
Abstract
Object categorization is an important cognitive adaptation, quickly providing an animal with relevant and potentially life-saving information. It can be defined as the process whereby objects that are not the same, are nonetheless grouped together according to some defining feature(s) and responded to as if they were the same. In this way, knowledge about one object, behavior or situation can be extrapolated onto another without much cost and effort. Many vertebrates including humans, monkeys, birds and teleosts have been shown to be able to categorize, with abilities varying between species and tasks. This study assessed object categorization skills in the gray bamboo shark Chiloscyllium griseum. Sharks learned to distinguish between the two categories, ‘fish’ versus ‘snail’ independently of image features and image type, i.e., black and white drawings, photographs, comics or negative images. Transfer tests indicated that sharks predominantly focused on and categorized the positive stimulus, while disregarding the negative stimulus.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The ability to place objects or items into discrete mental units which are treated equivalently is called categorization (Mervis and Rosch 1981). In the process, similarities among various objects and items are recognized and categories constructed that symbolize shared features (Spinozzi 1996). To be able to categorize is an essential skill for many invertebrate and vertebrate species when distinguishing between and identifying prey, predators, conspecifics or heterospecifics and for behaviors such as identifying new objects, solving problems and obtaining environmental information (Spinozzi 1996; Smith 2014). It is therefore one of the most basic ways to organize knowledge (Smith and Medin 1981). Usually, there are countless ways in which to categorize objects; for example, a shark could be categorized as a predator, a fish or an animal, as dangerous or scary or according to its scientific family, to name just a few (Richler and Palmeri 2014). It is obvious from this one example that categorization can include several different abstraction levels, natural as well as artificial ones.
A wide range of experiments has already been performed on categorization abilities of humans, primates and birds (for a review see Smith et al. 2012a, b; Smith and Minda 1998; Berg et al. 2014). Birds were the first non-human animals to be assessed in this respect (e.g., Lubow 1974; Aust and Huber 2001; Cook and Smith 2006; Berankova et al. 2014; Castro and Wasserman 2014). In one of the pioneering experiment, pigeons (Columba livia) were trained to distinguish between the presence and absence of humans on photographs and pecked for reinforcement only when provided with the former (Herrnstein and Loveland 1964). More recently, different levels of categorization, such as classifications based on basic (e.g., categories such as ‘cars’, ‘chairs’, ‘flowers’ or ‘humans’) and superordinate (e.g., categories such as ‘natural’ or ‘artificial’) levels (Lazareva et al. 2004) as well as many others (e.g., Huber 2001) followed. Studies on primates began in the 1980s, primarily on Macaques (Macaca mulatta, Macaca fuscata) (Yoshikubo 1985; Murai et al. 2004; Basile and Hampton 2013), squirrel monkeys (Saimiri sciureus) (Philips 1996), capuchin monkeys (Cebus apella) (D’Amato and Van Sant 1988; Spinozzi et al. 2004; Smith et al. 2012a), baboons (Papio anubis) (Bovet and Vauclair 1998) and chimpanzees (Jitsumori and Matsuzawa 1991; Vonk and MacDonald 2002; Spinozzi 1995). Jitsumori and Matsuzawa (1991) showed that taxonomic and intraspecific differences exist between monkeys and birds when confronted with different views or silhouettes of bodies and faces of humans and non-human species. While pigeons seemed to base categorization on human specific features, monkeys used more generalized traits. In another study, both pigeons and monkeys learned to identify concrete categories (e.g., choosing a picture of a specific bird species over a non-bird picture) better than abstract ones (e.g., choosing a bird picture over a non-bird animal picture or choosing an animal picture over a non-animal picture) (Roberts and Mazmanian 1988). In this study, pigeons also learned faster than monkeys. In a review, Smith et al. (2012b) also described fundamental differences in the categorization abilities of humans, monkeys and pigeons, with the former two learning one-dimensional rule-based tasks more quickly than two-dimensional information-integrated tasks, while pigeons learned both equally fast. Humans were also faster and chose more accurately than pigeons in categorization tasks that were facilitated by selective attention (Berg et al. 2014).
There are several models or theories as to how categorization may proceed; for example, categorization could be based on logical rule learning (based on similar properties), the construction of prototypes (where an unknown object is compared with a specific, mentally stored prototype) or exemplar learning (in which new information is categorized according to a selection of already stored memories within a category and how well it matches them) (Richler and Palmeri 2014; Smith 2014). Additionally, there are other more complex theories that combine several of these elements, such as the ‘cluster formation’ theory or the ‘rule plus exception’ model (Richler and Palmeri 2014).
Cichlids of the genus Pseudotropheus have previously been to shown to be able to discriminate between two-dimensional perceptual object categories (Schluessel et al. 2012). Individuals associated positive stimuli with being correct and negative stimuli with being incorrect choices, indicating that not only one but two categories were formed in the process of training. Results were supported by experiments showing that when a new stimulus unrelated to either of the trained categories was presented in combination with the negative or incorrect one, fish significantly often refrained from making a choice altogether (Schluessel et al. 2012). Follow-up studies showed that the ability to categorize also applied to three-dimensional objects (Schluessel et al. 2014a), but not to 2D symmetrical and asymmetrical symbols (Schluessel et al. 2014b). While sharks (Chiloscyllium griseum) performed significantly above chance level in categorization experiments on symmetry perception, cichlids performed rather poorly, with very few individuals learning, and then only on some of the tasks (Schluessel et al. 2014b). Quite probably though, this result was caused by an inability of the cichlids to perceive symmetry rather than an inability to categorize symmetrical and asymmetrical features, a notion that was supported by the fact that cichlids also showed no spontaneous preference for symmetry, while sharks did.
The aim of this study was to test the ability of bamboo sharks (C. griseum) to build and distinguish between two basic mental categories of two-dimensional visual objects. In recent years, more studies have begun to investigate the cognitive abilities of elasmobranchs (for a review, see Schluessel 2014), which, for a long time, were thought to be less advanced than those of teleosts or other vertebrates. Results of these studies indicate that bamboo sharks can visually discriminate a range of symbols and objects and orient visually according to landmarks (Fuss et al. 2014a, b; Schluessel and Bleckmann 2012). Additionally, bamboo sharks can perceive optical illusions in the same way as humans and discriminate and categorize artificial stimuli such as symmetrical and asymmetrical symbols (Fuss et al. 2014c; Schluessel et al. 2014a, b). Juvenile sharks in general are often subject to predation and usually feed on various benthic organisms and fish. Juvenile lemon sharks can distinguish between conspecifics and heterospecifics (Guttridge et al. 2009), and juvenile bamboo sharks inhabit various habitats such as mangroves, sea beds and coral reefs. Considering these information, the ability to categorize would be advantageous for sharks to have and expected to be present in juvenile C. griseum, as well as other shark species. C. griseum was chosen as the experimental animal due to its previously shown learning ability and high motivation levels. Apart from testing categorization abilities, the present study also aimed to determine whether sharks just learn the significance of the food-rewarded or ‘correct’ category as opposed to both categories (rewarded and unrewarded).
Materials and methods
Subjects
Subjects were nine juvenile gray bamboo sharks obtained from the Zoo ‘Haus des Meeres’ in Vienna. Individuals were bred in captivity and ranged between 20 and 35 cm in total length. Sharks were housed in pairs in interconnected 250- to 400-L aquaria (total volume approximately 4,000 L) that were filled with aerated and filtered saltwater (~50 µS) and kept at a temperature of ~25 °C. Food (shrimp, fish and squid) was only provided during experimental trials. Experiments were conducted during daylight hours; there was a natural light: dark cycle. Three animals died unexpectedly after finishing the second/third single-pair discrimination, and one shark was excluded from the last experiment (C2) as it ceased participation (swimming and feeding) for a long time but recovered again later on. The data collected for these four sharks prior to leaving the experiments were included in the analysis.
Apparatus
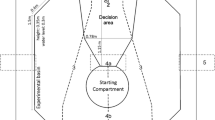
Experiments were performed using the same octagonal experimental basin and setup as outlined in Fuss et al. (2014a) (Fig. 1). Water depth was approximately 0.3 m. To exclude uncontrolled cueing and potentially disturbing external influences, the basin was surrounded by a white canopy (3.0 m × 3.0 m × 2.5 m). Ceiling mounted fluorescent tubes allowed an even illumination during the experiments (above canopy roof; Osram L 18 W, Lumilux Cool White, Germany).

Within the keyhole-shaped, gray PVC setup, a light gray guillotine door confined a starting compartment (0.43 m × 0.3 m × 0.35 m), in which sharks were placed before each trial. The experimenter was always situated behind the starting compartment and operated the guillotine door manually and remotely using a cable pull. After leaving the starting compartment, the sharks entered the cone-shaped experimental arena, terminated by a frosted screen on the opposite side of the starting compartment. A 0.33-m-long divider, attached to the frosted screen separated a left from a right division (Fig. 1), thereby allowing for an unambiguous decision making in response to the two stimuli displayed on either side of the frosted screen via a LED projector. As bamboo sharks usually swim close to the ground, stimuli were projected at a height of 3 cm above the basin floor. Feeders were installed just above the two projected stimuli, allowing food to be dropped into the setup remotely from either of the two feeders from the position of the experimenter.
Experimental procedure
For a correct choice to be recorded, sharks had to press their nose against the positive stimulus. Both feeders were baited during all trials to exclude unintentional cueing. Additionally, the water in the maze was stirred after each trial to preclude the accumulation of any olfactory cues on either side. Sessions were conducted once a day, always at the same time. Each session consisted of ten trials. For each session, the mean trial time and the percentage of correct choices was recorded. Trial time was defined as the time it took the shark to swim through the guillotine door and push against one of the two stimuli. It was recorded using a stopwatch. Training ended for each shark when the learning criterion was reached, which was defined as choosing the positive stimulus at least on seven out of ten trials (≥70 % correct choices) in three consecutive sessions. For each trial, the position of the positive stimulus was randomly determined prior to the experiments (five times on the right and five times on the left side); however, no single stimulus or stimulus category was shown more than twice consecutively on the same side. Intertrial time was 1 min. A trial was terminated but repeated if the shark did not make a choice within 2 min after raising the guillotine door. After three unsuccessful trials, a session was terminated. To prepare the sharks for subsequent transfer trials, which were interspersed with regular trials but were unrewarded, food was only provided in eight out of ten correct trials in the sessions following successful training. Prior to each session it was randomly determined which trials remained unrewarded (regardless of the shark’s actual choice). Not rewarding some of the regular trials was intended to prevent the sharks from realizing that only transfer trials were unrewarded and therefore not worth participating in. After each choice, the shark either returned to the SC by itself or was ushered back. Selected sessions were videotaped.
Pretraining
Prior to training, sharks had to get used to the apparatus and to feeding in the set up. Once sharks had learned to swim through the guillotine door, approach the food holders and associate them with food, training commenced.
Form discrimination
Training
In these two experiments, sharks (N = 9) had to learn to discriminate between a positive and a negative stimulus. Experiments served to test for the general motivation and learning ability of individuals. Sharks were first shown the stimulus combination ‘square’ versus ‘blank’. After reaching the learning criterion, sharks were then presented with the combination ‘square’ versus ‘circle’. The positive stimulus was always the square. After successful training, sharks commenced with the categorization training.
Object categorization
All stimuli shown during the object categorization experiments were obtained from the internet and reformatted/manipulated with the help of Microsoft Paint, Paint.net and Gimp©.
Single-pair discriminations
Sharks (N = 9) first had to perform in three single-pair discriminations, distinguishing between single images of fish and snails as representative objects for two randomly chosen basic mental categories, i.e., ‘fish’ and ‘snail’, to be tested later on. For these single-pair discriminations, black and white line drawings were used (Fig. 2). ‘Fish’ was always used as the positive stimulus. Care was taken to assure that picture pairs were similar in line thickness, amount of coloring etc. in order to avoid or minimize correct identification on the basis of any other characteristics but ‘fish’ and ‘snail’ itself.

Stimuli shown in the three single-pair discrimination experiments. Stimulus pair (a) was shown first (until the learning criterion was reached) followed by (b) and then (c) ‘fish’ was always the positive stimulus
Object categorization
Two categorization experiments were run to test whether sharks (N = 6) could discriminate between images of objects within the two categories ‘fish’ and ‘snail’, independent of single object features. Experiments only differed in regards to the pictures that were shown to the sharks. In C1 categorization trials, pictures consisted of black and white drawings, photographs, comics and negative images of snails and fish (Fig. 3). The two stimuli shown on any particular trial were always of the same type (Fig. 3), e.g., a black and white line drawing of a fish was shown against a black and white drawing of a snail. New stimulus pairs were shown on each trial. Within the two categories ‘fish’ and ‘snail’, care was taken that all positive (or all negative) objects shared no mutual features but differed in body structure, line strength, coloring (black and white components) and contrast, again to avoid correct identification on the basis of any other characteristics but ‘fish’ and ‘snail’ itself. In C2 categorization trials, only five sharks participated. Pictures also consisted of black and white drawings, photographs, comics and negative images of snails and fish (Fig. 4). However, here, pictures shown on any particular trial were randomly taken from any type, e.g., a real photograph of a snail was shown against a black and white drawing of a snail. Again, new stimulus pairs were shown on each trial.

Exemplary stimulus pairs shown during the first categorization experiment C1. A new stimulus pair was presented on each trial. Stimulus pairs featured different image types (e.g., photo, cartoon); however, stimuli within a pair were of the same type. ‘Fish’ was always the positive stimulus

Exemplary stimulus pairs shown during the second categorization experiment C2. A new stimulus pair was presented on each trial. Stimulus types varied within and between pairs. ‘Fish’ was always the positive stimulus
Transfer tests
Transfer tests were conducted to determine if the sharks could discriminate between one of the known objects (fish or snail) and an unknown object, i.e., in this case the image of either a plant, a jellyfish or a crustacean. The aim was to determine if sharks associated ‘fish’ with ‘being correct’ and/or ‘snail’ with ‘being wrong’. Also, in one type of test, two unknown objects were presented. Each session consisted of ten trials plus two transfer trials, which were randomly interspersed with regular trials. There were five types of transfer tests in total, i.e., ‘fish’ versus ‘plant’, ‘snail’ versus ‘plant’, ‘fish’ versus ‘jellyfish’, ‘snail’ versus ‘jellyfish’ and ‘jellyfish’ versus ‘crustacean’. The first two types were presented to each shark ten times each (whereby a different plant was shown on each trial), the other three types were presented only five times (there was only one picture of a jellyfish and of a crustacean), totaling 35 transfer tests for each individual shark (N = 5). The two transfer tests shown within a session were not necessarily of the same type and combinations were randomly varied.
Data analysis
The learning criterion was established to be ≥70.0 % correct choices on three consecutive sessions. The probability of achieving ≥70.0 % correct choices on three consecutive sessions by accident within a maximum of 30 sessions is <0.05 % (according to exact probability calculations; personal communication from R. Berthe). The following statistical tests were used for each individual fish (the criterion of statistical significance was p < 0.05): Yates Chi-square tests (χ 2) (including 95 % confidence intervals) were performed in order to test whether individual sharks chose the positive stimulus significantly more often than the wrong one during the transfer trials. A Wilcoxon signed-rank test was used to compare the performance of individuals (number of sessions and time) during the form discrimination and single-pair discrimination experiments, as well as object categorization training.
Results
Form discrimination
In the first form discrimination task, all sharks (N = 9) learned to discriminate between a square and a ‘blank’. Sharks needed between 4 and 25 sessions, with a mean of 17.9 ± 5.9 sessions (median 19) to reach the learning criterion. The mean trial time was 10.6 ± 9.6 seconds (s). In the second experiment, all sharks (N = 9) also learned to discriminate between a square and a circle; individuals took between 3 and 13 sessions with a mean of 6.8 ± 3.6 sessions (median 5). Sharks needed significantly fewer sessions in the second compared with the first task (t = 3.83, df = 8, P = 0.005). The mean trial time was 9.1 ± 6.5 s. Accordingly, all sharks went on to participate in the single discrimination experiments.
Single-pair discrimination
All but one shark reached the learning criterion in all three tasks. In the first and second single-pair discrimination task (Fig. 2), all sharks (N = 9) learned to discriminate between a fish and snail. In the first task, sharks needed between 3 and 13 sessions, with a mean of 9.6 ± 7.4 sessions (median 8) to reach the learning criterion. The mean trial time was 8.5 ± 5.9 s. In the second task (Fig. 2), sharks needed between 4 and 14 sessions with a mean of 8.8 ± 3.9 sessions (median 7). The mean trial time was 7.6 ± 4.1 s. Only eight animals completed the third task; of these eight, only seven reached the learning criterion. Sharks needed 4, 4, 4, 7, 9 and 39 sessions, with a mean of 11.1 ± 11.3 sessions (median 7). The mean trial time was 7.4 ± 3.8 s. Sharks needed approximately the same amount of sessions in all three tasks; there was no statistical difference between any pair of tasks (t = −0.053 to 0.27, 0.80 < P < 0.96).
Object categorization 1 (C1)
Sharks were presented with a new and unknown pair of fish/snail images on each trial (Fig. 3). Stimuli presented within a pair were of the same image type. All sharks (N = 6) reached the learning criterion. There was large intraspecific variation; most sharks needed less than ten sessions, i.e., 3, 6, 6, 7 and 9 sessions, while one shark needed 26 sessions to reach criterion. The mean was 9.5 ± 7.7 sessions (median 6.5); the mean trial time was 6.6 ± 3.0 s.
Object categorization 2 (C2)
Sharks were presented with a new and unknown pair of fish/snail images on each trial (Fig. 4). As opposed to C1, stimuli within a pair were not of the same image type. All sharks (N = 5) reached the learning criterion within 18 or fewer sessions. Again, there was large intraspecific variation, sharks needed 3, 3, 6, 12 and 18 sessions, respectively. The mean number of sessions needed was 8.4 ± 6.0 (median 6); the mean trial time was 6.6 ± 3.0 s.
Different sharks performed poorly in C1 and C2, e.g., the shark that needed 26 sessions for C1 only needed three sessions to complete C2, while the only shark that did not learn the third single-pair discrimination, took only seven sessions to learn C1 but 18 sessions for C2.
Transfer tests
Throughout the sessions, in which transfer tests were conducted, sharks maintained a mean performance of 71.0 ± 2.2 % correct choices for all regular (non-transfer) trials. The mean trial time ranged between 5.51 and 7.5 s for the five individuals. The results of the transfer test trials are shown in Fig. 4. When presented with the image of a fish (previously ‘correct’ and rewarded) and the unknown object ‘plant’, sharks still chose the fish stimulus almost as often as in training, in 67 % of trials, and this trend approached significance (sum of all χ 2 = 9.3, df = 5; P = 0.097). When presented with the image of the snail (previously ‘incorrect’) though, sharks did not prefer either one and chose according to chance (sum of all χ 2 = 2.1, df = 5; P = 0.84). Subsequently, sharks were presented with three additional types of transfer tests which were presented in a random order. Sharks chose according to chance between the previously ‘negative’ stimulus and an unknown object (snail/jellyfish; sum of all χ 2 = 4.0, df = 5; P = 0.55) and between two unknown objects (crustacean/jellyfish; (sum of all χ 2 = 3.2, df = 5; P = 0.67). Despite choosing the fish image in 68 % (17 out of 25) over the jellyfish in the third type of transfer test (previously ‘positive’ stimulus/unknown object), results in this transfer test were also not significant (sum of all χ 2 = 4.0, df = 5; P = 0.55). The mean trial time in the transfer tests ranged from 5.7 to 8.2 s per shark and was not significantly different from the mean trial time during the regular trials, which ranged from 5.5 to 7.5 s (Z = −2.92 E−103, P = 1.0). For individual results, please refer Table 1.
It is worthwhile mentioning the performance of one shark individually. It chose the formerly positive image of the fish over the plant (negative) and the jellyfish (unknown) image 100 % of the time (i.e., in 15 trials). Furthermore, it chose according to chance when presented with the snail/plant combination (50 %) but in subsequent trials chose the jellyfish 100 % over the snail image (formerly negative).
Discussion
Like teleosts, sharks (C. griseum) can form and discriminate visually between two two-dimensional object categories as for example between images of the organisms ‘fish’ and ‘snail’. However, while cichlids learned both the significance of the positive (rewarded) and the negative (unrewarded) stimulus (Schluessel et al. 2012), thereby forming two mental categories, sharks seemed to focus primarily on the former.
Sharks needed a mean of 18 sessions for the first form discrimination experiment, i.e., distinguishing between a square and a blank, but only seven sessions for the second, i.e., distinguishing a square from a circle. These findings confirmed results of previous studies on the visual discrimination ability of this species (Fuss et al. 2014c; Schluessel et al. 2014b), where sharks also needed significantly more sessions for the first discrimination task than for subsequent ones (despite those being more challenging). The decrease in sessions per task following the first task is usually related to the animals becoming familiar with the procedure. However, depending on the types of stimuli presented, other factors including memorization of the positive stimulus (if it remains the same) and the development of learning concepts can contribute to decreasing session numbers and trial times. According to these results, all animals proved suitable for subsequent categorization testing.
In the single-pair discriminations that followed, sharks needed approximately the same amount of sessions for the successful discrimination of the first ‘fish–snail’ stimulus pair (10 sessions) as for the second (9 sessions) or third (11 sessions). The numbers of sessions needed were similar to the mean number of sessions needed by cichlids (11) to distinguish between very similar line drawings of a fish and a snail (Schluessel et al. 2012). While the number of sessions the sharks needed to discriminate between three symmetry–asymmetry pairs in a previous study was much higher, i.e., 21, 23 and 25 sessions (Schluessel et al. 2014b) than in the present study, there was little variation in the relative number of sessions needed for the various tasks (range of 21–25). Similar results were obtained in the current study (range of 9–11). The results of the three studies show that approximately double the amount of sessions was needed by sharks and cichlids when two very artificial stimuli had to be distinguished, i.e., symmetrical and asymmetrical symbols, as opposed to differentiating between more realistic objects, such as fish and snail images.
Theoretically, the single-pair discriminations could have been mastered by sharks using selected features or components of the individual fish and snail images within a pair (such as one object is rounder, has a tip, is darker on the top, has a line on the left, etc.). However, this would not have been possible in the subsequent categorization experiments. Care was taken to assure that neither fish nor snail images shared recurring mutual image features such as, for example, the same line thickness or color, which would ‘give away’ the category (of course, object features such as fins on fish were recurring but these were identifying features for that category). As expected, all sharks reached the learning criterion in both categorization experiments indicating that discriminations were not just based on single image elements, but on category specific features or rules, identifying fish and snails, respectively. Previous studies in cichlids and sharks have shown that categorization learning occurred rapidly, proceeding at a much quicker pace than learning of single-pair discriminations (Schluessel et al. 2012, 2014a, b). With the exception of three instances, this was confirmed in the present study; most sharks needed less than ten sessions, and in several instances only three sessions (the minimum to fulfill the learning criterion) to complete both categorizations. Why three sharks needed 12, 18 or even 26 sessions for the completion of C1 or C2 could not be determined (this, however, greatly increased the overall session mean). Possibly, these individuals had differentiated between the single stimulus pairs based on image-specific features and had to shift strategies once they were constantly presented with new images, while the rest of the sharks had already started to do so prior to the categorization experiments. Effects of feeding as a cause for the variable performances can almost certainly be excluded, as can be experimental fatigue. Satiation rarely occurred before the end of a session and most animals would even continue (successful) participation beyond satiation. Trial times remained stable in the sharks that needed increased amount of sessions and despite performing below criterion, motivation levels (as indicated by swimming and feeding) did not decrease. There is generally large intraspecific variation regarding the performance of sharks and the session numbers recorded in the current study fall within previously observed ranges. At present, this seems like the most reasonable explanation for the discrepancy in session numbers, but there may have been other contributing factors.
It is unlikely that any of the sharks possessed an unconditional attraction (or spontaneous preference) for the symbol ‘fish’ as individual learning curves should have been much steeper in such a case. However, even if sharks had preferred ‘fish’ over ‘snail’ to begin with, the categorization experiments would have still shown that all fish images were consistently considered as ‘fish’ despite their variability and therefore, would have still shown the ability of sharks to categorize.
The results of the transfer tests were surprising, considering the performance of sharks during training and in the regular trials thereafter. Only one of the sharks chose the fish image over both unknown images (plant and jellyfish) in 100 % of all trials. Most of the other sharks chose the fish image in more than 50 % of the trials, but not significantly often. None of the sharks significantly preferred either of the unknown objects to the snail image, as had been expected due to results previously collected on cichlids. Instead, sharks chose according to chance. This suggests, that sharks must have recognized ‘fish’ as being ‘positive’ or ‘correct’, but that ‘snail’ was not necessarily associated with being ‘wrong’ or ‘incorrect’. Despite the fact that we attempted to prepare sharks for the transfer tests by not rewarding selected regular trials, the possibility remains that sharks eventually learned that transfer tests were unrewarded and subsequently chose according to chance or some other criterium irrespective of the learned categories. However, considering the performance of Shark 1, which exclusively chose the fish image, and considering a rather patchy distribution of correct and incorrect transfer tests, this seems unlikely. Sharks are generally quite curious and may have simply approached the unknown or novel stimuli out of interest as opposed to making an actual choice (‘novelty effect’). There was also no perceivable difference between showing different versions of an unknown object (such as several different plant images) or always showing the same image (as in case of jellyfish and crustacean). Lastly, only six individuals were trained in all tasks, and the number of transfer tests was therefore limited; it cannot be excluded that if more sharks had been assessed results would have been different and more straightforward to interpret.
The mean trial time decreased continuously throughout the experiments, from 10.6 (square/nothing) to 9.1 s (square/circle) to 8.4, 7.6 and 7.3 s in the three single-pair discriminations and from 6.6 to 6.5 s during C1 and C2 trials to 5.6 s in the regular trials during the transfer tests. The same decreasing trend applied to the standard deviations, indicating that as sharks performed better and more often, they also performed faster. The fact that trial time during the transfer tests did not significantly differ from regular trials, indicates that sharks were not disturbed or confused by the presence of unknown objects. On the contrary, cichlids, when presented with an unknown stimulus in combination with a positive stimulus, significantly often chose the positive one, but most often made no choice at all when presented with the combination of a negative and an unknown stimulus (Schluessel et al. 2012). This was further emphasized by very high trial times (Schluessel et al. 2012) and clearly indicated that cichlids had learned both types of categories, positive as well as negative.
Unlike many other studies, the present experiment used a two-alternative forced-choice procedure, in which the two stimuli to be discriminated were presented simultaneously instead of being shown one after another. This procedure may facilitate learning though the immediate comparison between the two stimuli or stimulus categories. Furthermore, in this study, a new stimulus pair was presented to the sharks on each trial of the categorization experiments. Reaching the learning criterion at the end of the training procedure was therefore already sufficient to show that sharks had successfully learned to distinguish the two basic categories. Accordingly, no additional generalization or transfer tests were necessary to confirm this ability. Transfer tests were only used to elucidate which strategy the sharks had applied in the process. At least when working with sharks, rays and cichlids, it is advantageous to limit the use of unrewarded transfer tests as animals often learn quickly that these kind of tests are unrewarded. This obviously influences the decision-making process and may therefore bias results.
The visual systems of vertebrates have developed along different evolutionary pathways, despite facing many of the same pressures. Like birds, fish (both teleosts and sharks) have to be able to recognize objects from different angles, at different distances or sizes, and as opposed to terrestrial vertebrates not just from the front, the back and the sides but also from above and below. Categorization offers a means for animals to reduce the amount of information they have to retain and respond to by learning to integrate similar features into a class rule (Delius et al. 2000; Huber 2001). This way, animals can respond faster and more appropriately to other organisms, such as predators, prey or conspecifics, and to new situations. It is therefore an essential ability to have for just about any organism living and acting in a complex environment. The present study showed that bamboo sharks discriminate 2D objects according to categories; however, as opposed to cichlids, they seemed to pay attention predominantly to the positive or rewarded stimulus and to a lesser extent to the negative one, as suggested by the transfer test results. In the future, it would be interesting to investigate the types of mechanisms that sharks applied in the process, while there are several theories concerning visual category learning in humans and birds (e.g., Richler and Palmeri 2014) little in this respect has been investigated in fish.
References
Aust U, Huber L (2001) The role of item- and category-specific information in the discrimination of people versus non-people images by pigeons. Anim Learn Behav 29:107–119
Basile BM, Hampton RR (2013) Monkeys show recognition without priming in a classification task. Behav Proc 93:50–61
Berankova J, Vesely P, Sykorova J, Fuchs R (2014) The role of key features in predator recognition by untrained birds. Anim Cogn 17:963–971
Berg ME, Ward MD, Dai ZJ, Arantes J, Grace RC (2014) Comparing performance of humans and pigeons in rule-based visual categorization tasks. Learn Motiv 45:44–58
Bovet D, Vauclair J (1998) Functional categorization of objects and of their pictures in baboons (Papio anubis). Learn Motiv 29:309–322
Castro L, Wasserman EA (2014) Pigeons’ tracking of relevant attributes in categorization learning. J Exp Psychol Anim Learn Cogn 40:195–211
Cook RG, Smith JD (2006) Stages of abstraction and exemplar memorization in pigeon category learning. Psychol Sci 17:1059
D’Amato MR, Van Sant P (1988) The person concept in monkeys (Cebus apella). J Exp Psychol Anim Behav Proc 14:43–56
Delius JD, Jitsumori M, Siemann M (2000) Stimulus equivalencies through discrimination reversals. In: Heyes C, Huber L (eds) The evolution of cognition. MIT Press, Cambridge, MA, pp 103–122
Fuss T, Bleckmann H, Schluessel V (2014a) Visual discrimination abilities in grey bamboo sharks (Chiloscyllium griseum). Zool 117:104–111
Fuss T, Bleckmann H, Schluessel V (2014b) Place learning prior to and after telencephalon ablation in bamboo and coral cat sharks (Chiloscyllium griseum and Atelomycterus marmoratus). J Comp Physiol 200:37–52
Fuss T, Bleckmann H, Schluessel V (2014c) The brain creates illusions not just for us: sharks (Chiloscyllium griseum) can ‘see the magic’ as well. Front Neural Circ 8:24. doi:10.3389/fncir.2014.00024
Guttridge TL, Gruber SH, Gledhill KS, Croft DP, Sims DW, Krause J (2009) Social preferences of juvenile lemon sharks Negaprion brevirostris. Anim Behav 78:543–548
Herrnstein RJ, Loveland DH (1964) Complex visual concept in the pigeon. Science 146:549–551
Huber L (2001) Visual categorization in pigeons. In: Cook RG (ed) Avian visual cognition. www.pigeon.psy.tufts.edu/avc/huber/
Jitsumori M, Matsuzawa T (1991) Picture perception in monkey and pigeons: transfer of right-up versus upside-down discrimination of photographic objects across conceptual categories. Primates 32:473–482
Lazareva OF, Freiburger LK, Wassermann EA (2004) Pigeons concurrently categorize photographs at both basic and superordinate levels. Psychon Bull Rev 11:1111–1117
Lubow R (1974) High-order concept formation in the pigeon. J Exp Anal Behav 21:475–483
Mervis CB, Rosch E (1981) Categorization of natural objects. Ann Rev Psychol 32:89–115
Murai C, Tomonaga M, Kamegai K, Terazawa N, Yamaguchi MK (2004) Do infant Japanese macaques (Macaca fuscata) categorize objects without specific training? Primates 45:1–6
Philips KA (1996) Natural conceptual behavior in squirrel monkeys (Saimiri sciureus): an experimental investigation. Primates 37:327–332
Richler JJ, Palmeri TJ (2014) Visual category learning. WIREs. Cogn Sci 5:75–94
Roberts WA, Mazmanian DS (1988) Concept learning at different levels of abstraction by pigeons, monkeys, and people. J Exp Psychol Anim Behav Proc 14:247–260
Schluessel V (2014) Who would have thought that ‘Jaws’ also has brains? Cognitive functions in elasmobranchs. Anim Cogn. doi:10.1007/s10071-014-0762-z
Schluessel V, Bleckmann H (2012) Spatial learning and memory retention in the grey bamboo shark (Chiloscyllium griseum). Zoology 115:346–353
Schluessel V, Fricke G, Bleckmann H (2012) Visual discrimination and object categorization in the cichlid Pseudotropheus sp. Anim Cogn 15:525–537
Schluessel V, Kraniotakes H, Bleckmann H (2014a) Visual discrimination of rotated 3D objects in Malawi Cichlids (Pseudotropheu sp.): a first indication for form constancy in fishes. Anim Cogn 17:359–371
Schluessel V, Beil O, Weber T, Bleckmann H (2014b) Symmetry perception in sharks (Chiloscyllium griseum) and cichlids (Pseudotropheus sp.). Anim Cogn 17:1187–1205
Smith JD (2014) Prototypes, exemplars, and the natural history of categorization. Psychon Bull Rev 21:312–331
Smith EE, Medin DI (1981) Categories and concepts. Harvard University Press, Cambridge
Smith JD, Minda JP (1998) Prototypes in the mist: the early epochs of category learning. J Exp Psychol Learn Mem Cogn 24:1411–1436
Smith JD, Crossley MJ, Boomer J, Church BA, Beran MJ, Ashby FG (2012a) Implicit and explicit category learning by capuchin monkeys (Cebus apella). J Comp Physiol 126:294–304
Smith JD, Berg ME, Cook RG, Murphy MS, Crossley MJ, Boomer J, Spiering B, Beran MJ, Church BA, Ashby FG, Grace RC (2012b) Implicit and explicit categorization: a tale of four species. Neurosci Biobehav Rev 36:2355–2369
Spinozzi G (1995) Categorization in monkeys and chimpanzees. Behav Brain Res 74:17–24
Spinozzi G (1996) Categorization in monkeys and chimpanzees. Behav Brain Res 74:17–24
Spinozzi G, Lubrano G, Truppa V (2004) Categorization of above and below spatial relations by tufted capuchin monkeys (Cebus apella). J Comp Psychol 118:403–412
Vonk J, MacDonald SE (2002) Natural concepts in a juvenile gorilla (gorilla gorilla gorilla) at three levels of abstraction. J Exp Anal Behav 78:315–332
Yoshikubo S (1985) Species discrimination and concept formation by rhesus monkeys (Macaca mulatta). Primates 26:285–299
Acknowledgments
We would like to thank S. Braun for help with animal caretaking, maintenance and repairs. We are specifically grateful to the ‘Haus des Meeres’ in Vienna for supplying the animals used during this study and to H. Bleckmann for providing laboratory space and support. The research reported herein was performed under the guidelines established by the current German animal protection law. This study was funded by a DFG Grant to V.S. (SCHL1919/4-1). The experimental work presented herein was conducted by Diandra Duengen as part of her Bachelor-Thesis at the University of Bonn.
Conflict of interest
The authors declare that they have no conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Schluessel, V., Duengen, D. Irrespective of size, scales, color or body shape, all fish are just fish: object categorization in the gray bamboo shark Chiloscyllium griseum . Anim Cogn 18, 497–507 (2015). https://doi.org/10.1007/s10071-014-0818-0
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10071-014-0818-0