
Abstract
The limnological record of human impact on catchment land cover and on lake sedimentation during the historical period has been established for Pinto Lake in Central Coastal California. In addition, the sedimentary record of the ‘pre-impact’ condition preserves evidence of a climatic control on the nature of lake sedimentation. Chronological marker horizons have been determined using pollen data in combination with the documented land-use history and introductions of exotic species. Further chronological data have been determined using 14C and 137Cs. The impact of Mexican and Euro-American immigrants and their ‘imported’ land-use practices is clearly reflected in an order of magnitude increase in the rate of lake sedimentation to c. 9 kg m−2 year−1 (c. 2 cm year−1) between 1770 and 1850. Here, the occurrence of exotic plant species indicates disturbance as early as c. 1769–1797, whilst redwood deforestation between 1844 and 1860 represents the most significant human impact. Changes in the nature of sedimentation prior to this reveal a high degree of sensitivity to changes in precipitation where subtle decreases in lake level and the supply of runoff-derived mineral matter have resulted in two periods of organic lake sedimentation c. 650–900 and 1275–1750. Set against this background condition of high sensitivity, the dramatic impacts of Euro-American settlement are unsurprising.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Although studies of climate impact on terrestrial sediment flux during the Holocene provide a long-term context for sustainable management of lake and lagoon ecosystems, the period of most acute human impact on catchments and river systems across the globe is the last 200 years or so (Mannion 1995). Furthermore, as noted by Battarbee (2000), whilst late Holocene climate signals are either obscured or made ambiguous by the overprint of human activity, the problem is more chronologically constrained in North America where most catchments remained relatively undisturbed until European immigration. In much of North America, human-accelerated runoff did not begin until the late 18th or the 19th centuries (Knox 2000). Indeed, the record from Frain’s Lake, Michigan (Davis 1976) shows a dramatic increase in sediment influx following deforestation, with eventual stabilisation at a higher level than that of pre-disturbance times. The research documented here focuses in particular on the impact of human activity through clearance and agriculture on sediment delivery to lowland catchments. In the case of Pinto Lake, located in lowland Central Coastal California on the northeastern shore of Monterey Bay (Fig. 1), we have sought to resolve the impact of Mexican and Euro-American settlement and recent agricultural and urban development on sedimentation, with a view to quantifying sediment flux to the coastal zone and determining the lifetime of surface water resources.

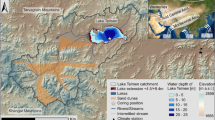
Location of Pinto Lake within the Watsonville lakes region, Santa Cruz (a), and the morphology of Pinto Lake and its catchment (b)
Thematically, our research takes an ecohydrology perspective (cf. Zalewski 2002) in examining lake system response. In seeking to identify the underlying patterns of sedimentation controlled by climatic variability, thus providing a context for evaluating the significance of human impact and the inherent resilience of Pinto Lake to such disturbance, we reflect on the role of biota and process interaction in moderating various elements of catchment and lake robustness. This understanding provides a framework for future management strategies that enhance the natural capacity of the catchment and wetland continuum to absorb and/or process sediments, nutrients and pollutants (Wolanski et al. 2004)
In being attuned to the key research questions of the PAGES-HITE programme, this study focuses on establishing the nature and response of recent lake sedimentation in lowland Central Coastal California and seeks to investigate: (a) the extent of human impact on the lake ecosystem; (b) how these impacts have interacted with natural climate-driven variability in lake level, water quality and sediment flux; and (c) what evidence the sedimentary record provides in the context of resilience, rates of recovery, irreversibility and future sensitivity. Thus, it provides a long-term perspective and context for catchment management where monitoring data or instrumental records are largely absent. Threats to lake systems such as Pinto Lake include climate change, nutrient enrichment from domestic and agricultural sources, reduced water quality from extraction of freshwater, pollution from toxic metals and organic compounds derived from the surrounding catchment and from the atmosphere, and accelerated infill from enhanced catchment soil erosion and changes in biological productivity.
Thus, our research also specifically addresses key research questions of PAGES-LIMPACS in that we aim:
-
To establish the natural, pre-disturbance condition (i.e. prior to Mexican immigration and Euro-American settlement)
-
To identify the impact of human activity and separate this from natural variability
-
To identify and quantify (in terms of sediment flux) the relative roles of different causes of lake change (e.g. catchment land-use, water quality and lake productivity)
-
To set goals for future land-use and catchment management based on knowledge of past and present lake status.
Pinto Lake—physiography and environmental change
Location and morphology
Pinto Lake is located in the Central Coast region of California some 5 km to the north-east of Watsonville in the Pajaro Valley (Fig. 1a). Several creeks feed the lake, which then drains through an underground outlet pipe into Corralitos Creek, the Pajaro River, and eventually into Monterey Bay at the south end of Sunset Beach State Park. Pinto Lake is located in a small valley between two sides of a terrace deposit (Fig. 1b), and is considered to have formed as a consequence of sag pond development along the Zayante Fault (S. Lydon, personal communication; Dupre 1975). The low terraces that border Pinto Lake are characterised by medium to heavy textured soils near the surface and more compact below (Grunwald et al. 1974). These terraces are highly vulnerable to erosion, especially gully erosion.
The lake is roughly oval in shape with four tributary arms to the north formed from earlier incision of the terrace landscape. As a result of this incision coupled with tectonic uplift, the lake margins are generally quite steep on all but the southern shore. The main lake basin reflects these steep sides, and is 6–8 m deep over much of its 0.35 km2 area. The exception to this is the tributary arms which are less than 2 m and commonly little more than a metre or so deep. Pinto Lake has a catchment area of about 5.5 km2, of which 2.2 km2 is relatively steep slope, giving a lake to catchment ratio of c. 1:15. The land-use immediately around the lake includes both Regional and County parks, with the catchment being characterised by small urban developments, grassland, woodland and agriculture—including the growing of row crops and pasture.
Central Coast climate
The climate is characterised by cool, wet winters and warm, dry summers. During winter, the North Pacific Subtropical High weakens and is displaced equatorwards, allowing frontal systems from the Aleutian Low to sweep further south (Johnson 1977). Strengthening of the Pacific High during spring and summer re-establishes its dominance. At the same time, a low-pressure system forms over the interior, generating an almost continuous flow of moist onshore air during the warm season, which is cooled over the cold upwelling ocean waters and leads to the persistence of fog in summer. Precipitation is both seasonally and interannually variable (Redmond and Koch 1991). A significant component of the interannual variability can be attributed to the El Niño/Southern Oscillation (ENSO), with both extremes of the ENSO phenomenon (El Niño and La Niña) showing a teleconnective effect in western North America (Kiladis and Diaz 1989; Diaz and Markgraf 1992). The consensus regarding the link between ENSO and California precipitation supports increased precipitation in the south and a reduced or reversed relationship with precipitation in the north (Fu et al. 1986; Schonher and Nicholson 1989). McCabe and Dettinger (1999) also found a significant correlation between average and (the following) winter precipitation for the Central Coast with Type 1 El Niño years for the period 1961–1990. Similarly, Cayan et al. (1999) have shown the strongest links between SOI- (El Niño) and climate in Central Coastal California, with an increase in the number of abnormally high precipitation/streamflow days.
A plot of moving averages in precipitation in Coastal California between Los Angeles and Monterey reveals a cyclicity with an amplitude of 100–150 mm and a periodicity of 25–35 years (Cooke and Reeves 1976). Indeed, a similar 20–30 years periodicity is recognised in tree rings from the Transverse Ranges of the Santa Barbara Basin (Michaelson et al. 1987). A number of predominantly wet years dominate the periods 1883–1890, 1904–1916 and 1936–1944, with the interval from 1861 to 1891 also being on record as a very stormy period for Arizona and Utah (Ely et al. 1993). In addition, Engstrom (1996) writes of a widespread flooding and storm event in California in 1861/1862. In San Francisco, for example, 619 mm of precipitation (c. five times normal) fell in January. The intervening periods contain more than a usual number of dry years, with dry periods in the years 1862–1864, 1869–1871, 1881–1883, 1897–1900, 1928–1934, 1946–1950 and 1958–1961 (Cooke and Reeves 1976). Of these dry periods, the two most notable date from the close of the late 1890s and the late 1920s to early 1930s.
Human impact on the Central Coast region
The landscape of Coastal California was largely unaffected by early nomadic aboriginals who lived by fishing and exploiting seasonally available resources (Patch and Jones 1984; Gordon 1996). In Monterey Bay, whilst the Ohlone Indians maintained a littoral existence, hunting and gathering was important for other tribes. These tribes created fields to support grazing and to grow crops (Frenkel 1970) and depleted the woodlands for firewood. In the Santa Cruz region, the raised marine terrace landscape was maintained as grassland (Gordon 1996).
In the mid-18th century Spain asserted control on Alta California through the establishment of Presidios and Missions, with 1769 marking California’s inclusion in the Spanish Empire. Immigration replaced native land-use with permanent settlements based on a pastoral lifestyle. Immigrants created grazing land by burning, and subsequent cattle ranching led to considerable trampling and increased demand for fodder and pasture. Cereals, legumes and orchard plants were also established, although soil tilling and irrigation was generally a localised activity. Land degradation was exacerbated following Mexico’s independence from Spain in 1822, leading to the introduction of more liberal policies that allowed greater access to land. According to Jarrell (1982), more than one half of the land in Santa Cruz County was divided into ranchos ranging in size from 45 to 30,000 acres. This initiated agriculture on a large scale, which began locally in the Pajaro valley in 1823 (Lewis 1994). Cattle ranching thrived, burning was still employed to improve pasturage, and small-scale cultivation continued (Allen 1935).
Following the Mexican–American War, California became a US territory. With the discovery of gold in 1848, mining not only changed the social structure of the State but also directly and indirectly altered the landscape. When first settled by the Americans, the Santa Cruz Mountains at the northern margin of the Pajaro Valley were heavily wooded by redwoods and evergreen forests. Commercial logging began in the early 1850s in the northern part of Monterey Bay, and by 1860 Santa Cruz County became the second largest redwood producer in the State (Jarrell 1982). The logging industry increased through the 1880s, but had begun to decline by 1901 due to exhaustion of redwood stands. Tanbark oak was also heavily forested for tanning hides, making Santa Cruz County one of the largest tanning centres in 1870, but this too was in decline by 1918 (Gordon 1996). Douglas fir continued to be logged commercially into the mid-20th century, creating further areas of bare soil and logging debris.
By the early 1860s, following the decline in gold mining after 1852, the Watsonville area developed as an agricultural centre. However, the effects of intense grazing between 1850 and 1863, coupled with the 1862 flood and the 1863–1864 drought (Engstrom 1996), brought with it the search for alternative prospects for land in the form of cereal cultivation (Lantis 1970). This trend of agricultural expansion led to further destruction of woodland, with American settlers clearing trees for firewood and to make the land suitable for arable purposes (Gordon 1996). From the mid-19th century, large-scale farming stretched for miles along the Central Coast valleys, characterised by extensive ploughing, channelised drainage and irrigation. By the end of the century, a diversified agricultural economy had, therefore, replaced grazing as the dominant economy.
In terms of geomorphic and sedimentary evidence of catchment destabilisation linked to human activity, the first surveyors’ descriptions in 1850, the 1870s and the 1880s revealed that channels with defined banks in valley floors had become well-established (Cooke and Reeves 1976). Within Coastal California, arroyo lengthening during the period of 1850 into the 1920s can be linked to the changes in the flora and soil structure of grasslands and woodlands which were extensively transformed as a result of introduced land-use practices. Even in areas distant from cultivated lands, livestock ranching played an important role in determining catchment instability. Mudie and Byrne (1980) determined that pre-Euro-American settlement sedimentation rates in coastal marshes from southern and central California were approximately 1 and 0.5 mm year−1, respectively. These are significantly lower than those rates observed during the 20th century for the same sites, i.e. 5 and 1 mm year−1, respectively. Rapid coastal sedimentation is further illustrated in Elkhorn Slough, where Patch and Jones (1984) record the presence of a Eucalyptus grain at a depth of 340 cm—indicating deposition over a time period no greater than 120 years at an average rate of c. 25–30 mm year−1.
Methods
Fieldwork in 1998 recovered a 2.3 m core (PL1998) from the bed of Pinto Lake. From consideration of previous estimates of coastal sedimentation in the region of between 1 and 5 mm year−1 (e.g. Mudie and Byrne 1980; Eittreim et al. 2002), this sediment had the potential to record changes in lake sedimentation over the period of c. 400–2500 years bp. However, to ensure capture of a pre-impact condition, efforts were made to recover a deeper lake sediment record in 2001. In this case, a 6.2 m core (PL2001) was collected using a Stitz piston corer deployed from a raft.
The approach taken in this study adopts the combined palaeoecology and radiometric dating methodology used with considerable success in previous investigations of this kind in the United States. Robbins et al. (1978) combined 210Pb, 137Cs and pollen chronologies from the sedimentary record of lakes Ontario and Erie to obtain sediment accretion rates since the onset of intensive regional forest clearance in approximately 1850 ad. Indeed, the increase in Ambrosia sp. (ragweed) is commonly linked to the initial clearing of land and intensive agriculture by European settlers in the US. Brugam (1978) used the first occurrence of Ambrosia and Rumex with increases in Poaceae and decreasing Tsuga (western hemlock) to mark the arrival of European farming in Connecticut, recording two changes in sedimentation rate in Linsley Pond around 1700 ad. With the addition of 14C dating and tephrochronology, a similar approach has been used to resolve the impact of the Spanish Conquest and Colonial periods on lake systems in Mexico (e.g. Davies et al. 2004). In the context of coastal wetlands, Brush (1989, 1994) was able to identify the impacts of intensive land clearance in the tidal tributaries of Chesapeake Bay when more than 20% of the total land area was deforested and put under cultivation, resulting in a twofold increase in sedimentation rate. In the Potomac estuary, rates of sedimentation from unsupported 210Pb dating compare well with rates determined from the increase in Ambrosia pollen representing the ‘agricultural revival’ in 1840 ad (Brush et al. 1982). Similarly, Cole and Liu (1994) recognise significant human impact on Santa Rosa Island since about the 1840s, with a peak in charcoal followed by increasing fungal spores and pollen from Ambrosia sp., the goosefoot family, storksbill and cypress. Here, estuary sedimentation increased from 0.07 to 1.34 cm year−1 as a consequence.
Accordingly, we have sought to establish a timeframe for the period of deposition based on palaeoecological phenomena preserved in the sedimentary record which can be linked to the history of human activity in the region. Particular emphasis is placed on vegetation change related to human impact and/or lake margin ecology brought about by changes in lake level. Further chronological data have been acquired from 14C and 137Cs dating, with additional reference to previous research in which a well-resolved chronology of DDT pollution was determined for PL1998 (Mayers 2001).
Pollen
Pollen slides were prepared by carrying out a series of acid washes interspersed with washes of double distilled water. Carbonates were removed using 10% HCl, then alkali soluble humic compounds were removed by washing in 10% NaOH. The samples were filtered through a 100 μm sieve mesh in order to remove any coarse material. Clay was removed with 10% sodium pyrophosphate, and the samples then left in HF acid and decanted into a calcium hydroxide solution. Samples were washed with glacial acetic acid, followed by a wash with acetic anhydride and sulphuric acid in order to remove any soluble cellulose. Safranine was added to stain the pollen grains and samples were mounted onto microscope slides with glycerol.
Loss-on-ignition
Loss-on-ignition (LOI) was used as a proxy for organic content. Approximately 5 g of material was dried at 105°C to determine air-dry weight. This was then ignited at 450°C for 4 h. Once cool, the samples were re-weighted to determine LOI (LOI = (weight loss in furnace/dry weight) × 100).
Heavy metals
Metal content was determined using energy dispersive X-ray fluorescence. Cellulose film-lined sample holders were filled to a depth of c. 3 mm with dried and disaggregated sediment. Samples were placed, in sequence, under both Fe and Cd sources to determine the spectra of characteristic X-ray photons given off during a 250 s count time. The instrument was calibrated periodically during analysis using known reference materials. The data were finally processed using DECONV4 to resolve overlapping photon energies and to determine absolute concentrations.
Radiometric dating
Three bulk samples of organic-rich lake sediment were submitted for conventional or AMS 14C dating (dependent upon sample size). The results of these determinations are given in Table 1. The upper 90 cm of PL1998 was analysed for 210Pb, 226Ra, 137Cs and 241Am by direct gamma assay using an ORTEC HPGe GWL series well-type coaxial low background intrinsic germanium detector (Appleby et al. 1986). 210Pb was determined via its gamma emissions at 46.5 keV, and 226Ra by the 295 keV gamma-rays emitted by its daughter isotope 214Pb following 3 weeks of storage in sealed containers. 137Cs and 241Am were measured by their emissions at 662 and 59.5 keV, respectively.
Results
Geochemical data
Significant shifts in the nature of lake sedimentation may be observed from down-core trends in metal and organic content. Indeed, inspection of the LOI trend from PL2001 (Fig. 2) reveals two prolonged periods of increased organic content in the lower part of the record, at 600–550 cm and 500–375 cm, and a more short-lived phase at c. 240 cm.

Composite pollen diagram (PL1998 and PL2001) and LOI curve. Pollen concentrations are expressed as relative percentages of total land pollen for trees and shrubs and herbs, and relative percentages of total land pollen and aquatics for aquatics. The star shown on the PL2001 pollen diagram denotes the presence of Erodium cicutarium at 328 cm
According to Mayers (2001) heavy metal concentration data in PL1998 may be used to identify changes in sediment provenance (at a depth of 115 cm) brought about by the construction of a dam and an outfall pipe at the southern end of Pinto Lake, and a consequent increase in lake level, some time between 1948 and 1952 (from aerial photographic evidence). Similarly, down-core plots of lithogenic elements such a K, Ti and Zr concentration may be interpreted as temporal variations in mineral input, and when plotted relative to LOI or S reveal sudden increases or spikes in mineral content that may be related to short-lived periods of high catchment runoff. Whilst acknowledging that down-core variations in metal content may be a function of sedimentation rate as well as supply, here Ti concentration is employed as a simple proxy for mineral input which shows a strong negative correlation with LOI (r=−0.95). The observed trend (Fig. 3) confirms the presence of two periods of low mineral input (high organic content) below a general Ti content of c. 5 mg g−1 between depths of 600–550 cm and 500–375 cm. Furthermore, the uppermost 2 m of the sedimentary record are characterised by a subtle up-core decrease in Ti concentration—perhaps reflecting a gradual decrease in the supply of runoff-derived mineral matter.

Downcore plot of Ti concentration for PL2001
Pinto Lake pollen
The composite pollen record from both PL1998 and PL2001 provides evidence of significant vegetation change as a result of natural and anthropogenic influences on the lake-catchment ecosystem (Fig. 2). The sequence spans the time period of considerable botanical and land-use change in Central Coastal California, as well as recording changing marginal wetland communities due to temporal trends in climate and lake level. It is, therefore, surprising that the pollen spectrum changes very little through the sedimentary record. However, those changes considered to be of some significance in establishing a chronology with reference to the history of land use and in resolving the nature and forcing mechanisms of changing lake sedimentation are outlined below.
Changes in the composition of local terrestrial woodland
There is compelling evidence for a widespread decline of local T–C–T (Taxaceae–Cupressaceae–Taxodiaceae) woodland (i.e. Sequoia). A marked difference is apparent between the two cores and also at the base of PL1998. Corylus pollen is consistently present (though only at low levels) in PL1998 (<3%), but is noticeably more abundant in sediments that predate the decline of T–C–T woodland (PL2001). Of interest here is that the land around Pinto Lake and Corralitos Lagoon was named the ‘land of lakes and hazelnuts’ by the missionary Juan Crespi in the 1770s because of the abundance of ‘hazel’ trees (Mayers 2001). Although frequent in the lower portion of the sedimentary record, an increase in Quercus pollen observed in PL1998 after the reduction of local redwoods is indicative of a significant shift in woodland structure and landscape surrounding Pinto Lake. There is also potential evidence of earlier woodland disturbance in PL2001, such as at c. 350 cm where Lithocarpus (dwarf oak) and Quercus (oak) decline. This decrease is also accompanied by a decrease in organic matter. In terms of introduced or ‘exotic’ tree species, the first Eucalyptus grains are found at a depth of c. 190 cm in PL1998, with further increases in abundance at c.150 and 135 cm. Similarly, the occurrences of Tsuga (western hemlock) at c. 25 cm and Acacia (African thornbush) in the surface sample of the core are both indicative of recent planting.
Changes in the composition of riparian woodland
The occurrence of Alnus pollen in core PL2001 appears related to depositional periods of reduced organic sedimentation. The lowermost (615 cm) pollen sample is dominated by Alnus pollen (52% TLP) related to the basal minerogenic sediment. Alnus is low or absent in the lower organic phase (605–560 cm) but is seen to increase within the overlying minerogenic phase between 560–510 cm. Salix (willow) pollen grains, like Alnus, are derived from riparian and lake marginal communities. Salix pollen is present beneath 230 cm at a low abundance (<5%). Here, however, there is a marked increase in Salix to a peak (23%) at 185 cm in PL1998, from which a gradual decline is visible to the top of the core. Salix is often poorly represented in wetland/lake pollen diagrams (even from surface samples near a canopy of Salix). A change of this magnitude suggests a colonisation of lake-margin and/or stream floodplains.
Grass and sedge pollen
Grass (Poaceae) pollen is ubiquitous in the pollen diagram, representing the regional and local influx of terrestrial and wetland grass species. In addition, large (>37 μm) diameter grass pollen grains were found throughout the core. In the lower half of the core (below c. 300 cm in PL2001) increases in the frequency of grass and sedge pollen correspond with observed changes in organic content, suggesting periodic increases in the extent of marginal wetland communities. For example, the transition between the organic and minerogenic sediments between 400 and 375 cm is accompanied by an increase in both grass and sedge. This relationship is seen to a lesser extent in the lower organic phase. Cyperaceae (sedge) pollen will have almost certainly been derived from local marginal wetland communities. Sedge pollen also increases during phases of increased organic deposition in the core, most noticeably with the well-constrained LOI peak at 240 cm. This may be evidence of an event/short term phase of wetland expansion or inwash of organic sediments from marginal wetlands.
Herb pollen
The herb pollen spectrum is dominated by types typical of California understory, disturbed ground and wetland habitats. The difficulty of identification to species level means only a general description of herbaceous vegetation patterns can be given.
Artemesia (e.g. sagebrush) is present through the core. Asteraceae subf. Asteroideae (e.g. the daisy family) is the dominant pollen type in the sample at 296 cm that suggests an expansion of daisy numbers caused by growth on recently disturbed soils and inwash into the water body (Turner 2000). Another herb type often associated with disturbed soils is Chenopodiaceae, which is ubiquitous in the upper 390 cm but less so in the lower sediments. The change from organic to minerogenic sedimentation at c. 375 cm in PL2001 is followed by the sequential occurrence of non-native Plantago lanceolata (narrow-leaved plantain) that appear first at 350 cm, followed by Erodium cicutarium at 328 cm and Rumex acetosella (sheep sorrel) at 288 cm.
Aquatic pollen and Pediastrum
The dominant aquatic pollen type found in the core is Typha latifolia (broad-leaved cattail), though it is more prevalent in the upper 210 cm than below. Similar to the expansion of Salix, this shift indicates a hydrological change, but in this case the establishment of permanent flooded areas. Interestingly, below 250 cm Isoetes (quillwort) is found. Quillworts (e.g. Isoetes nuttallii) occupy vernal pools, drainage channels and lake margins. Nuphar (yellow pond lily) pollen indicates deep, clear water conditions in the lake. An increase in Nuphar is seen between 50 and 30 cm that may reflect the 1961 management decision to release Rotenone into the lake to remove invasive fish species. This may have led to improved water transparency as a result of a reduction in suspended sediment caused by benthic grazing species (i.e. carp). Menyanthes (i.e. M. trifoliate—buckbean) pollen shows a marked increase between depths of 70 and 15 cm. This is indicative of an expansion of wet sedge fen and pools in local wetland habitats (Griffen 1975).
The lack of knowledge on their ecological requirements and identification of types to species level hinder the interpretation of Pediastrum counts in the sedimentary record. In conjunction with the pollen and sedimentological data, we see that at a depth of 360 cm in PL2001 high Pediastrum counts are associated with low organic deposition and mineral inwash that seems to have resulted in eutrophic lake waters. Similarly, the increase in Pediastrum from a depth of 95 cm suggests increased eutrophic conditions and the expansion of marginal aquatics, as a result of the raising of the water level and flooding of the tributary arms of the lake from the early 1950s. Indeed, this is supported by the diatom assemblage data above c. 95–115 cm where there is evidence of planktonic diatom blooms at the detriment of benthic lifeforms (Mayers 2001). The decrease in Pediastrum observed at 65 cm signifies a change in lake water chemistry, which is associated with the increase in Nuphar reflecting a shift towards oligotrophic conditions. However, a return is observed at c. 35 cm, corresponding with increased Menyanthes pollen which suggests that recent Pediastrum has followed lake-margin macrophyte growth.
Chronology of sedimentation in Pinto Lake
Important chronological data are provided by both 14C and 137Cs chronologies. Details on the 14C ages are given in Table 1, providing well-resolved ages for organic-rich lake sediment immediately adjacent to three distinct stratigraphic transitions at 593, 563 and 499 cm. These ages span a time period extending from c. 650–1275 ad, characterised by two very contrasting phases of lake sediment accretion. In terms of the more recent chronology of lake sedimentation, only 137Cs was of sufficient activity in the uppermost 90 cm of PL1998 to provide a reliable means of dating, with unsupported 210Pb activity being below the limits of detection. Presumably, low unsupported 210Pb was due to the dilution of atmospheric fallout by rapid sediment accretion. The 137Cs activity versus depth profile (Fig. 4) shows a well-resolved peak between 72 and 88 cm, reaching 36 Bq kg−1. Above this, the activity trend shows a variable decrease up-core. Assuming a constant rate of sedimentation and that the 137Cs profile reflects the fallout rate (e.g. Pennington et al. 1973), the pronounced peak may be dated to the 1963 (±7) atmospheric fallout maximum. These radiometric age data are summarised in Table 2.

137Cs profile for PL1998
Further chronological marker horizons may be obtained for the 20th century from down-core trends in terrestrially-derived pollutants (Mayers 2001). For example, the down-core record of DDT concentration can be linked to the history of its manufacture and application as a pesticide on crops in fields adjacent to the waterbody of concern. Mayers (2001) noted that both peak use and a significant reduction in DDT can be identified from the record of DDT and its derivatives in PL1998. Here, the dates for initial increase (1940 onward) and the ban on the use of DDT in California (1970) can be assigned to the trends in ΣDDT observed at depths of 129–122 cm and c. 68 cm, respectively.
The pollen data give a series of chronological markers to which we may assign a variable degree of confidence and uncertainty dependent upon the available data on local history, vegetation change, land-use, hydrology and rainfall. For example, the reduction in T–C–T and Pinus and, to a lesser degree, Corylus and Alnus pollen in association with the subtle increase in Quercus at c. 230 cm appears to correspond to the known deforestation of redwoods around Pinto Lake from 1844 onwards when Rancho Corralitos was given over to logging. Alternatively, a slightly later date may be assigned from the 1850–60s which saw the expansion of logging on a commercial scale in the northern part of Monterey Bay (Gordon 1996) and the operation of eight sawmills in Santa Cruz County by 1860. Hence, the timing of this main phase of redwood clearance cannot be resolved more accurately than 1844–1860.
The earlier reduction in Quercus and Lithocarpus pollen at c. 350 cm, accompanied by the first occurrence of P. lanceolata and, subsequently, E. cicutarium and R. acetosella, may be linked to the establishment of Rancho Corralitos in 1821 or the larger ranchos of the Pajaro Valley from 1823 onward (Lewis 1994), which resulted in 33,000 acres of farmland in the valley by 1887. In the absence of pollen data at finer resolution or any independent dating of the occurrence of alien species associated with Mexican and subsequent Euro-American immigration, these data can only be resolved with confidence to the period of 1820–1860. However, an early destabilisation scenario may also be considered in which these putative indicators of early catchment disturbance are dated with reference to the regional history of immigration and disturbance. For example, seeds of E. cicutarium have been found in adobe bricks from the earliest missions—along with domesticated cereals. Mensing and Byrne (1998) date the first arrival of E. cicutarium to 1755–1760 in a core from the Santa Barbara Basin. Similarly, at San Joaquin marsh, Davis (1992) used the occurrence of E. cicutarium at a depth of 117 cm to give a date of c. 1776–1797, coinciding with the founding of the San Juan Capistrano and San Juan Bautista missions. Extensive disturbance enabled the continuation of alien species invasion. R. acetosella, a common weed found along roadsides and in pastures, was introduced during the Mexican era (1825–1848 ad) (Watson 1880) and became widespread due to grassland disturbance during the 1830s as a result of the spread of cattle ranches. Similarly, P. lanceolata was introduced by American settlers in the 1850s (Frenkel 1970) and became widespread in the 1860s (Mudie and Byrne 1980). In this case, the section of PL2001 between c. 350 and c. 325 cm is likely to date from the latter half of the 18th century, which saw the discovering of Pinto Lake by Don Gaspar de Portola in 1769 (Lydon 1994), and the establishing of the Mission in Santa Cruz in 1791. The early settlers established a pastoral lifestyle based on stock-rearing, agriculture and logging, and the land around Pinto Lake as a corral for keeping sheep and horses for the Santa Cruz Mission. From these pollen data, the first evidence of human impact may date from as early as 1769–1797, but more likely 1791–1823 (following the foundation of Santa Cruz mission but prior to the initiation of widespread arable agriculture on the Pajaro valley).
Eucalyptus is also an introduced species, and is now a dominant sight on the landscape. The plants were first sold in San Francisco as ornamentals from 1850 (Gordon 1996) and later became popular as shade providing trees and widespread planting began in 1860 across the State. The severe lack of hardwood through the 1870s, combined with a malaria outbreak (which Eucalyptus was believed to ward off) in the 1880s, initiated a second phase of planting on an unprecedented scale. The State Board of Agriculture began to request the establishment of artificial forests to compensate for land degradation across California. Due to the increased interest in the species, the period 1905–1912 was known as the Eucalyptus ‘boom’ (Santos 1997). Dating with Eucalyptus, however, is complicated by the fact that it is insect pollinated (Martin 1963) and would have to grow in close proximity to the lake in order for the pollen grains to be incorporated into the sediment profile. Thus, local planting histories may be more important than regional patterns of establishment. Local advertisements encouraging the planting of Eucalyptus date from 1909 (Outwest, Vol. 21, No.5 1909) and newspaper articles refer to the establishment of Eucalyptus groves in the Pajaro region in 1910 (Evening Pajaroan, February 23 1910), which corresponds to the aforementioned boom period. The occurrence of Eucalyptus pollen at c. 190 cm may, therefore, be linked to the first planting of Eucalyptus in the region between 1860 and 1910. The second increase in Eucalyptus at c. 150 cm depth, perhaps even that at c. 135 cm, probably dates from c. 1880–1935—based on the planting encouraged by the malaria outbreak and the earliest photograph found of the area. Further resolution of these ages is problematic due to the lack of detail on the timing and distribution of plantations at this time, which is, unfortunately, crucial information due to the fact that Eucalyptus pollen is a localised distribution. However, investigation of the aerial photographic record reveals considerable changes in the Eucalyptus canopy during the early part of the 20th century. Extrapolating the observed trend of increasing canopy area back to a point of origin using an historical sequence of aerial photographs, dates the planting of the main cluster of Eucalyptus trees to c. 1910–1920. Hence, the increase in Eucalyptus pollen at 153–135 cm can be further resolved to 1910–1935.
Those pollen markers that are considered to be well-constrained are given in Table 2. In addition, three potential ages (earliest, best estimate and latest) are given for the equivocal pollen evidence of early catchment disturbance. Although this evidence could equally be rejected as a chronological waypoint, the sequential occurrence of three pollen indicators of Mexican immigration (albeit with P. lanceolata occurring anomalously early in this sequence) provide strong evidence of imported land-use practices preserved in the sedimentary record between c. 350 and 325 cm.
From the chronological data given above, the rate of lake sediment accumulation may be investigated for the period of c. 650 ad to the present (Fig. 5). However, by far the majority of the chronological marker horizons (pollen, 137Cs and DDT) date from the late 18th century to the mid-1900s. Unfortunately, this bias towards the period of lake sedimentation following human impact makes it difficult to establish the timing and nature of the transition from the pre-impact condition. This is particularly significant for the period of early catchment disturbance where both the impact and timing of land-use change resulting from immigration are poorly resolved. The level of uncertainty associated with the pollen ages determined for the late 18th and 19th centuries is a consequence of limited specific detail in relating known regional trends in vegetation and erosion history to their expression at the local level. Despite these shortcomings, a good deal of correspondence can be seen within the early (14C) and late (‘post-clearance’ pollen, DDT and 137Cs) chronological data—irrespective of uncertainties in the history of early immigration and land use. This lends a considerable degree of credence to the links made between the pollen data and catchment history. Indeed, if the trends in sediment accretion from the early and late datapoints were to be extrapolated to a point of overlap, they would probably cross at the depth where the pollen show credible evidence of early catchment disturbance, i.e. c. 350 cm, but at a slightly earlier time. This would then be indicative of disturbance related to Mexican immigration at the earliest inferred time.

Bivariate plot of determined chronological marker horizons against cumulative dry mass (kg m−2) for PL2001. The slope of the trend gives a clear illustration of sediment accumulation rate both prior to and following human impact on the Pinto Lake catchment
Discussion
The pollen data confirm a significant degree of human impact on catchment vegetation and lake hydrology. Perhaps the clearest evidence of this is found at a depth of c. 230 cm in PL1998 where the marked decline in T–C–T pollen is accompanied by reductions in Corylus, Alnus and Pinus and an increase in Quercus followed by steady increases in Salix, Artemisia, Poaceae and T. latifolia. This is considered to be indicative of redwood and riparian woodland clearance in the catchment in 1844–1860, leading to the expansion of grassland and sagebrush, the opportunistic increase in understory woodland, and to an expansion of lake marginal wetlands as a result of the reduced catchment evapotranspiration. An earlier period of catchment disturbance is seen more in the presence of alien species than in unequivocal evidence of clearance. Whilst the increase in Chenopodiaceae, Ranunculus-type pollen and Pediastrum at c. 375 cm in PL2001 may be indicative of early human impact, as might a subtle change in composition of the woodland pollen spectrum at c. 350 cm, it is the sequential occurrence of large Poaceae, P. lanceolata, E. cicutarium and R. acetosella between c. 350 and c. 285 cm that provide less equivocal evidence of Mexican immigration. Although this period of limited human impact cannot be chronologically constrained by independent dating evidence, it appears to coincide with the documented discovery of Pinto Lake and founding of the Santa Cruz mission around 1769–1797 (although this requires early invasion by P. lanceolata).
Hence, the upper 3.5 m or so of the record recovered from Pinto Lake preserves evidence of a period of sedimentation from c. 1800 (Fig. 5). The model of sedimentation for Pinto Lake describes a roughly linear rate of sediment accumulation of c. 9 kg m−2 year−1 (c. 2 cm year−1) for the last 200 years or so, despite significant changes in land use and, more recently, catchment management during this time. Prior to this, there is evidence of a significantly slower rate of sedimentation (c. 0.7 kg m−2 year−1), lending support to an order of magnitude increase in the rate of sediment accumulation following on from immigration in the late 18th and early 19th centuries. Significantly, the evidence suggests that this increase in sedimentation rate has remained almost unabated to the present. Although measures to limit soil erosion have been put in place during the latter half of the 20th century, there has not been a significant reduction in the rate of sediment accumulation. However, again considering Ti concentration as a proxy for catchment erosion (Fig. 3), a subtle and progressive reduction in minerogenic content has taken place since about 1920. Diatom and Pediastrum evidence reveals that this reduction in mineral matter has probably been compensated by an increase in biological productivity within the lake and, hence, the overall sedimentation rate has not declined (Mayers 2001). This has important implications for the perceived benefits to lake infilling of measures to reduce soil erosion.
Significantly, however, the most recent ‘switch’ in the nature of lake sedimentation from more organic to minerogenic accretion appears to precede all the pollen evidence of human impact on the landscape, with the exception of increasing Chenopodiaceae and Ranunculus-type pollen and Pediastrum. Indeed, Pinto Lake has experienced at least three major changes in the nature of sedimentation prior to this. During periods of more organic sediment accumulation, Poaceae and Cyperaceae increase whilst Alnus decreases. From this, it may be inferred that the expansion of lake marginal wetlands is followed by hydroseral succession to alder communities or an increase in runoff-derived mineral input from alder-dominated riparian areas within the catchment. The sequence of change from minerogenic to more organic sediment accumulation may be attributed to one (or a combination) of three causes:
Lake level
As described in the earlier section on lake morphology, the tributary arms on the northern shores of Pinto Lake are characterised by shallow water (c. 1 m) and, as such, are highly sensitive to lake level change. In addition, their steep littoral shores and the marked break of slope between these arms and the main body of the lake basin (see Fig. 1b) impose morphological constraints on any ecological response. From the pollen data, increased organic content is generally accompanied by increasing grass and sedge pollen with no overall change in woodland taxa but with a reduction in Alnus. This association would suggest an increase in marginal wetland/aquatic communities related to a change in lake level and an expansion of these wetland environments in the tributary arms. However, the nature of lake level change resulting in this expansion would be dependent on the pre-existing condition and the amplitude of change. For example, a high lake level would act to drown these marginal wetland areas and restrict their distribution to the steep, narrow shores—thus limiting their opportunity to trap mineral input and leading to minerogenic lake sedimentation. A subtle fall from this level would increase the areal distribution of these wetlands and increase their potential to intercept runoff-derived mineral input to the lake, thus encouraging organic lake sedimentation. A further fall would expose these marginal areas and restrict the lake to the steep-sided main basin (minerogenic lake sedimentation returns). Hence, marginal wetland expansion occurs within a rather limited range of lake level.
Fire
Fire and other aboriginal land management practices would act to destabilise catchment soils and increase the supply of runoff-derived mineral matter. Although the ‘opportunistic’ existence of the Ohlone Indians clearly made best use of fishing and seasonally available resources, Keeley (2002) recognises that the Coast Ranges were regions of high Indian density and that fire would have been used to enhance cultural resources, increase seed, bulb and fruit production, increase available space for travel and for deer and wildfowl management, improve water resources, and reduce susceptibility to attack from wildlife. Indeed, a substantial proportion of the Central Coast landscape was converted from scrubland to grassland by the native American Indians, which was then sustained and/or expanded by the Euro-American settlers. Set within this context, it seems unlikely that grassland management by aboriginals would be expressed in the form of two distinct phases of decreased mineral input to Pinto Lake, particularly when the pollen data exhibit no corresponding change in the surrounding woodland (with the exception of decreased Alnus). Indeed, the observed increase in the proportion of grass and sedge and decreasing Alnus pollen during periods of organic sedimentation seems counter to a causal link between grassland development and enhanced runoff-derived mineral input.
Precipitation
Changes in precipitation on a timescale longer that that identified from historical and instrumental data may have a bearing on both lake level and runoff-derived mineral input from the catchment. From the data presented here, we propose two periods of reduced precipitation in the region, i.e. low runoff coupled with the expansion of shallow water marginal communities, dating from c. 650–900 and 1275–1750 ad. Lower annual precipitation would reduce the input of runoff-derived mineral matter from the catchment without any large-scale changes in the pollen spectra.
Further insight regarding the significance of changes in lake sedimentation prior to the late-18th century may be obtained from mass accumulation rates for particular metals over different time periods spanned by the chronological markers. Using Ti concentration as mineral matter proxy, and LOI and S concentration as organic matter indicators, temporal trends in mineral and organic sediment accumulation may be examined (Fig. 6). Although marked increases in all these proxies may be observed at c. 1800, it is Ti that exhibits that greatest proportional increase from about 1825 onward. The predominance of mineral input continues until c. 1940, at which point LOI also decreases but the accumulation of S is maintained. These data not only confirm the dramatic increase in lake sediment accumulation during the early 19th century, they are similarly indicative of a decrease in mineral input and an increase in biological productivity from the mid-20th century. Of equal significance is the fact that the amplitude of change in the mass accumulation of Ti, LOI and S is very limited during the period 650–1800 ad—during which time significant changes in the nature of lake sedimentation take place. From this, it can be inferred that only subtle (although prolonged) shifts in lake level and runoff may be sufficient to induce a significant change in the nature of lake sedimentation. Indeed, this is supported by the pollen evidence in that no large-scale changes in the woodland, herb or aquatic taxa are observed during this time. Against this background of high sensitivity, it is not surprising that imported land-use practices of Mexican and Euro-American immigration had such a dramatic impact.

Bivariate plots of mass accumulation against age for a organic matter (LOI), b sulphur concentration and c titanium concentration
Conclusions
In terms of the pre-impact condition, there is limited detail on the forcing of the rate and nature of lake sedimentation prior to the Mexican era. Pinto Lake appears to have been an environment where both sedimentation and lake level were driven by long-term climate change in which reduced precipitation leads to a subtle reduction in runoff and an expansion of shallow wetland lake margin communities—particularly in the four tributary arms at the northern end of the lake. The data presented here show the nature of lake sedimentation to be highly sensitive to these subtle changes in climate, but that the rate of sediment accumulation is more robust.
Set within this ‘pre-impact’ condition of high sensitivity to climate change, there is clear evidence of the impact of changing or ‘imported’ land-use on the rate of lake sedimentation. The impacts of Mexican and Euro-American ranching, agriculture and logging are reflected in the order of magnitude increase in the rate of lake sedimentation, both minerogenic and organic, after about 1825. Evidence of the impact of earlier immigration is difficult to detect and to constrain chronologically because it is set against a pre-existing background condition of minerogenic lake sedimentation, i.e. the occurrence of alien plant species generally associated with Mexican immigration post-dates the onset of increased minerogenic sedimentation.
The fact that the rate of lake sedimentation has remained unabated since the early 19th century, perhaps due to a replacement of reduced mineral inwash by increased biological productivity in more recent times, is indicative of the extent to which the sensitivity threshold controlling the nature of sedimentation has been exceeded by imported land-use practices. Furthermore, it seems that existing measures to regulate soil loss have been relatively ineffective in reducing the rate of lake infilling. Increasing sedimentation due to human impact has clearly reduced the lifetime and long-term resilience of the lake due to an enhanced rate of infilling. This has important implications for agriculture in the region, dictating the need for careful stewardship of land-use—not only from the perspective of soil erosion but also with regard to the transport and fate of fertilisers and pesticides and, indeed, the flux of sediment, nutrients and pollutants to the coast. The sensitivity of the lake is illustrated by the nature of lake sedimentation in response to climate change prior to human impact. In terms of resilience to human impact, however, the lake-catchment system has illustrated a high degree of sensitivity to land-use change—to the extent that once the disturbance condition was established, the enhanced sediment accumulation rate did not recover (despite recent erosion management).
In the context of future research, further work is required to fully resolve the control on the nature of lake sedimentation during the pre-impact condition. This may be achieved through diatom and charcoal analysis of PL2001 to determine changes in lake level and fire incidence. Further 14C dating coupled with high-resolution pollen analysis will also elucidate the chronology and nature of impact on the vegetation resulting from early Spanish and Mexican immigration. This latter phase of research will further resolve the cause of most recent switch in lake sediment accumulation from more organic to minerogenic sedimentation at c. 390–350 cm.
Our findings illustrate the importance of ecohydrology in understanding lake system response and resilience to climate and human impact, not simply through the well-established linkage between catchment land-use/cover and endo/exogenic sediment supply but through a more complex set of interactions in which this linkage is moderated by lake morphometry, lake level, the extent of shallow marginal wetland ecosystems, water quality and aquatic ecology. As a result, we consider ecohydrological principles to be of considerable significance in formulating future policy and management practices. All things being equal, the present rate of infilling gives Pinto Lake a lifetime of 250–400 years. As a leisure amenity, this lifetime is considerable and is significantly longer than that envisaged for many leisure facilities in Santa Cruz County. From an ecohydrological management perspective, infilling will result in considerable changes in lake margin habitat (as well as consequent changes in the nature of lake sedimentation), thus enhancing Pinto Lake’s amenity value as a wetland wildlife preserve. Sadly, as lake volume reduces, the sediment, nutrient and pollutant impacts of local land-use are likely to become more acute. In addition, with future climate change, Pinto Lake’s water budget will become increasingly threatened.
Themes of crucial relevance for future policy and planning are, therefore: the mosaic of land-use in terms of soil erosion potential; ecohydrological principles in managing wetland habitats as a means of moderating water, sediment, nutrient and pollutant flux; and water resource lifetime and balancing priorities in resource use.
References
Allen RH (1935) The Spanish land grant system as an influence in the agricultural development of California. Agric Hist 9(3):126–142
Appleby PG, Nolan PJ, Gifford DW, Godfrey MJ, Oldfield F, Anderson NJ, Battarbee RW (1986) 210Pb dating by low background gamma counting. Hydrobiologia 141:21–27
Battarbee RW (2000) Palaeolimnological approaches to climate change, with special reference to the biological record. Quaternary Sci Rev 19:107–124
Brugam RB (1978) Pollen indicators of land-use change in southern Connecticut. Quaternary Res 9:349–362
Brush GS (1989) Rates and patterns of estuarine sediment accumulation. Limnol Oceanogr 34(7):1235–1246
Brush GS (1994) The Chesapeake Bay estuarine system. In: Roberts N (ed) The changing global environment. Blackwell, Oxford, pp 397–416
Brush GS, Martin EA, DeFries RS, Rice CA (1982) Comparisons of 210Pb and pollen methods of determining rates of estuarine sediment accumulation. Quaternary Res 18:196–217
Cayan DR, Redmond KT, Riddle LG (1999) ENSO and hydrologic extremes in the western United States. J Clim 12:2881–2893
Cole KL, Liu G-W (1994) Holocene paleoecology of an estuary on Santa Rosa Island, California. Quaternary Res 41:326–335
Cooke RU, Reeves RW (1976) Arroyos and environmental change in the American South-West. Clarendon Press, Oxford, p 213
Davis MB (1976) Erosion rates and land use history in southern Michigan. Environ Conserv 3(2):139–148
Davis OK (1992) Rapid climatic change in coastal southern California inferred from pollen analysis of San Joaquin Marsh. Quaternary Res 37:89–100
Davies SJ, Metcalfe SE, MacKenzie AB, Newton AJ, Endfield GH, Farmer JG (2004) Environmental changes in the Zirahuen Basin, Michoacan, Mexico, during the last 1000 years. J Palaeolimnol 31(1):77–98
Diaz HF, Markgraf V (eds) (1992) El Niño. Historical and Paleoclimatic aspects of the southern oscillation. Cambridge University Press, Cambridge
Dupre WC (1975) Quaternary history of the Watsonville lowlands, North Central Monterey Bay region, California. Unpublished PhD Thesis, Stanford University, p 145
Eittreim SL, Xu JP, Noble M, Edwards BD (2002) Towards a sediment budget for the Santa Cruz shelf. Mar Geol 181:235–248
Ely LL, Enzel Y, Baker VR, Cayan DR (1993) A 5000-year record of extreme floods and climate change in the southwestern United States. Science 262:410–412
Engstrom WN (1996) The California storm of January 1862. Quaternary Res 46:141–148
Frenkel RE (1970) Ruderal vegetation along some California roadsides. University of California Publications in Geography, 20, University of California Press, Berkeley, p 163
Fu C, Diaz HF, Fletcher JO (1986) Characteristics of the response of SST in the central Pacific associated with warm episodes of the southern oscillation. Mon Weather Rev 114:1716–1738
Gordon BL (1996) Monterey Bay area: natural history and cultural imprints. The Boxwood Press, California, p 375
Griffen K (1975) Vegetation studies and modern pollen spectra from the Red Lake peatland, Northern Minnesota. Ecology 56:531–546
Grunwald et al (1974) Environmental impact report. Pinto Lake regional park, Santa Cruz County, F868-S3G, 78
Jarrell R (1982) History. In: Warrick SF (eds) The natural history of the UC Santa Cruz Campus. Publication no.11, Environmental Field Program, University of California, Santa Cruz, pp 1–33
Johnson DL (1977) The origin of island mammoths and the quaternary land bridge history of the northern Channel Islands, California. Quaternary Res 10:204–225
Keeley JE (2002) Native American impacts on fire regimes of the California coast ranges. J Biogeogr 29:303–320
Kiladis GN, Diaz HF (1989) Global climatic anomalies associated with extremes in the Southern Oscillation. J Clim 2:965–976
Knox JC (2000) Sensitivity of modern and Holocene floods to climate change. Quaternary Sci Rev 19:439–457
Lantis DW (1970) California: land of contrast, 2nd edn. Wadsworth Publishing Company Inc, Belmont
Lewis B (1994) Watsonville remembered – a selection of KOMY-radio broadcasts. Santa Cruz County History Journal 1, Santa Cruz Historical Trust Inc
Lydon S (1994) Toward rediscovering the Monterey Bay region and its history. Santa Cruz County Hist J 1:4–8
Mannion AM (1995) Agriculture and environmental change: temporal and spatial dimensions. Wiley, Chichester
Martin PS (1963) The last 10,000 Years. A fossil pollen record of the American Southwest. University of Arizona Press, Tucson
Mayers C (2001) An environmental history of the recent human and climatic impacts on Central Coastal California: the Record from Pinto Lake, Santa Cruz County. Unpublished PhD Thesis, University of Liverpool, p 287
McCabe GJ, Dettinger MD (1999) Decadal variations in the strength of ENSO teleconnections with precipitation in the western United States. Int J Climatol 19:1399–1410
Mensing S, Byrne R (1998) Pre-mission invasion of Erodium cicutarium in California. J Biogeogr 25:757–762
Michaelson J, Haston L, Davis FW (1987) 400 years of central California precipitation variability reconstructed from tree-rings. Water Resour Bull 23(5):809–818
Mudie PJ, Byrne R (1980) Pollen evidence for historic sedimentation rates in California coastal marshes. Estuarine Coastal Mar Sci 10:305–316
Patch D, Jones T (1984) Paleoenvironmental change at Elkhorn Slough: implications for human adaptive strategies. J Calif Gt Basin Anthropol 6(1):19–43
Pennington W, Cambray RS, Fisher EM (1973) Observations on lake sediments using fallout 137Cs as a tracer. Nature 242:324–326
Redmond KT, Koch RW (1991) Surface climate and streamflow variability in the western United States and their relationship to large-scale circulation indices. Water Resour Res 27:2381–2399
Robbins JA, Edgington DN, Kemp ALW (1978) Comparative 210Pb, 137Cs, and pollen geochronologies of sediments from Lakes Ontario and Erie. Quaternary Res 10:256–278
Santos RL (1997) The Eucalyptus of California. Alley-Cass Publications, California
Schonher T, Nicholson SE (1989) The relationship between California rainfall and ENSO events. J Clim 2(11):1258–1269
Turner S (2000) High resolution palaeoenvironmental analyses of coastal wetland sediments from south east Sicily. Unpublished PhD Thesis, Brunel University
Watson S (1880) California Geological Survey: vol 1, Botany. Harvard University Press, Cambridge
Wolanski E, Boorman LA, Chicharo L, Langlois-Salou E, Lara R, Plater AJ, Uncles RJ, Zalewski M (2004) Ecohydrology as a new tool for sustainable management of estuaries and coastal waters. Wetland Ecol Manag 12:235–276
Zalewski M (2002) Ecohydrology—the use of ecological and hydrological processes for sustainable management of water resources. Hydrol Processes 47:823–832
Acknowledgements
The authors would like to thank Peter Appleby for undertaking the gamma-spectrometry in the Environmental Radioactivity Research Centre, University of Liverpool. Scott Anderson is thanked for his kind assistance with the identification of the pollen, and Gerry Weber for help with the fieldwork and providing essential equipment. Chris Spencer and Richard Mourne are acknowledged for their assistance with the collection of PL2001—which was made possible through financial assistance from the University of the West of England as well as the University of Liverpool. Thanks are also extended to Traci Roberts, Rob Ketley, Burton Gordon and Sandy Lydon for their assistance concerning the environmental history of Pinto Lake, and to the numerous field assistants including: Dave McKnight, Andy Morse, Katie Willis and Matt Wright. Sandra Mather is also acknowledged for the final production of the figures, and Hilda Hull, Irene Cooper and Bob Jude for their technical support. This research formed the basis of a PhD studentship completed by Claire Mayers, which was funded via the Department of Geography, University of Liverpool, Dumkow Scholarship.
Author information
Authors and Affiliations
Corresponding author
Additional information
An erratum to this article can be found at http://dx.doi.org/10.1007/s10113-006-0016-y
Rights and permissions
About this article
Cite this article
Plater, A.J., Boyle, J.F., Mayers, C. et al. Climate and human impact on lowland lake sedimentation in Central Coastal California: the record from c. 650 ad to the present. Reg Environ Change 6, 71–85 (2006). https://doi.org/10.1007/s10113-006-0013-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10113-006-0013-1