
Abstract
The aim of this study was to culture sponge juveniles from larvae. Starting from larvae we expected to enhance the survival and growth, and to decrease the variation in these parameters during the sponge cultures. First, settlement success, morphological changes during metamorphosis, and survival of Dysidea avara, Ircinia oros, Hippospongia communis, under the same culture conditions, were compared. In a second step, we tested the effects of flow and food on survival and growth of juveniles from Dysidea avara and Crambe crambe. Finally, in a third experiment, we monitored survival and growth of juveniles of D. avara and C. crambe transplanted to the sea to compare laboratory and field results. The results altogether indicated that sponge culture from larvae is a promising method for sponge supply and that laboratory culture under controlled conditions is preferred over sea cultures in order to prevent biomass losses during these early life stages.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Sponges produce secondary metabolites of pharmaceutical interest (Blunt et al., 2005; Sipkema et al., 2005a). The supply of these bioactive metabolites in sufficient amounts for preclinical and clinical assays is an unsolved problem (Fusetani, 2000; Procksh et al., 2003; Sipkema et al., 2005b). Chemical synthesis is too expensive and wild sponge collection has a negative impact on the environment and endangers the sustainable use of marine resources (according to convention on biological diversity rules). Sponge cultures have been assayed since the beginning of the last century (Moore, 1910); more recently, besides mariculture (in situ sponge culture; Pronzato et al., 1999; Duckworth and Battershill, 2003; Mendola, 2003), three methods for culturing sponges under controlled conditions have been used: explant culture (Osinga et al., 2001; de Caralt et al., 2003; Hoffmann et al., 2003; Nickel and Brümmer, 2003), primmorphs (Müller et al., 1999; Zhang et al., 2003), and cells (Pomponi and Willoughby, 1994; Sipkema et al., 2003; De Rosa et al., 2003). However, most of these methods have encountered drawbacks with regard to survival and contamination (especially cell cultures) and/or growth rates (explants and primmorphs), and more research is required to make them suitable for scaling up sponge biomass production.
In explant cultures, we make use of the regenerative capacity of sponges (Simpson, 1984). It has been shown, however, that explants have very slow and variable grow rates (Osinga et al., 1999; de Caralt et al., 2003; Sipkema et al., 2006). The large variability in growth rates of explants could be due to the different ages of the sponges from which the explants were made, because slow and variable growth has also been reported for adult sponges in the sea (Turon et al., 1998; Garrabou and Zabala, 2001). To optimize and standardize culture conditions, we need to prevent this variability by culturing sponges of the same age. However, age classes cannot be determined for adult sponges because their size is not necessarily related to their age. As a consequence, it is difficult to avoid variation in cultures of explants obtained from adult individuals.
In vitro cultures of sponges have been developed from adult individuals. Long-term continuously proliferating cell cultures have failed so far, and the maintenance of axenic cell lines (i.e., microbe-free) has proved difficult (Rinkevich, 2005). Continuous cell lines require proliferative stem cells. In adult sponges, the cells closer to stem cells are archeocytes, which are able to differentiate in several cell types (e.g., oocytes, sclerocytes, pinacocytes). Some authors have performed cell culture from archeocyte enriched cell fractions, which presented active DNA synthesis, but failed to obtain continuous cell proliferation because of an early cell death (Pomponi and Willoughby, 1994; Zhang et al., 2003).
Embryos are a source of totipotent and proliferating cells, and thus embryonic cells could be a good source from which to start cell culture. However, only one attempt to culture cells from sponge larvae has been reported. The results were promising, as the culture was more resistant to infections by microorganisms and survived longer than those from adult cells (Rinkevich et al., 1998).
By culturing sponges from larvae, we attempt to circumvent the three main constraints that are commonly found in cultures from adults. First, we expect higher growth rates during the juvenile stages of sponges, as this has been reported for small individuals (Garrabou and Zabala, 2001). Moreover, variability may be reduced, because culturing larvae warrants the same age for all the individuals (cohorts) and a similar behavior under culture. Second, larval survival in the laboratory is expected to be higher than at sea because of the absence of predators and competitors (Uriz et al., 1996a). Third, larval culture can be a source of more suitable starting material (embryonic cells) for the development of cell culture. Consequently, larval culture offers some advantages with respect to other assayed methods.
The sponge species for our study were Dysidea avara, Ircinia oros, Hippospongia communis, and Crambe crambe, which were selected because of the interest in the species for pharmaceutical applications and their abundance in the western Mediterranean. They produce several bioactive compounds with commercial value. D. avara produces avarol (Uriz et al., 1996b), a sesquiterpenoid antitumor (Müller et al., 1985) and antiviral compound (Sarin et al., 1987) that is a potent in vitro inhibitor of HIV-1 reverse transcriptase. (Loya and Hizi, 1990). I. oros produces a cyclic sesterpenoid (Cimino et al., 1972) with anti-inflammatory properties. H. communis has antifungal activity because of the production of untenospongin B (Rifai et al., 2004). Finally, C. crambe produces crambines and crambescidins, which are derivatives of a pentacyclic guanidine with anticancer and antiviral activities (Jares-Erijman et al., 1991; Balconi et al., 1995).
The general objective of this study was to culture sponges from larvae, which was never done before, in order to try to improve biomass supply for biotechnological purposes. The study was performed in the laboratory under several food and flow conditions, and at sea. Three experiments with different goals were conducted. The aim of the first experiment was to select the most adequate species for culturing among Dysidea avara, Ircinia oros, and Hippospongia communis. In this experiment, settling, metamorphosis, and juvenile survival in the laboratory were monitored. In the second experiment, we assayed larval culture of D. avara, the most successful species in the previous experiment, and C. crambe, already reported to have high survival (Uriz et al., 1998) under different environmental conditions. Finally, a third experiment was addressed to compare juvenile survival and growth of settlers of both species (D. avara and C. crambe) after transplanting them to the sea.
Materials and Methods
Collection, Maintenance of Sponge Larvae, and Experimental Procedures
For the first experiment, ripe individuals of I. oros, H. communis, and D. avara were collected from l’Escala (western Mediterranean Sea) and transferred to an open aquarium system (CMIMA-CSIC, Barcelona, Spain) at the beginning of June of 2003. I. oros and H. communis released larvae in mid-July, and D. avara released at the end of July (similar periods to those previously reported in sponges at sea; Mariani et al., 2005).
Swimming larvae of I. oros (N = 224), H. communis (N = 30), and D. avara (N = 230) were collected with a pipette and transferred to six-multiwell dishes, which were placed in aquaria filled with filtered (0.7 μm pore diameter) seawater at field temperature (20°C). After 24 h, the larvae started to settle spontaneously on the six-multiwell dish bottom and metamorphosed into juveniles after 5 to 7 days. We monitored settlement success, morphological changes, and survival of settlers in time.
For the second experiment, we selected D. avara, the most successful species in the previous experiment, and C. crambe, already reported to have high survival (Uriz et al., 1998).
Larvae of D. avara were taken after release from ripe sponges maintained in the open system aquarium (CMIMA) in July 2004, as in the previous experiment. Conversely, larvae of C. crambe were obtained by carefully tearing ripe individuals (Uriz et al., 1998) collected from the sea (Blanes, western Mediterranean). Larvae were placed in aquaria filled with filtered (0.7 μm pore diameter) seawater at field temperature (20°C). The aquaria bottom was covered by plastic (11 × 8 cm) sheets where larvae settled spontaneously. After settlement, juveniles of C. crambe (N = 50) and D. avara (N = 30) were submitted to the experimental conditions, in aquaria of Wageningen University (Sipkema et al., 2006). The variables measured were survival and growth.
Flow treatment consisted of unidirectional, constant flow of 5 cm/s, measured daily with a flowmeter (2031 H Series), and was compared against static condition. In this experiment living algae was used as a food.
Food treatment consisted in feeding the juveniles with 5 10 5 cells/ml of living algae versus feeding the juveniles with dried algae (equivalent carbon content). The alga used, as a source of food, was Phaeodactylum tricornutum (proven suitable in previous studies, Osinga et al., 2003; Sipkema et al., 2006). This treatment was conducted under the above-mentioned flow conditions.
The third experiment was also conducted on D. avara (N = 164) and C. crambe (N = 175). Larvae were released from ripe sponges (July 2005), settled on rigid plastic plates in the laboratory, and 1 week later transferred to the sea (western Mediterranean). Survival and growth were monitored as a function of time.
Morphology, Survival and Growth Rate Measurements
Morphological features, during metamorphosis and juvenile stages, were observed through a stereomicroscope. Survival and growth rates were monitored every 15 days by taking pictures through a stereomicroscope (in the laboratory) and with an underwater camera (in the field). Image analysis was performed to quantify increases in area of juveniles via NIH Image (public domain) software.
In the first experiment, settlement success was calculated as the percentage of larvae attached to the substratum after 1 week with respect the total number of swimming larvae.
Survival of juveniles was assessed in the three experiments as the percentage of juveniles that were alive on a given day. Death of juveniles was made evident by changes in colour, shape, and size (i.e., paler colour, rounder shape, and small size) ending in sponge disaggregation. To compare two survival curves, we used the Gehan’s Wilcoxon test statistics. For comparing more than two survival curves, we used the analysis Comparing Survival in Multiple Groups (Statistica software).
Growth rates were derived from changes in the sponge area with time (i.e., 15 days) by the equation:
Where A t and A t–1 are the sponge areas at time t and at time t–1, respectively (Turon et al., 1998; Garrabou and Zabala, 2001; Mariani et al., 2000). The changes in surface area are an acceptable estimation of growth when organisms mainly grow in two dimensions, as encrusting sponges do (Turon et al., 1998). Although the method did not take into account increases in thickness and thus the measured growth rates could underestimate true growth, the method was suitable here because juveniles were relatively flat and the measures obtained were used only for comparative purposes.
Differences in growth rates between treatments and species were analyzed statistically using a nonparametric version of the repeated-measures analysis of variance (ANOVAR). This method was used because data did not meet the circularity assumption (Mauchly’s sphericity test) required for parametric ANOVAR (Potvin et al., 1990; Von Ende, 1993). It is based on a permutation test by means of a two-level randomisation method (Manly, 1991; Turon et al., 1998). The whole series of data was randomized 4999 times (plus the observed one) to approximate the null hypothesis distribution of the sum of squares for each factor and their interaction, and then we examined how extreme were the observed values in this distribution. An effect was judged significant when the observed sum of squares was exceeded by less than 5% of the corresponding values in the randomization series. A modified version on the Turbopascal program used by Turon et al. (1998) was applied to perform the permutation tests.
Differences in growth rates on juveniles fed with either living or dried algae at the last point time of the culture were analyzed by a two-way ANOVA. Data normality and homogeneity were determined by a Kolmogorov-Smirnov test and the Levene’s test, respectively.
To compare the extent of variation in growth rates in adult individuals (Literature data) and in larvae cultures, the variation coefficients were calculated as a (standard error/ mean) × 100.
Results and Discussion
Larval Collection and Morphological Changes at Settling
Sponge larvae were obtained in the laboratory from ripe sponges: 224 larvae from I. oros, 30 from H. communis, and 424 from D. avara, and 325 from C. crambe. This method is the most effective to obtain sufficient larvae for reproducible experiments (i.e., enough replicates per treatment). Alternative methods such as collecting sponge larvae from the field are more complex because of the relatively short period of larval release and the fast larval dilution once released (Mariani et al., 2005). In addition, there are few studies about the period of larval release, which is species-specific and may vary depending on the geographical location (water temperature, hydrodynamism).
The larvae of Hippospongia communis, Ircinia oros, Dysidea avara (Figure 1A) and Crambe crambe (Figure 1B) are parenchymella that consist of a pseudostratified layer of flagellated cells surrounding an internal mass of cells (Boury-Esnault and Rützler, 1997). Larvae swam during a variable but short period (from 3 to 7 days in the laboratory) before settling. Usually, D. avara and C. crambe larvae settled sooner than H. communis and I. oros larvae. Larvae of the four species attached to the substrate through their posterior pole and became hemispherical. Settled larvae flattened as the cells spread on the substrate (growth marginal layer) and initiated metamorphosis. During metamorphosis, the few cell types present in the larva differentiated into several adult cell types (Amano and Hori, 1996, 2001) and rearranged to develop the aquiferous system. Settled larvae completed metamorphosis in about 1 week and a prominent exhalant tube was formed. From then on, juvenile sponges were able to filter water and feed. Nevertheless, settlers of H. communis and I. oros did not show conspicuous inhalant/exhalant orifices even through light microscope. At this state, most juveniles of the two species died before forming the skeletal fibers.

(A) Larva of D. avara. (B) Larva of C. crambe. (C) D. avara juvenile after 2 days from settlement. Arrowheads, choanocyte chambers; gm, growth marginal layer-the white points correspond to cells. (D) C. crambe juveniles after 2 days from settlement. gm, Growth marginal layer.
Most juveniles of D. avara and C. crambe survived, allowing morphological changes to be monitored. Two days after settlement a monolayered pseudoepithelium (growth marginal layer) was visible through the stereomicroscope (Figure 1C, D). In the case of D. avara, 1-week-old juveniles already showed from 20 to 80 choanocyte chambers and a long exhalant tube (Figure 2A). These early juveniles are called rhagons. Fifteen days after settlement, some structural elements were formed: rudiments of spongin fibers in D. avara (Figure 2B, C) and spicules and collagen in C. crambe. Choanocyte chambers increased in number and were densely packed. In that stage, sponge juveniles grew mainly in height. One month after settlement, the choanocyte chambers are connected to a complex network of inhalant and exhalant canals. The completely organized exhalant canals flow into a cloacal exhalant tube that ends in an osculum (Figure 2C). In some of these juveniles with more than 200 choanocyte chambers, more than one exhalant tube was present (Figure 2C, D).

(A) D. avara, 1-week-old juveniles. Black arrowheads, few choanocyte chambers; white arrowhead, exhalant tube. (B) D. avara, 15-day-old juvenile. Black arrowhead, skeletal fiber; white arrowhead, exhalant tube. (C) D. avara, 1-month-old juveniles. f, Skeletal fibers; black arrowhead, canals; white arrowheads, exhalant tubes. (D) C. crambe, 15-day-old juveniles. Arrowheads, two exhalant tubes.
Settlement Success, Survival and Growth Rates of Juveniles
-
1.
Species Comparison In the first experiment, differences in settlement success and survival were found among H. communis, I. oros, and D. avara. The three species presented a high settlement success: 66.6% in H. communis, 99.5% in D. avara, and 94.5% in I. oros (Figure 3). The different percentage of settlers for the three species under the same laboratory conditions suggests that success in settlement may depend on species-specific environmental requirements and/or particular larval characteristics. Although general acceptable conditions have been found in our cultures since the settlement rates obtained appeared to be higher than those at sea (Uriz et al., 1998), optimal culture conditions should be species specific. The high settlement success obtained in the laboratory may be due to the extremely favorable conditions that larvae encountered there for settlement (i.e., still water and no substrate competition). At sea, larvae do not settle immediately but are transported by currents and must swim until they find appropriate conditions (Kaye and Reiswig, 1991; Uriz et al., 1998). During the swimming period, larvae are spending their reserves, which may become depleted before settling. Depletion of reserves in early life stages may strongly affect the success in the next stages. It has been reported for other invertebrates that adults perform worse when they originate from settlers with lower energy contents (Marshall and Keough, 2003). It also has been shown for the demosponge Sigmadocia caerulea that juveniles originating from larvae that settled fast survived better, grew faster, and were more regular in shape than those originating from long living larvae (Maldonado and Young, 1999). Survival of juveniles with time was significantly different among the three species (P < 0.05, Comparing Survival in Multiple Groups) (Figure 4). D. avara was the species with the highest percentage of survivors (P < 0.05 in both comparisons) throughout the experiment. Survival in this species ranged from 100% (at day 5) to ca. 80% (at day 80), reaching 62% at the end of the experiment (after 3 months). There were no significant differences (P = 0.8943 Gehan’s Wilcoxon Test) between H. communis and I. oros. Only 15% of H. communis juveniles survived at the end of the experiment, and no survivors of I. oros occurred after day 50, despite settlement success was high (see earlier). In the three species, a maximum increase in area took place at the beginning of the culture: at week 1 in D. avara and at week 2 in H. communis and I. oros. This maximum increase in area did not represent true growth but corresponded to biomass redistribution due to the reorganization processes involved in metamorphosis. After metamorphosis, juveniles displayed sequential phases of growth: first they increased in area by extending the marginal growth layer and then they formed structural elements, grew in thickness, and decreased in area. Some juveniles that could not grow in thickness due to a physical barrier (sandwich culture) grew exclusively in area (Figure 5), achieving the largest size. Thus, as stated in the methods section, we were aware that the measured growth underestimated real growth but was considered accurate enough for comparative purposes. Taken into account settlement success and survival rates measured during the experiment, we selected D. avara among the three species studied, as a model for the next experiment in which different environmental conditions were assayed. C. crambe was selected as a second species because juveniles have been reported to show high survival rates in a previous study (Uriz et al., 1998).
-
2.
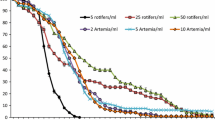
Comparison of Culture Conditions In a second experiment, we tested the effects of water flow and food type (living versus dried algae) on the survival and growth of juveniles of D. avara and C. crambe. Survival of both species was high in all treatments (between 80% and 100%) except for D. avara under no-flow conditions, which showed a significantly lower survival (32%) (P < 0.001 Comparing Survival in Multiple Groups; Figures 6 and 7). As for growth rates, no significant differences (P = 0.504) were observed between settlers of D. avara cultured under flow and no-flow, but the interaction term (time × treatment) was significant (P < 0.001), indicating that the trends changed with time. Conversely, C. crambe showed a significantly higher growth rate under flow conditions (P < 0.001) than without flow, and also the trend varied with time (significant interaction term P < 0.001). Comparing both species, differences were significant under both conditions: C. crambe grew more (P < 0.001) than D. avara under flow, in particular during the first months, and the interaction term (time × treatment) was also significant (P < 0.001). In contrast, D. avara grew more (P < 0.001) than C. crambe under no flow and the trends run in parallel with time (no significant interaction term, P = 0.202; Figure 8). When the kind of food was tested, no differences between C. crambe fed with either dried or living algae were observed (P = 0.138). In contrast, a significantly higher growth rate was recorded when D. avara was fed with dried algae (P < 0.001) than with living algae. In a comparison of both species, C. crambe showed higher growth rates (P < 0.01) than D. avara when the sponges were fed with living algae, and the interaction term was significant (P < 0.001). In contrast, no differences were found when the food consisted of dried algae (P = 0.334; Figure 9). However, at the last time point of monitoring (after 2 months), differences in growth rates were significantly higher for both species fed with dried algae (two-way ANOVA, P < 0.01). The results found altogether reflect the particular characteristics of each species. C. crambe has a conservative life strategy with slow growth, lower clearance rates (Turon et al., 1997), and lower volume of choanocyte chambers (Uriz et al., 1995; Galera et al., 2000) than D. avara, which makes the former species less able to capture food under adverse conditions (i.e., lack of flow). Conversely, D. avara has an opportunistic life strategy with higher growth and clearance rates than C. crambe and also has a strong filtering activity (Ribes et al., 1999); thus it is able to capture food from the water even under no-flow conditions (lower particles availability). These differential capabilities may explain why C. crambe grew more under flow conditions that can enhance the low sponge capacity for pumping water and facilitate the availability of food to the sponge. On the other hand, D. avara has been reported to be a dynamic species with a fast metabolism (Turon et al., 1997) and thus it seems to need relatively high amounts of food for survival. Dried algae were more effective than living algae to feed sponges likely because dried algae consisted of broken cells with a smaller particle size than intact algal cells (from 4 to 8 μm, P. tricornutum). In previous studies, the highest clearance rates were obtained with 1 μm particle size for D. avara and C. crambe, while larger particles (4 μm) were retained with less efficiency (Turon et al., 1997). However, although the size of the algae used can enter the sponge ostia in both species (ostia diameter is 11.4 ± 1.2 μm in C. crambe and 30.8 ± 2.2 μm in D. avara, Galera et al. 2000), they may not enter through the smaller orifice (prosopyle) connecting inhalant canals and choanocyte chambers (de Vos et al., 1991), which may complicate algae capture. If we consider the last observation time (see Figure 9), we can conclude that the two types of food determined the final growth rate for both species. The dry algae appeared to be significantly more effective as a food source than the living algae, probably due to a higher retention rate of the sponge for the dry algae.
-
3.
Juvenile Survival and Growth in the Field In a third experiment, juveniles of D. avara and C. crambe settled in the laboratory were transferred to the sea in order to monitor survival and growth under natural conditions. Juveniles of C. crambe had a significantly higher survival than those of D. avara (P < 0.001, Gehan’s Wilcoxon test). During the first 20 days of the monitoring, both species experienced a high mortality (about 60%) and from then on, differences between species became evident: all the D. avara juveniles died after day 60 whereas 24% of C. crambe juveniles survived at that day. Moreover, 20% of C. crambe juveniles survived until day 136 (Figure 10). The higher mortality of D. avara juveniles in the field may be due to a higher palatability and a more fragile consistency of this species with respect to C. crambe (Uriz et al., 1996a). Juveniles cultured in the field survived significantly less than juveniles cultured in the laboratory (previous experiment) for the two species (Figure 11) (P < 0.001 Comparing Survival in Multiple Groups). In natural conditions, a high mortality is frequently reported for juvenile stages of sponges because of predation, competition for the substrate with other benthic organisms (Uriz et al., 1996a), and physical perturbations. All these drawbacks can be avoided by culturing larvae in the laboratory, and, thus, culture of juveniles under controlled conditions are recommended for avoiding biomass losses at these early stages. Growth rates at sea were not significantly different between species (Figure 12) (P = 0.101, and interaction term P = 0.235). D. avara, growth rates at sea were similar (P = 0.294) to those in the laboratory fed with dried algae. However, under less favorable conditions, juveniles grown in the laboratory grew significantly less that those at sea (P < 0.001). In contrast, C. crambe juveniles grew significantly more (P < 0.01) at sea than in the laboratory regardless of the experimental conditions (Figure 12). In the field site where juveniles were placed, C. crambe is much more abundant than D. avara, which may indicate better environmental conditions for the former species to grow. Further, settlers of D. avara appeared to be more fragile than those of C. crambe and thus unable to resist physical perturbations. Predation can also contribute to the differential mortality of both species since D. avara has been reported to be grazed by sea urchins (Uriz et al., 1996a), whereas no predators are known for C. crambe. Further, growth did not differ between juveniles cultured in the laboratory and those transplanted to the sea. Conversely, survival was enormously higher in the laboratory. Comparing growth rates of juveniles of both species in the laboratory under the best conditions (flow and dried algae as a food) with those of adult individuals monitored during ca. 4 years in the field (Turon et al., 1998, 2000): D. avara juveniles presented higher growth rates in our cultures (average 0.44 ± 0.22 per month) than adults did in the field (average 0.03 ± 0.01 per month, Turon et al., 1998). Similarly, considerable differences in growth rates were observed between juveniles (0.46 ± 0.2 per month) and adults of C. crambe (average 0.1 ± 0.02 per month, Turon et al., 1998). In contrast, growth variability was higher in juveniles (this study) than in adult sponges for both species. The variation coefficient for larval growth obtained from this study was 66.6% and 48.9% for D. avara and C. crambe juveniles respectively, while it was 33.3% and 20% for D. avara and C. crambe adults, respectively (Turon et al., 2000). These data reflect the inherent growth variability in sponges since, even eliminating the age factor, the level of variation is high. However, the comparison between juveniles and adults must be taken with care because the phases of growth in thickness in our juveniles were associated with a decrease in area (see earlier), which inevitably was incorporated in our growth data, contributing to the variability recorded. Altogether, these results point to sponge larvae as a promising source for the sponge/metabolite supply, because the culture is performed during the period of maximum sponge growth and thus the culture yield can be optimized. The study has also provided interesting information about biological aspects on the sponge life cycle, helping to fill the gap in the literature about juvenile stages, which are difficult to follow in the field because of their small size.

Settlement success of larvae of D. avara, H. communis. and I. oros under the same laboratory conditions (filtered seawater at 20°C under static conditions).

Survival of D. avara, I. oros, and H. communis juveniles with time under the same laboratory conditions (filtered seawater at 20°C under static conditions).

(A) Lateral view of a juvenile D. avara showing its thickness. (B) D. avara juvenile allowed growing only in two dimensions (sandwich culture).

Survival of D. avara and C. crambe juveniles with time under flow versus no-flow conditions; juveniles were fed with living algae. D. avara survival under no-flow conditions showed the significantly lowest survival (P < 0.001).

Survival of D. avara and C. crambe juveniles with time fed with living algae versus dried algae; juveniles were under flow conditions. No significant differences were observed.

Time course of the growth rate of D. avara (N = 30) and C. crambe (N = 50) juveniles, under flow versus no-flow conditions; juveniles were fed with living algae. Vertical lines are standard errors.

Time course of the growth rate of D. avara (N = 30) and C. crambe (N = 50) juveniles, fed with living versus dried algae; juveniles were under flow conditions. Vertical lines are standard errors.

Survival of D. avara and C. crambe juveniles with time at sea.

Survival comparison of D. avara and C. crambe juveniles with time under the best laboratory conditions (with flow and fed with dried algae), and in the field.

Comparison of growth rates of D. avara (N = 30) and C. crambe (N = 50) juveniles under the best laboratory conditions (with flow and fed with dried algae), and in the field (D. avara N = 164, and C. crambe N = 175). Vertical lines are standard errors.
References
Amano S, Hori I (1996) Transdifferentiation of larval flagellated cells to choanocytes in the metamorphosis of the demosponge Haliclona permollis. Biol Bull 190, 161–172
Amano S, Hori I (2001) Metamorphosis of coeloblastula performed by multipotential larval flagellated cells in the calcareous sponge Leucoselenia laxa. Biol Bull 200, 20–32
Balconi G, Erba E, Bonfontani M, Filipperschi S, Garcia de Quesada T, Jimeno JM, D’Incalci M (1995) Antiproliferative activity and mode of action of novel compounds of marine origin. Eur J Cancer 31 (Suppl 6), S26
Blunt JW, Coop BR, Munro MH, Northcote PT, Prinsep MR (2005) Marine natural products. Nat Prod Rep 22, 15–61
Boury-Esnault N, Rützler K (1997) Thesaurus of Sponge Morphology (Washington, DC: Smithsonian Contributions to Zoology), p 596
Cimino G, De Stefano S, Minale L, Fattorusso E (1972) Ircinin-1 and -2, linear sesterpenes from the marine sponge Ircinia oros. Tetrahedron 28, 333–341
De Caralt S, Agell G, Uriz MJ (2003) Long-term culture of sponges explants: conditions enhancing survival and growth, and assessment of bioactivity. Biomol Eng 20, 339–347
De Rosa S, De Caro S, Iodice C, Tommonaro G, Stefanov K, Popov S (2003) Development in primary cell cultura of demosponges. J Biotechnol 100, 119–125
De Vos L, Rützler K, Boury-Esnault N, Donadey C, Vacelet J (1991) Atlas of Sponge Morphology (Washington and London: Smithsonian Institution Press)
Duckworth A, Battershill CN (2003) Sponge aquaculture for the production of biologically active metabolites: the influence of farming protocols and environment. Aquaculture 221, 311–329
Fusetani N (2000) Introduction In: Drugs from the Sea, Fusetani N, ed. (Basel: Karger) pp 1–5
Galera J, Turon X, Uriz MJ, Becerro M (2000) Microstructure variation in sponges sharing growth form: the encrusting demosponges Dysidea avara and Crambe crambe. Acta Zool 81, 93–107
Garrabou J, Zabala M (2001) Growth dynamics in four Mediterranean demosponges. Estuar Coast Shelf S 52, 293–303
Hoffmann F, Rapp HT, Zöller T, Reitner J (2003) Growth and regeneration in cultivated fragments of the boreal deep water sponge Geodia berretti Bowebank, 1858 (Geodiidae, Tetractinellida, Demospongiae). J Biotechnol 100, 109–118
Jares-Erijman EA, Sakai R, Rinehart KL (1991) Crambiscidins: new antiviral and cytotoxic compounds from the sponge Crambe crambe. J Org Chem 56, 5712–5715
Kaye HR, Reiswig HM (1991) Sexual reproduction in four Caribbean commercial sponges III Larval behaviour, settlement and metamorphosis. Invertebr Reprod Dev 19, 25–35
Loya S, Hizi A (1990) The inhibition of human immunodeficiency virus type 1 reverse transcriptase by avarol and avarone derivatives. FEBS Lett 269, 131–134
Maldonado M, Young CM (1999) Effects of the duration of larval life on postlarval stages of the demosponge Sigmadocia caerulea. J Exp Mar Biol Ecol 232, 9–21
Manly FJ (1991) Randomization and Montecarlo Methods in Biology (London: Chapman and Hall)
Mariani S, Uriz MJ, Turon X (2000) Larval bloom of the oviparous sponge Cliona viridis: coupling of larval abundance and adult distribution. Mar Biol 137, 783–790
Mariani S, Uriz MJ, Turon X (2005) The dynamics of sponge larvae assemblages from northwestern Mediterranean nearshore bottoms. J Plankton Res 3, 249–262
Marshall DJ, Keough MJ (2003) Variation in the dispersal potential of non-feeding invertebrate larvae: the desperate larva hypothesis and larval size. Mar Ecol Prog Ser 255, 145–153
Mendola D (2003) Aquaculture of three phyla of marine invertebrates to yield bioactive metabolites: process developments and economics. Biomol Eng 20, 37–82
Moore HF (1910) A practical method of sponge culture. Bull US Bur Fish 28, 545–585
Müller WEG, Maidhof A, Zahn RK, Schröder HC, Gasic MJ, Heidemann D, Bernd A, Kurelec B, Eich E, Sibert G (1985) Potent antileukemic activity of the novel cytostatic agent avarone and its analogues in vitro and in vivo. Cancer Res 45, 4822–4827
Müller WEG, Wiens M, Batel R, Steffen R, Schröder HC, Borejevic R, Custodio MR (1999) Establishment of a primary cell culture from a sponge: primmorphs from Suberites domuncula. Mar Ecol Prog Ser 178, 205–219
Nickel M, Brümmer F (2003) In vitro sponge fragment culture of Chondrosia reniformis (Nardo, 1847). J Biotechnol 100, 147–159
Osinga R, de Beukelaer PB, Meijer EM, Tramper J, Wijffels RH (1999) Growth of the sponge Pseudosuberites (aff) andrewsi in a closed system. J Biotechnol 70, 155–161
Osinga R, Kleijn R, Groenendijk E, Niesink P, Tramper J, Wijffels R (2001) Development of in vivo sponges cultures: particle feeding by the tropical sponge Pseudosuberites aff andrewsi. Mar Biotechnol 6, 544–554
Osinga R, Belarbi EH, Molina Grima E, Tramper J, Wijffels RH (2003) Progress towards a controlled culture of the marine sponge Pseudosuberites andrewsi in a bioreactor. J Biotechnol 100, 141–146
Pomponi SA, Willoughby R (1994) “Sponge cell culture for the production of bioactive metabolites”. In: Sponges in Time and Space, van Soest R, van Kempen TMG, Braekman JC, eds. (Rotterdam: A Balkema) pp 395–400
Potvin C, Lechowicz MJ, Tardif S (1990) The statistical analysis of ecophysiological response curves obtained from experiments involving repeated measures. Ecology 71, 1389–1400
Procksh P, Edrada-Ebel RA, Ebel R (2003) Drugs from the sea-opportunities and obstacles. Mar Drugs 1, 5–17
Pronzato R, Bavestrello G, Cerrano C, Magnino G, Manconi R, Pantelis J, Sarà A, Sidri M (1999) Sponge farming in the Mediterranean Sea: new perspectives. Mem Qld Mus 44, 485–491
Ribes M, Coma R, Gili JM (1999) Natural diet and grazing rate of the temperate sponge Dysidea avara (Demospongiae, Dendroceratida) through an annual cycle. Mar Ecol Prog Ser 176, 179–190
Rifai S, Fassoune A, Kijjoa A, Van Soest R (2004) Antimicrobial activity of Untenospongin B, a metabolite from the marine sponge Hippospongia communis collected from the Atlantic Coast of Morocco. Mar Drugs 2, 147–153
Rinkevich B (2005) Marine invertebrate cultures: new millennium trends. Mar Biotechnol 7, 429–439
Rinkevich B, Blisko R, Ilan M (1998) Further steps in the initiation of cell cultures from embryos and adult sponge colonies. In Vitro Cell Dev Biol 34, 753–756
Sarin PS, Sun D, Thornton A, Müller WEG (1987) Inhibition of replication of etiologic agent of AIDS by avarol and avarone. J Natl Cancer Inst 78, 663–666
Simpson TL (1984) The Cell Biology of Sponges (New York: Springer-Verlag)
Sipkema D, Snijders APL, Schröen CGPH, Osinga R, Wijjffels R (2003) The life and death of sponge cells. Biotechnol Bioeng 85, 239–247
Sipkema D, Franssen MCR, Osinga R, Tramper J, Wijffels RH (2005a) Marine sponges as pharmacy. Mar Biotechnol 7, 142–162
Sipkema D, Osinga R, Schatton W, Mendola D, Tramper J, Wijjffels R (2005b) Large-scale production of pharmaceuticals by marine sponges: sea, cell or synthesis? Biotechnol Bioeng 90, 202–222
Sipkema D, Yosef NAM, Adamczewski M, Osinga R, Tramper J, Wijffels RH (2006) Hypothesized kinetic models for describing growth of globular and encrusting demosponges. Mar Biotechnol 8, 40–51
Turon X, Galera J, Uriz MJ (1997) Clearance rates and aquiferous systems in two sponges with contrasting life-history strategies. J Exp Zoo 278, 22–36
Turon X, Tarjuelo I, Uriz MJ (1998) Growth dynamics and mortality of the encrusting sponge Crambe crambe (Poecilosclerida) in contrasting habitats: correlation with population structure and investment in defence. Funct Ecol 4, 631–639
Turon X, Galera J, de Caralt S, Tarjuelo I, Uriz MJ (2000) Pautas de crecimiento en esponjas incrustantes del Mediterranio: seguimiento a largo plazo de Crambe crambe (Poecilosclerida) y Dysidea avara (Dendroceratida). XI Simposium Ibérico: Estudios del Bentos Marino Málaga, Spain
Uriz MJ, Turon X, Becerro MA, Galera J, Lozano J (1995) Patterns of resource allocation to somatic, defensive, and reproductive functions in the Mediterranean encrusting sponge Crambe crambe (Demospongiae, Poecilosclerida). Mar Ecol Prog Ser 124, 159–170
Uriz MJ, Becerro MA, Tur JM, Turon X (1996a) Location of toxicity within the Mediterranean sponge Crambe crambe (Demospongiae: Poecilosclerida). Mar Biol 124, 583–590
Uriz MJ, Turon X, Galera J, Tur JM (1996b) New light on the cell location of avarol within the sponge Dysidea avara (Dendroceratida). Cell Tissue Res 285, 519–527
Uriz MJ, Maldonado M, Turon X, Martí R (1998) How do reproductive output, larval behaviour, and recruitment contribute to adult spatial patterns in Mediterranean encrusting sponges? Mar Ecol Prog Ser 167, 137–148
Von Ende CN (1993) Repeated measures analysis: growth and other time-dependent measures. In: Design and Analysis of Ecological Experiments, Scheneider SM, Gurevitch J, eds. (Oxford: Oxford University Press) pp 113–137
Zhang X, Cao X, Zhang W, Yu X, Jin M (2003) Primmorphs from archaeocytes-dominant cell population of the sponge Hymeniacidon perleve: improved cell proliferation and spiculogenesis. Biotechnol Bioeng 84, 583–590
Acknowledgments
This study was partially funded by the project from the European Union INTERGEN CICYT: CTM2004-05265-C02-02/MAR. We thank M. Ribes, E. Cebrian, and A. Blanquer for their help in larval supply and field monitoring, and X. Turon for facilitating the program to perform the permutation tests.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
de Caralt, S., Otjens, H., Uriz, M.J. et al. Cultivation of Sponge Larvae: Settlement, Survival, and Growth of Juveniles. Mar Biotechnol 9, 592–605 (2007). https://doi.org/10.1007/s10126-007-9013-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10126-007-9013-5