. Aquac Res 29:857–886
Murray HM, Gallant JW, Johnson SC, Douglas SE (2006) Cloning and expression analysis of three digestive enzymes from Atlantic halibut (Hippoglossus hippoglossus) during early development: predicting gastrointestinal functionality. Aquaculture 252:394–408
Naess T, Hamre K, Holm JC (2001) Successful early weaning of Atlantic halibut (Hippoglossus hippoglossus L.) in small shallow raceway systems. Aquac Res 32:163–168
Naess T, Lie O (1998) A sensitive period during first feeding for the determination of pigmentation pattern in Atlantic halibut, Hippoglossus hippoglossus L., juveniles: the role of diet. Aquac Res 29:925–934
Nakaniwa M, Hirayama M, Shimizu A, Sasaki T, Asakawa S, Shimizu N, Watabe S (2005) Genomic sequences encoding two types of medaka hemopexin-like protein Wap65, and their gene expression profiles in embryos. J Exp Biol 208:1915–1925
O.-Jordal a-E, Torstensen BE, Tsoi SC, Tocher DR, Lall S, Douglas SE (2005) Small scale cDNA microarray analysis of expression of genes for lipid metabolism in liver of Atlantic salmon (Salmo salar L.)—effect of dietary rapeseed oil replacement. J Nutr 135:2355–2361
Önal U, Langdon C (2005) Perfomance of zein-boud particles for delivery of riboflavin to early fish larvae. Aquac Nutr 11:351–258
Pfaffl MW, Horgan GW, Dempfle L (2002) Relative expression software tool (REST©) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res 30:e36
Rise ML, Douglas SE, Sakhrani D, Williams J, Ewart KV, Rise M, Davidson WS, Koop BF, Devlin RH (2006) Multiple microarray platforms utilized for hepatic gene expression profiling of GH transgenic coho salmon with and without ration restriction. J Mol Endocrinol 37:259–82
Ronnestad I, Helland S, Lie Ø (1998) Feeding Artemia to larvae of Atlantic halibut (Hippoglossus hippoglossus L.) results in lower larval vitamin A content compared with feeding copepods. Aquaculture 165:159–164
Rosenlund G, Stoss J, Talbot C (1997) Co-feeding marine fish larvae with inert and live diets. Aquaculture 155:183–191
Sarrapoulou E, Kotoulas G, Power DM, Geisler R (2005) Gene expression profiling of gilthead sea bream during early development and detection of stress-related genes by the application of cDNA microarray technology. Physiol Genomics 23:182–191
Sharpe CR, Pluck A, Gurdon JB (1989) XIF3, a Xenopus peripherin gene, requires an inductive signal for enhanced expression in anterior neural tissue. Devel 107:701–714
Solbakken JS, Berntssen MHG, Norberg B, Pittman K, Hamre K (2002) Different iodine and thyroid levels between Atlantic halibut larvae fed wild zooplankton or Artemia from first exogenous feeding until post metamorphosis. J Fish Biol 61:1345–1362
Steffen LS, Guyon JR, Vogel ED, Howell MH, Zhou Y, Weber GJ, Zon LI, Kunkel LM (2007) The zebrafish runzel muscular dystrophy is linked to the titin gene. Dev Biol 309:180–192
Tusher V, Tibshirani R, Chu C (2001) Significance analysis of microarrays applied to ionizing radiation response. Proc Natl Acad Sci U S A 98:5116–5121
Vilhelmsson O, Martin SAM, Medale F, Kaushik SJ, Houlihan DF (2004) Dietary plant-protein substitutes affects hepatic metabolism in rainbow trout (Oncorhynchus mykiss). Br J Nutr 92:71–80
Wang H, Kesinger JW, Zhou Q, Wren JD, Martin G, Turner S, Tang Y, Frank MB, Centola M (2008) Identification and characterization of zebrafish ocular formation genes. Genome 51:222–235
Williams TD, Diab aM, George SG, Godfrey RE, Sabine V, Conesa A, Minchin SD, Watts PC, Chipman JK (2006) Development of the GENIPOL European flounder (Platichthys flesus) microarray and determination of temporal transcriptional responses to cadmium at low dose. Environ Sci Technol 40:6479–6488
Yang C-T, Hindes AE, Hultman KA, Johnson SL (2007) Mutations in gfpt1 and skiv2l2 cause distinct stage-specific defects in larval melanocyte regeneration in zebrafish. PLoS Genet 3:886–900
Yúfera M, Fernández-Diaz C, Pascual E, Sarasquete C, Moyano FJ, Diaz M, Alarcon FJ, Garcia-Gallego M, Parra G (2000) Towards an inert diet for first-feeding gilthead seabream Sparus aurata L. larvae. Aquac Nutr 6:143–152
Yúfera M, Fernández-Diaz C, Pascual E (2005) Food microparticles for larval fish prepared by internal gelation. Aquaculture 248:253–262
Yúfera M, Kolkovski S, Fernández-Diaz C, Rinchard J, Lee KJ, Dabrowski K (2003) Delivering bioactive compounds to fish larvae using microencapsulated diets. Aquaculture 227:277–291
Yúfera M, Pascual E, Fernández-Diaz C (1999) A highly efficient microencapsulated food for rearing early larvae of marine fish. Aquaculture 177:249–256
Zhang H, Jia Y, Cooper JJ, Hale T, Zhang Z, Elbein SC (2004) Common variants in glutamine:fructose-6-phosphate amidotransferase 2 (GFPT2) gene are associated with type 2 diabetes, diabetic nephropathy, and increased GFPT2 mRNA levels. J Clin Endocrinol Metab 89:748–755
Zhang W, Colman RW (2007) Thrombin regulates intracellular cyclic AMP concentration in human platelets through phosphorylation/activation of phosphodiesterase 3A. Blood 110:1475–1482
Acknowledgements
This project entitled: PLEUROGENE: Flatfish genomics—Enhancing commercial culture of Atlantic halibut and Senegal sole, was funded by the Genome Canada–Genome España joint program. Dr. Vindhya Mohindra was supported by an Overseas Associateship from the Department of Biotechnology, India. We thank Sean Tibbetts for the initial preparation of the microdiet, David O'Neil of the IMB microscopy laboratory for the electron micrographs of the microdiet, Jason Williams of the IMB microarray laboratory for the assistance with the microarrays, and the staff of Scotian Halibut Ltd. for the sampling. The critical review of this manuscript by Darrin Reid is gratefully acknowledged. All microarray data have been deposited at the Gene Expression Omnibus with accession number GSE13024. This is NRC publication number 2009-42827.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Murray, H.M., Lall, S.P., Rajaselvam, R. et al. Effect of Early Introduction of Microencapsulated Diet to Larval Atlantic Halibut, Hippoglossus hippoglossus L. Assessed by Microarray Analysis. Mar Biotechnol 12, 214–229 (2010). https://doi.org/10.1007/s10126-009-9211-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10126-009-9211-4