
Abstract
Comparative research can shed light on the evolutionary roots and cognitive underpinnings of prosocial behavior by revealing not only positive instances of prosocial motivations in other species, but also the boundary conditions of these motivations. To explore factors that may constrain prosocial behavior, we examined whether brown capuchins (Cebus apella), which demonstrate regard for the welfare of conspecifics in other contexts, would behave prosocially in a minimal-cost instrumental helping task. We observed that when given the opportunity to share tokens that allow a conspecific to obtain food from an apparatus, capuchins showed no regard for another individual’s welfare. Subjects transferred tokens to an adjacent chamber when the apparatus was present, but did so just as often when the chamber was empty as when there was a recipient present to obtain food. While capuchins are sensitive to others’ welfare in some contexts, the current results suggest that they do not spontaneously produce goal-specific helping actions on behalf of a conspecific. The lack of regard for others exhibited in this context provides insights into the factors that may constrain prosocial behavior in capuchins and other primate species.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Prosocial behavior presents an evolutionary puzzle. Intuitively, natural selection should favor only those traits that maximize self-interest (Darwin 1871). Behaving prosocially—that is, intentionally acting for the benefit of others (Eisenberg et al. 2006)—appears inconsistent with behaving so as to maximize relative fitness. However, an extensive body of theoretical work in evolutionary biology and beyond has explored how behaviors that are altruistic at the proximate level can nonetheless be evolutionarily stable strategies (Hamilton 1964; Trivers 1971; Axelrod 1984; see Nowak 2006 for recent review). Indeed, observational and experimental research over the past several decades has revealed prosocial behaviors in a variety of animal species (see reviews in Dugatkin 1997; Clutton-Brock 2009).
The emerging body of work on prosocial behavior in the animal kingdom yields yet another puzzle, however. Although numerous species act to benefit others, there exists a surprising degree of variation in the contexts under which they do so. Within nonhuman primates—the taxonomic group most extensively studied to date—we find notable inconsistencies in the extent to which different species show regard for others’ welfare in a given context. Chimpanzees (Pan troglodytes), for example, appeared consistently insensitive to others’ welfare in food-donation tasks in which they had the opportunity to deliver food to another conspecific (Silk et al. 2005; Jensen et al. 2006; Vonk et al. 2008; Yamamoto and Tanaka 2010). Although some species (e.g., cotton-top tamarins, Saguinus oedipus: Cronin et al. 2009; Stevens 2010) have behaved like chimpanzees on such tasks, ignoring the welfare of their conspecific partner, other species (e.g., capuchin monkeys, Cebus apella: de Waal et al. 2008; Lakshminarayanan and Santos 2008; marmosets, Callithrix jacchus: Burkart et al. 2007) have selectively chosen the prosocial option and thus exhibit what appears to be preferences for others’ welfare.
Primates also exhibit within-species variation when tested across different tasks. In contrast to their performance in the experiments described above, chimpanzees show remarkable prosocial motivation in other contexts. It has been observed that chimpanzees help other conspecifics in several real-world situations; they console victims of aggression (de Waal and van Roosmalen 1979; de Waal and Aureli 1996; Aureli and de Waal 2000), share food (de Waal 1989), groom conspecifics (de Waal 1997), and cooperate during hunting activities (e.g., Boesch 2002; Mitani et al. 2000; Muller 2005). Chimpanzees also help others in more controlled experimental tests of instrumental helping. Warneken and colleagues, for example, have shown that chimpanzees will reliably help human experimenters and conspecifics achieve a goal (Warneken and Tomasello 2006; Warneken et al. 2007; Melis et al. 2010). A rather different pattern of experimental performance, however, has been observed in capuchin monkeys—whereas in forced-choice contexts, brown capuchins exhibited greater regard for others than did chimpanzees (de Waal et al. 2008; Lakshminarayanan and Santos 2008), this species exhibited relatively weak prosocial preferences in the only instrumental helping task performed to date in this species (Barnes et al. 2008).
Although this variability in primate prosocial behavior is perplexing, it may provide an opportunity to uncover both the boundary conditions and the psychological mechanisms that underlie prosocial behavior in the animal kingdom, by allowing us to explore how social-cognitive abilities, particularly goal attribution capacities, constrain the expression of prosocial preferences. The prevalence of other-benefiting behavior within a given species or in a certain context may depend on the species’ capacity to infer goals and on the ease with which another’s goal can be inferred from the situation or by overt indications or requests (see discussion in Melis et al. 2010; Jaeggi et al. 2010; Silk 2009).
To explore these factors, Warneken, Melis, and colleagues presented chimpanzees with situations in which they could help a recipient achieve a goal and systematically varied a number of different motivational and cognitive factors (Warneken and Tomasello 2006; Warneken et al. 2007; Melis et al. 2010). They manipulated the salience of an experimenter’s goal, for example, by varying whether she reached and grasped for an out-of-reach object (goal condition) or looked at but did not reach for the object (no-goal condition). In these studies, they also manipulated motivational factors including presence of a reward for helping, the identity of the recipient (conspecific or human experimenter), the energy-expenditure involved, and whether food was present. In these experiments, chimpanzees helped significantly more often when a recipient indicated a goal, but helped consistently regardless of whether the recipient was another chimpanzee or a human experimenter, and did so for food and non-food items, irrespective of whether rewards were present.
One interpretation offered by these authors is that chimpanzees demonstrate prosocial motives in tasks where goals are made salient (Warneken and Tomasello 2006; Warneken et al. 2007; Melis et al. 2010) but not in contexts where these explicit goal cues are lacking (e.g., Silk et al. 2005; Jensen et al. 2006; Vonk et al. 2008), suggesting that social-cognitive demands may constrain the expression of prosocial preferences in nonhumans. In this way, mapping out the task-specific boundaries of a given species’ prosocial tendencies can shed light on cognitive and motivational prerequisites for the emergence of prosociality in a given species.
In the current experiment, we extend this line of work begun by Warneken and colleagues. Instrumental helping tasks can provide an ideal methodology for teasing apart competing hypotheses about the cognitive and motivational prerequisites for prosociality, as one can vary both motivational variables (e.g., the recipient identity and the costs and potential payoffs) and social-cognitive factors (e.g., the simplicity of the instrumental context and the salience of the end goal). To date, however, only chimpanzees have been tested across a broad range of experimental contexts. Here, we extend instrumental helping tasks to an important population for understanding the phylogenetic history of prosocial behavior: capuchin monkeys. Capuchins are an important test species because they show some understanding of goals and intentions (Barnes et al. 2008; Phillips et al. 2009) and show regard for others’ welfare on forced-choice tasks (de Waal et al. 2008; Lakshminarayanan and Santos 2008), yet have exhibited relatively fragile prosocial preferences in instrumental helping tasks performed to date. Using Warneken et al.’s out-of-reach paradigm, Barnes et al. (2008) found that although capuchins would hand an object to a human experimenter, they were highly susceptible to motivational manipulations. Capuchins were very sensitive, for example, to the presence of motivating rewards; when a food reward was available, they handed over an object at ceiling levels regardless of whether the experimenter was reaching for it. Only in the absence of a reward did the capuchins’ response depend on whether an experimenter was indicating a particular goal. This suggests that capuchins possessed some of the requisite social-cognitive skills to help in this task—they were able to infer a goal from the experimenter’s reach—but that the motivation to fulfill that goal was secondary to the subject’s own motivation to obtain a reward. Similarly, capuchins weighted their own costs quite heavily, refusing to help in a situation that demanded more effort on their part (Barnes et al. 2008, Experiment 1). Thus, in contrast to chimpanzees’ robust helping across motivational manipulations, spontaneous helping in capuchins appears very limited by specific motivational concerns.
Here, we expand on this work by identifying three possible constraints on capuchin helping behavior and exploring how the removal of such constraints affects capuchins’ sensitivity to others’ welfare. First, the current task allowed capuchins to help a recipient towards whom they should be especially motivated to behave prosocially—specifically, a familiar in-group conspecific—rather than a human experimenter. Second, in contrast to Barnes et al. (2008), subjects in the current experiment had no opportunity to obtain food themselves by helping (or refusing to help) the recipient. Finally, by first training our subjects to perform a novel goal action themselves and allowing them to observe the action before every trial, we hoped to maximize the ease with which they could recognize the recipient’s goal and identify a means of completion, without introducing reward expectancies or ceiling-level target behaviors.
In our instrumental helping task, capuchin monkeys were allowed to help a conspecific recipient by sharing tokens that the recipient alone could use to obtain food from a “vending machine” apparatus. For token sharing to be construed as instrumental helping—that is, involving a representation of the recipient’s goal and a motivation to complete it—token transfers should be more frequent in this helping context than in control conditions in which either the recipient or the object of the goal (the apparatus) is missing. Sustained transfers in the absence of the apparatus would suggest that subjects are insensitive to the specific goals and potential payoffs of the recipient. Transfers when the recipient is absent would suggest that the presence of the apparatus perhaps increases the salience of the token-transfer action, but that the transfers are not socially motivated.
Methods
Subjects
We tested brown capuchin monkeys. Our subjects live in a socially housed colony at the Comparative Cognition Laboratory at Yale University. They live in a naturalistic social enclosure that is divided into six sections, allowing the capuchins some control over their proximity to other colony members. Capuchins have ad libitum access to water and are fed a diet of monkey chow twice each day as well as treats such as fruit, nuts, and cereal. Our test group consisted of four females and two males (NN, MD, HG, MP, HR, and AG), all subadult or adult. Our recipient individual (JM) was an adult female from the group who was related to two of the individuals in our sample (MD and MP).
Apparatus
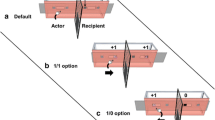
All capuchins used in this study were trained to deposit a specific set of tokens (cylindrical pieces of PVC pipe; 5 cm long, 3 cm diameter, not used in other studies) into a “vending machine” apparatus in order to receive a food reward (single grape, ~2.5 g). The apparatus was constructed from foam core and a PVC pipe and was operated surreptitiously by an experimenter stationed behind the machine. When a monkey dropped a token through a hole in the foam core, the experimenter covertly deposited a grape into the PVC pipe. The apparatus was attached (during conditions for which it was present) to the right experimental chamber (see Fig. 1).

Experimental apparatus and setup (goal condition). In the goal condition, the subject was provided with 5 PVC tokens and a recipient was placed in the adjacent chamber with the vending apparatus attached. The setup for the no-goal condition was identical to the goal condition except that the apparatus was removed, and the no-recipient condition was identical except that the recipient was removed
Procedure
Training
Both the subjects and the recipient (JM) first underwent three training phases in order to learn the nature of the tokens and vending machine apparatus, followed by a series of testing phases.
In the first training phase (training phase 1), each individual learned to operate the vending machine apparatus using the tokens. During this phase, the monkey was enclosed in the right experimental chamber with the apparatus attached to this chamber. The monkey was then provided with a token and given 5 min of exposure to the apparatus. If the monkey deposited the plastic token in the correct opening, he or she received a grape through the apparatus and was given another token to use. If the individual deposited the token 10 times and received 10 grapes during the 5-min session, the session was considered a success. Several monkeys succeeded immediately, but others did not understand the task on the first session. These monkeys were allowed to remain in the enclosure while another monkey completed the session and were then subsequently retested on training phase 1. All monkeys successfully deposited the token during their next session. Monkeys were considered to have passed this apparatus training stage upon completing three consecutive successful sessions.
In the second training phase (training phase 2), subjects were exposed to the existence of two openings between the adjacent chambers and the possibility of receiving items, specifically tokens, through these openings. Each individual was enclosed in the right experimental chamber, while a human experimenter stood in the left chamber. The experimenter provided the subject with the plastic token by passing it through an opening between the chambers so that the capuchin could then deposit the token into the machine. Individuals were considered to have passed this training stage upon completing a single session of receiving and depositing 10 tokens in 5 min.
In the third training phase (training phase 3), we established that capuchins understood that there were openings between the two chambers through which a token could pass when tested from the enclosure on the opposite side. Each subject was enclosed in the left experimental chamber, while a human experimenter stood in the right chamber. In this training stage, the human experimenter offered a grape in exchange for a token. Importantly, the token used in this training phase was not the same plastic token that operated the vending machine; instead, we used another object (a familiar metal trading token), which our subjects had learned previously could be exchanged with humans for food (e.g., Chen et al. 2006; Lakshminarayanan et al. 2008; Lakshminarayanan et al. in press). In this way, we were able to teach the capuchins how to pass items through the opening between the chambers without reinforcing them for the vending machine-specific token. We considered subjects to have passed this training phase when they completed a single session of passing the metal token through the opening ten times, in exchange for ten grapes.
Testing
Once all six subjects (HR, MP, HG, AG, NN, and MD) and the selected recipient (JM), completed these three training phases, they were run through a series of experimental trials. Every subject underwent three sessions (10 min each) in each of three conditions: a goal condition, a no-recipient condition, and a no-goal condition, counterbalanced for order. Subject monkeys were brought into the left experimental chamber, while the recipient and/or apparatus (depending on condition) were in the right experimental chamber. The two chambers were separated by a single wire-mesh partition, which contained two small (4.5 × 4.5 cm) square openings. The token used to operate the apparatus fit smoothly through these openings.
An apparatus reminder session was run immediately before each of the experimental conditions, during which the recipient was given three tokens and used them to get three grapes from the vending machine. This session was included so that the subject would have a chance to observe the recipient successfully using tokens to get grapes from the vending machine. To avoid negative reactions based on inequity at this stage, the subject also received three grapes that were handed by a human experimenter. The subject was given these three grapes before the recipient received her three tokens to prevent the subject from learning spurious contingencies between the recipient using tokens and the subject getting grapes.
In the primary test condition, the goal condition, the subject in the left chamber was given five tokens but did not have access to the apparatus, whereas the recipient had access to the apparatus but no tokens. In other words, the subject had the opportunity to help the recipient by passing her up to five tokens. If the subject passed a token, the recipient could deposit the token to obtain a grape. Experimenters recorded online how many tokens the subject transferred to the adjacent enclosure.
Frequency of token transfer in this goal condition was compared to transfer in two control conditions. In one control condition, the no-goal condition, the recipient was still present, but did not have access to the vending machine. The no-goal condition was in all other ways identical to the goal condition. The purpose of the no-goal condition was to rule out the possibility that token transfers were merely due to preferences for pushing tokens through holes or passing items to a conspecific. If transfers were motivated by the welfare of the conspecific partner, subjects would pass the tokens less often in this condition (when the recipient had no access to the vending machine) compared to the goal condition (when the vending machine was present for the recipient to use).
In the second control condition, the no-recipient condition, the vending machine was attached but there was no recipient present to operate it. The no-recipient condition was in all other ways identical to the goal condition. The purpose of the no-recipient condition was to rule out the possibility that the presence of the apparatus made the tokens more salient for the subject or somehow highlighted the token-passing action, irrespective of any benefit it might provide to the partner. If transfers were motivated by prosocial concerns, subjects would pass the tokens less often in this condition (when the recipient was not present to use the vending machine) than in the goal condition (when the recipient was present to use the vending machine).
All sessions were coded online by recording the number of tokens that had been transferred at the end of each 10-min experimental trial. Each trial was also recorded on a Sony video camera for documentation purposes.Footnote 1
Results
As shown in Fig. 2, the mean number of tokens transferred differed across conditions. Although subjects rarely transferred tokens in the no-goal condition (Mean = 0.53 transfers, SE = 0.35), we saw higher rates of transfer in both the goal condition (Mean = 2.00, SE = 0.54) and the no-recipient condition (Mean = 2.33, SE = 0.53). We analyzed the total number of tokens transferred across the three conditions using a repeated measures ANOVA that had condition (goal condition, no-goal condition, and no-recipient condition) and session number (first, second, and third) as factors. Mauchly’s test indicated that the assumption of sphericity had been satisfied (Mauchly’s W2=0.662, P = 0.538), justifying the use of parametric analyses. One individual in our initial sample did not perform any transfers throughout the entirety of the experiment and was therefore not included in the analyses. However, the effects remain comparable with this individual included. There was no effect of session number (F 2,8 = 0.04, P = 0.96) and no interactions across session and condition (F 4,16 = 2.19, P = 0.12), but we did find a significant effect of condition (F 2,8 = 8.51, P = 0.01).

Mean (+SE) number of token transfers across conditions
To follow up on this significant main effect, we used the Fisher’s LSD (uncorrected) pairwise comparison method to compare token transfers in each of our three conditions. These post hoc tests (see Fig. 2) revealed that the rate of transfers during the no-goal condition differed significantly from the no-recipient condition (Mean difference = 1.80, P = 0.009). The rate of transfers was also higher in the goal condition than in the no-goal condition, though this difference was not significant two-tailed (Mean difference =1.47, P = 0.066). Importantly, there was no significant difference between the goal and no-recipient conditions (Mean difference = 0.33, P = 0.460). Although there was high between-subjects variability, with some individuals passing at overall higher rates than others, no individuals demonstrated a helpful pattern of behavior.
Discussion
Given that capuchins have shown regard for conspecifics’ welfare in various food-donation tasks (de Waal et al. 2008; Lakshminarayanan and Santos 2008), but more limited regard in instrumental helping tasks (Barnes et al. 2008), the current experiment was designed to shed light on the specific contours of prosociality in this species. In this experiment, subjects transferred more tokens to an adjacent chamber when a token-receiving apparatus was present in that chamber (goal condition and no-recipient condition) than when it was absent (no-goal condition). Crucially, however, when the apparatus was attached, capuchins did not differentiate between conditions when a recipient was present to use the tokens and when there was no recipient. Thus, in contrast to previous studies (de Waal et al. 2008; Lakshminarayanan and Santos 2008), we find no evidence of a prosocial preference in this task.
Importantly, the obtained pattern of token transfers suggests that the capuchins do comprehend the physical setup and are willing and able to perform the relevant behavior. Subjects shared tokens during some trials, and not at ceiling levels, suggesting that helpful tendencies would be detected by the experimental measure. Because the transfer rate depended on the presence or absence of the apparatus, we conclude that our subjects were indeed attending to the adjacent enclosure and moderating their token transfers accordingly. If subjects were oblivious to the conditions in the recipient chamber, or did not understand the relationship between the tokens and the apparatus, we would expect there to have been no systematic differences between these conditions. Instead, we find a significant difference between the no-recipient and no-goal conditions, and a difference between the goal and no-goal conditions, though not significant two-tailed (P = 0.066). Furthermore, capuchins have been shown to successfully transfer tools in a very similar experimental setup that involved tool exchange rather than prosocial sharing (Westergaard et al. 2007). It seems, therefore, that failure to help cannot be explained by failure to understand the task.
As described above, we took into account several cognitive and motivational factors in designing this experiment, such that it should have elicited helpful behaviors if instrumental helping were indeed part of this species’ behavioral repertoire. This experimental setup allowed for helping directed toward an in-group conspecific and provided subjects with no opportunity to gain for themselves. Furthermore, subjects in this experiment were personally familiar with the specific goal of their conspecific partner, having been previously trained on the same task. Nonetheless, factors remain that could have limited the expression of prosocial preferences. Given that the outcome of a helpful action was a reward to the recipient when the subject received nothing, prosociality could potentially have been constrained by inequity aversion. In a recent study pitting prosocial motives against fairness concerns, Brosnan et al. (2010) found that large inequities reduced prosocial tendencies in capuchins. However, inequity aversion would predict a low rate of token transfer in the goal condition. Instead, we see that subjects do transfer tokens frequently when the apparatus is present, they simply fail to differentiate based on the presence of a recipient.
Given that this experiment was intended as a rather liberal test of capuchin instrumental helping, failure to help in this task not only provides insights into the limits of capuchin prosociality but also sheds interpretive light on previous work in this area. One plausible explanation for relatively frequent token transfers in both conditions where the apparatus was present is that the apparatus merely highlighted apparatus-related objects or actions for subjects. The sustained token transfers observed in the presence of the apparatus (regardless of the presence of a recipient) raise important questions about the role of goal cues in this and previous helping studies. In this experiment, conditions in which the apparatus was present function similarly to the “reach” conditions in previous work on instrumental helping (Warneken and Tomasello 2006; Warneken et al. 2007; Barnes et al. 2008). As described above, the presence of explicit goal cues like reaches or vocal solicitations have been construed as social-cognitive manipulations that affect whether an actor shows regard for another individual. An alternative interpretation is that such cues make certain end states or action patterns salient, without eliciting any particular inferences about how the completion of that action benefits another individual.
The fact that token transfers depended on the presence of the apparatus but not the presence of a recipient lends credence to this action-salience explanation. Future work on instrumental helping will be necessary to determine whether cues like reaching for objects affect subjects’ behavior by conveying another individual’s desire or by highlighting a particular action that the subject might then choose to perform independently of the benefit it provides to another. Researchers can gain traction on this question by designing experiments that dissociate cues that highlight certain actions from cues that indicate potential benefits to another individual. The dual controls included in this experiment (i.e., a control in which a goal state is made salient but the recipient absent and a control in which the recipient is present but without a goal) should be considered in future work in this area.
The current results constitute a step toward a more complete picture of the boundary conditions for prosocial behavior in this species. Given converging evidence for capuchin prosocial preferences in other tasks, it seems unlikely that failure to help in our experiment can be attributed to total disregard for others. Because this experimental design eliminated many of the previously posited constraints on prosocial behavior, failure to help in this task requires further explanation. One possible interpretation of our result is that capuchins can represent goals and are motivated by the welfare of familiar others, but are unable to integrate relevant representations in the way that is required for successful helping. Instrumental helping involves representing the physical context and representing another individual’s goal state and employing these representations in service of a prosocial motivation. Previous work has demonstrated that capuchins can comprehend intentions and goal-directed actions (Barnes et al. 2008; Phillips et al. 2009) and prefer options that benefit partners (de Waal et al. 2008; Lakshminarayanan and Santos 2008). In this task, however, capuchins do not appear to exploit action understanding to generate positive outcomes for their conspecific partner.
If goals, constraints, and actions are represented independently from representations of potential benefits to social partners, the results reported here might reflect a failure to integrate these representations. This interpretation aligns with existing evidence that primates are able to exploit certain mental state representations more readily in competitive than cooperative contexts (Hare et al. 2000, 2001; Flombaum and Santos 2005) and would provide a plausible constraint on the contexts in which capuchins behave prosocially. It may be that a crucial difference between the prosocial behaviors observed in humans and those found in more distantly related species lies in the fact that humans show regard for others in more flexible and diverse contexts because they are able to make use of relevant social-cognitive representations in a way that certain other species are not. Future research will certainly be necessary to unpack the exact constraints that prevent instrumental helping in this experiment and to distinguish between this account and other possible interpretations of our data.
Regardless of whether this particular interpretation holds, the current results reveal the importance of future work on specific forms of prosociality such as instrumental helping. Disentangling how existing paradigms differentially recruit goal and action representations and prosocial motivations will be an important project for the field. We observed no evidence of instrumental helping in a species that is capable of representing others’ goals (Barnes et al. 2008) and providing benefits to others in the absence of social cues (de Waal et al. 2008; Lakshminarayanan and Santos 2008). We therefore suggest that although capuchins are motivated by the welfare of others and possess the capacity to represent intentions and goal-directed actions, they do not necessarily integrate these to produce spontaneously helpful actions. More importantly, this result adds to a growing picture of animal prosociality and emphasizes the need for additional theoretical and empirical work on factors that may constrain prosocial behavior, creating the puzzling inconsistences observed across species and contexts.
Notes
Due to a camera hard drive error, eleven of these video recordings were unusable. These video files were distributed evenly across conditions, and performance on these trials did not differ systematically from performance on the remaining trials.
References
Aureli F, de Waal FBM (2000) Natural conflict resolution. University of California Press, Berkeley, CA
Axelrod R (1984) The Evolution of Cooperation. Basic Books, New York
Barnes JL, Martinez M, Langer M, Hill T, Santos LR (2008) Helping behavior and regard for others in capuchin monkeys (Cebus apella): An evolutionary perspective on altruism. Biol Lett 4(6):638–640
Boesch C (2002) Cooperative hunting roles among Taï chimpanzees. Hum Nat 13(1):27–46
Brosnan S, Houser D, Leimgruber K, Xiaoe E, Chenc T, de Waal FBM (2010) Competing demands of prosociality and equity in monkeys. Evol Hum Behav 31:279–288
Burkart J, Fehr E, Efferson C, van Schaik CP (2007) Other-regarding preferences in a non-human primate: common marmosets provision food altruistically. Proc Natl Acad Sci 104(50):19762–19766
Chen MK, Lakshminaryanan V, Santos LR (2006) The evolution of our preferences: evidence from capuchin monkey trading behavior. J Pol Econ 114(3):517–537
Clutton-Brock T (2009) Cooperation between non-kin in animal societies. Nature. 462:51–57
Cronin KA, Schroeder KKE, Rothwell ES, Silk JB, Snowdon CT (2009) Cooperatively breeding cottontop tamarins (Saguinus oedipus) do not donate rewards to their long-term mates. J Comp Psychol 123:231–241
Darwin C (1871) The descent of man and selection in relation to sex. John Murray, London
de Waal FBM (1989) Food sharing and reciprocal obligations among chimpanzees. J of Hum Evol 18(5):433–459
de Waal FBM (1997) The Chimpanzee’s service economy: Food for grooming. Evol Hum Behav 18(6):375–386
de Waal FBM, Aureli F (1996) Consolation, reconciliation, and a possible cognitive difference between macaque and chimpanzee. In: Russon AE, Bard KA, Parker ST (eds) Reaching into Thought: The Minds of the Great Apes. Cambridge University Press, Cambridge, UK, pp 80–110
de Waal FBM, van Roosmalen A (1979) Reconciliation and consolation among chimpanzees. Behav Ecol Sociobiol 5:55–66
de Waal F, Leimgruber K, Greenberg A (2008) Giving is self-rewarding for monkeys. Proc Nat Acad Sci 105:13685–13689
Dugatkin LA (1997) Cooperation among animals. Oxford University Press, New York
Eisenberg N, Fabes RA, Spinrad TL (2006) Prosocial development. In: Eisenberg N (ed) Handbook of child psychology: Social, emotional and personality development Vol 3. Wiley, Hoboken, NJ, pp 646–718
Flombaum JI, Santos LR (2005) Rhesus monkeys attribute perception to others. Curr Biol 15:447–452
Hamilton WD (1964) The genetical evolution of social behaviour. J Theor Biol 7:1–52
Hare B, Call J, Agnetta B, Tomasello M (2000) Chimpanzees know what conspecifics do and do not see. Anim Behav 59:771–785
Hare B, Call J, Tomasello M (2001) Do chimpanzees know what conspecifics know? Anim Behav 61:139–151
Jaeggi AV, Burkart JM, Van Schaik CP (2010) On the psychology of cooperation in humans and other primates: combining the natural history and experimental evidence of prosociality. Phil Trans R Soc B 365:2723–2735
Jensen K, Hare B, Call J, Tomasello M (2006) What’s in it for me? Self-regard precludes altruism and spite in chimpanzees. Proc R Soc Lond B273:1013–1021
Lakshminarayanan VR, Santos LR (2008) Capuchin monkeys are sensitive to others’ welfare. Curr Biol 18:R999–R1000
Lakshminarayanan V, Chen MK, Santos LR (2008) Endowment effect in capuchin monkeys (Cebus apella). Phil Trans Royal Soc B: Biol Sci 363:3837–3844
Lakshminarayanan V, Santos LR, Chen MK (in press) The evolution of decision-making under risk: framing effects in monkey risk preferences. J Exp Soc Psychol. doi:10.1016/j.jesp.2010.12.011
Melis AP, Warneken F, Jensen K, Schneider AC, Call J, Tomasello M (2010) Chimpanzees help conspecifics obtain food and non-food items. Proc Biol Sci. doi:10.1098/rspb.2010.1735
Mitani JC, Merriwether DA, Zhang C (2000) Male affiliation, cooperation and kinship in wild chimpanzees. Anim Behav 59(4):885–893
Muller MN (2005) Conflict and cooperation in wild chimpanzees. Adv Study Behav 35:275–331
Nowak M (2006) Five rules for the evolution of cooperation. Science 314:1560–1563
Phillips W, Barnes JL, Mahajan N, Yamaguchi M, Santos LR (2009) ‘Unwilling’ versus unable’: Capuchins’ (Cebus apella) Understanding of human intentional action? Dev Sci 12(6):938–945
Silk JB (2009) Social preferences in primates. In: Glimcker P, Camerer C, Fehr E, Poldrack R (eds) Neuroeconomics: decision making and the brain. Elsevier, London, pp 269–283
Silk JB, Brosnan SF, Vonk J, Henrich J, Povinelli DJ, Richardson AS et al (2005) Chimpanzees are indifferent to the welfare of unrelated group members. Nature 437:1357–1359
Stevens J (2010) Donor payoffs and other-regarding preferences in cotton-top tamarins (Saguinus oedipus). Anim Cog. doi:10.1007/s10071-010-0309-x
Trivers R (1971) The evolution of reciprocal altruism. Q Rev Biol 46(1):35–57
Vonk J, Brosnan SF, Silk JB, Henrich J, Richardson AS, Lambeth SP et al (2008) Chimpanzees do not take advantage of very low cost opportunities to deliver food to unrelated group members. Anim Behav 75:1757–1770
Warneken F, Tomasello M (2006) Altruistic helping in human infants and young chimpanzees. Science 311:1301–1303
Warneken F, Hare B, Melis AP, Hanus D, Tomasello M (2007) Spontaneous altruism by chimpanzees and young children. PLoS Biol 5:e184
Westergaard GC, Evans TA, Howell S (2007) Token mediated tool exchange between tufted capuchin monkeys (Cebus apella). Anim Cog 10:407–414
Yamamoto S, Tanaka M (2010) The influence of kin relationship and reciprocal context on chimpanzees’ other-regarding preferences. Anim Behav 79(3):595–602
Acknowledgments
The authors would like to thank Jordan Comins, Elizabeth Spelke, Sally Boysen, and two anonymous reviewers for helpful comments on an earlier draft of the manuscript. We also thank Tanya Mayer, Mary Altonje, Camille Buchanan, Cora Mukerji, and Caroline Drucker for their help running these studies, and the members of the Comparative Cognition Laboratory at Yale University for valuable discussions about the experimental design. We thank the McDonnell Foundation Scholar Award and Yale University for their help funding this work. This research was conducted in compliance with federal laws of the United States of America and with the regulations of Yale University. The authors declare that they have no conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Skerry, A.E., Sheskin, M. & Santos, L.R. Capuchin monkeys are not prosocial in an instrumental helping task. Anim Cogn 14, 647–654 (2011). https://doi.org/10.1007/s10071-011-0399-0
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10071-011-0399-0