
Abstract
Visual co-orientation with another’s gaze direction (gaze following) may provide important information about the location of food, social interactions or predators. Gaze following has been shown in a variety of mammals, but only in few bird species, and has not been tested in precocial birds at all. It has been suggested that gaze following is an anti-predator behaviour, and in Common ravens (Corvus corax) and rooks (C. frugilegus), it emerges shortly after fledging, at a time when young birds leave the predator-safe nest. However, if gaze following is adaptive, the developmental pattern should differ between altricial and precocial birds. Greylag geese (Anser anser) are highly social birds with a precocial development. Goslings move and feed independently within 24 h post-hatching, and they are highly vulnerable to aerial predators. We therefore predicted that greylag geese are capable of gaze following and that they develop this skill already pre-fledging. We experimentally tested 19 hand-raised greylag goslings for their ability to follow a conspecific’s gaze when they were between 10 days and 6 weeks old. In line with our predictions, first responses were already detectable in 10-day-old goslings. Our results therefore not only demonstrate that greylag geese follow the gaze of conspecifics into distant space, but that they also develop this ability much earlier than altricial birds.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The gaze direction (defined as head-and-eye orientation; Emery 2000) of an individual can convey important information about the environment and about the individual’s interests and goals (Baron-Cohen 1995). The ability to detect (e.g. Burger et al. 1991; Carter et al. 2008; Hampton 1994; Ristau 1991; von Bayern and Emery 2009) or to follow the gaze direction of another individual to a position in space (“gaze following”, Emery 2000) therefore may be highly beneficial, as it may alert individuals to the presence of predators, conspecifics, food or other resources (Povinelli and Eddy 1996; Tomasello et al. 1998). Two different modes of gaze following have been proposed (Gomez 2005): individuals may follow gaze (1) into distant space, i.e. they visually co-orient to a position above, behind or to the side of another individual (e.g. Kaminski et al. 2005) or (2) geometrically around a barrier, i.e. they re-position themselves if a barrier prevents them from seeing what another individual is looking at (e.g. Bugnyar et al. 2004; Tomasello et al. 1999).
Two cognitive models have been suggested to underlie the different modes of gaze following. The low-level model suggests gaze following to be an automatic visual orientation reflex according to which subjects align their gaze direction with others and search along this line of sight until they find something of interest. This model is assumed to be at work in gaze following into distant space (Povinelli and Eddy 1996). The high-level model associated with geometrical gaze following assumes an understanding for other individuals’ perception (Povinelli and Eddy 1996), although an alternative hypothesis suggests that a low-level mechanism and experience with barriers may be sufficient to achieve geometrical gaze following (Tomasello et al. 1999).
The two modes of gaze following may not only differ in their cognitive pre-requisites. Bugnyar et al. (2004) and Schloegl et al. (2007) argued that the two modes also may serve different functions. This interpretation was based on the observation that in Common ravens (Corvus corax), the two modes followed different developmental trajectories and that the onset of each mode corresponded with important developmental changes in the life of the birds. Gaze following into distant space emerged shortly after fledging, when birds increased their mobility and became increasingly vulnerable to predators. Indeed, mortality in ravens is highest in the first month post fledging (Bugnyar, unpubl. data). Therefore, gaze following into distant space may serve primarily as an anti-predator behaviour. In contrast, geometrical gaze following was not detectable until several months later, when ravens became completely independent from their parents. At this age, ravens improve their cache protection strategies and begin to use barriers as a visual screen during caching events (Bugnyar et al. 2007). Consequently, geometrical gaze following may be of functional importance mainly when animals employ barriers as visual shields, e.g. to cache food or socially interact outside the view of others. In line with this idea, similar developmental patterns of gaze following were found in another food-caching corvid, the rook (Corvus frugilegus; Schloegl et al. 2008).
Beside corvids, the ontogeny of gaze following into distant space has only been studied in primates, and again the developmental patterns correspond roughly with important changes in life, i.e. with the end of infancy (Ferrari et al. 2000; Tomasello et al. 2001). Further, chimpanzees (Pan troglodytes) follow gaze towards objects within their visual field before objects outside their visual field (Okamoto et al. 2002), and the first occurrence of gaze following in barbary macaques (Macaca sylvanus) occurred when the adult’s gaze was accompanied by facial expressions (Teufel et al. 2010).
Our understanding of the ontogeny of gaze following remains fragmentary not only because of the limited number of species investigated but also because all show an altricial development of young. In birds, altricial young remain stationary in the nest until fledging and are entirely dependent on food provisioning through their parents. In contrast, precocial birds are mobile within a few hours after hatching, follow their parents away from the nest and feed independently. As such, they are vulnerable to aerial and ground predators much earlier than altricial birds. This leads to very different ontogenetic trajectories, e.g. precocial, but not altricial birds open their eyes within hours post-hatching (Starck and Ricklefs 1998), whereas altricial birds develop larger brains than precocial ones (e.g. Iwaniuk and Hurd 2005). In consequence, the development of gaze following in corvids may not be fully representative for birds in general.
We therefore investigated the ontogeny of gaze following into distant space in the precocial, highly social greylag geese (Anser anser). From summer until late winter, greylag geese form large flocks, presumably to reduce the predation risk (e.g. Kotrschal et al. 2006; Rutschke 1982). Pairs are long-term monogamous and show bi-parental care for up to seven goslings, which leave the nest 24 h after hatching (Lorenz 1988). Geese are highly vulnerable to predation and are regularly preyed upon by ground as well as aerial predators (Hemetsberger 2002; Kotrschal et al. 1992; Lorenz 1988). Goslings are particularly vulnerable, as no escape flight is possible before fledging. This, in turn, should make gaze following into distant space highly advantageous, as it may allow individuals to faster detect and thus escape predators. If gaze following developed in adjustment to ecological needs (Bugnyar et al. 2004), we would predict that gaze following into distant space, as an anti-predator behaviour (1) should be present also in the highly social, precocial geese, and (2) should develop at an earlier life history stage than in altricial birds, i.e. shortly after hatching and pre-fledging.
Methods
Animals
A non-migratory flock of greylag geese was introduced into the valley of the River Alm in Upper Austria by Konrad Lorenz in 1973 (Lorenz 1988). The geese are unrestrained and roam the valley between the Konrad Lorenz Research Station (KLF) and a lake 10 km to the south, where they roost at night. During daytime, they can be found at the meadows and ponds close to the KLF, where they are provided with supplemental food twice a day year-round to keep the flock accessible. Birds are individually marked with coloured leg bands and their life histories have been monitored continuously since the establishment of the flock. In recent years, flock size varied between 130 and 150 individuals. As in other populations, natural predation, mainly by red foxes (Vulpes vulpes) and occasionally golden eagles (Aquila chrysaetus), is common and may account for losing up to 10% of the adult flock per year (Hemetsberger 2001). About 25% of the flock are hand-raised following a well-established hand-raising tradition (Hemetsberger et al. 2010). Hand-raised geese are fully integrated into the flock and they establish social bonds with goose partners and raise offspring indistinguishable from goose-raised geese (Hemetsberger et al. 2010).
The 19 focal individuals of this study hatched within one week in early April 2008 and were hand-raised in three sibling groups of six, six and seven individuals (Table 1). Eggs originated from five different nests (A–E, Table 1), but we considered all individuals raised by one human foster parent as siblings regardless of genetic relatedness (Frigerio et al. 2003). Goslings were individually marked with coloured leg bands within 3 h after hatching. To minimize the risk of infection with parasites and subsequent early mortality, the three sibling groups did not have contact with the rest of the flock during their first 3 weeks of life. Thereafter, they were moved to the same area where most geese also raise their goslings (“OGB”, Lorenz 1988), and from then on, their daily spatio-temporal activity patterns followed those of goose families. All focal individuals survived until fledging at an age of 8–10 weeks and fully integrated into the flock.
Experimental set-up and procedure
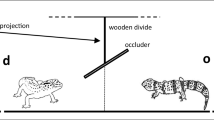
We tested goslings for their ability to follow a sibling’s look-up when subjects were ten to 43 days old, following well-established test-procedures (e.g. Loretto et al. 2010; Schloegl et al. 2008). Experiments were conducted in an arena with two compartments, separated by a wire-mesh partition with an opaque screen 0.4 m and later 0.6 m high (Fig. 1). The size of the arena was adjusted to the size/age of the goslings and initially measured 0.5 m × 1.0 m in the first raising location (“PB”) and 1.0 m × 1.5 m in the second location (“OGB”). Due to local conditions and logistic constraints, all four arena walls were opaque in PB, while one length of the arena in OGB was composed of wire mesh and, therefore, allowed visual contact between tested birds and their siblings. This change in the arena design was necessary to allow filming the experiments and to maintain visual contact between all goslings and the foster parent also in the bigger arena in OGB.

Experimental set-up to test greylag goslings’ ability to follow a sibling’s gaze into distant space. The wire-mesh part of the partition is drawn by a dotted line, the opaque part of the partition by a solid black line
Goslings were habituated to the set-up prior to the start of the experiments, yet separation from the human foster parent and their siblings frequently caused stress responses. Therefore, experiments were conducted by the human foster parent of the respective sibling group. The human foster parent positioned herself close to and in view of the tested goslings to keep the subjects’ social stress as low as possible. For the same reason, the goslings not partaking in an experimental session remained with their foster parent and thus were audible and, in OGB, visible to their siblings in the arena. However, the goslings outside the arena were not able to see the gaze cue given to the model bird in the arena.
Experimental sessions consisted of three conditions per focal individual. For each focal, the three conditions were conducted in direct succession and in randomized order, with a minimum interval of 30 s between conditions. In the test condition, the focal bird (i.e. the observer) and one of its siblings (i.e. the model) were positioned in the two adjacent compartments of the arena. The experimenter (E) projected a laser point onto the opaque partition on the side of the model until the model looked up to the laser point. We scored if the observer looked up within 5 s after the model’s look-up. If the observer looked up after the elapsed 5 s, we did not consider this as gaze following and thus did not score the look-up. Control1 (C1) was conducted to test whether the focal bird could perceive the laser point presented on the model’s side. For this purpose, E projected a laser point onto the model’s side of the opaque partition for 5 s, but no model was present. We scored if the observer looked up during the laser point projection or, as in the test, within 5 s after the projection of the laser point. During Control2 (C2), both the model and the observer were present, but no laser point was projected. From a 2-min video, we chose at random three 20-s sequences in which the model bird did not look up. We discarded the first 10 s of each sequence to exclude that observers may have responded to model look-ups occurring prior to the onset of the sequence and scored whether or not the observer looked up during the last 10 s of this sequence (Loretto et al. 2010). The total duration of both controls thus was 10 s and as such a more conservative measure for the comparison with the test condition.
The position (left/right of the partition) of the subjects was randomized across conditions and sessions. Experiments were conducted daily, whereby each gosling was tested as the observer every third day, and no gosling was observer and model on the same experimental day. The first twelve experimental days were conducted in PB. After moving to OGB, the subjects had an experimental break of one day to habituate to the new surroundings. After twelve experimental days in OGB, we interrupted the experiments for one week to motivate the subjects again.
All sessions were videotaped with a Sony Handycam DCR-HC23E for later analysis. We conducted a total of 167 trials per condition. Due to motivational problems, the number of trials per subject varied, but each focal individual participated in at least seven trials per condition. Several trials were later discarded due to technical problems or no look-up of the model in the test condition. This resulted in a final number of 128 test trials, 157 C1 trials and 160 C2 trials, with a range of three to ten trials per individual and condition (Table 1).
Data analysis
As birds commonly move eyes and head simultaneously, we used head orientation and head movement to ascertain a look-up. Bird eyes possess two foveas; they primarily use the lateral fovea in monocular vision (i.e. turning the head and looking with one eye) to look at targets in the distance and the frontal fovea in binocular vision to look at targets in close proximity (Dawkins 2002). Thus, birds usually use monocular looking to look at objects in the sky and consequently, the determining factors to detect look-ups were the inclination of the head compared to the neck, the position of the beak (lateral or horizontal) and the turning of the head for looking with one eye only (Bugnyar et al. 2004; Dawkins 2002). To assess the subject’s receptiveness for the gaze cues, we also determined the duration of lying, standing, walking and pacing around as well as the number of distress calls emitted by the observer during the first 20 s of each trial. “Pacing around” was defined as walking briskly back and forth along the arena walls with the head alternately close to the ground or stretched high, indicative of the intention to leave the arena. In contrast, “walking” constituted slower, undirected movement with the head in a relaxed position. The duration of model look-ups was measured from the videotapes to the nearest 0.1 s. All trials were scored from videotapes by S.K., and a second person (B.M.W.) scored a randomly selected 14% of the trials. To be able to analyse both the observer’s and the model’s behaviour, both compartments of the test arena were visible on the videotape, but the scorers could not see the laser point and were blind to if, when and on which side of the barrier the laser point was projected. The two human scorers showed an excellent inter-observer reliability of 97% (Cohen’s κ = 0.92).
We conducted generalized linear mixed models (GLMMs) using the GenStat 12.1 statistical package, applying the restricted maximum likelihood (REML) procedure for repeated sampling with an unbalanced design (GenStat 2007). All GLMMs were constructed with the binomial variable “observer look-up” as the response variate and identity of the model and identity of the observer as random terms to account for repeated measurements (445 trials in 19 individuals). The first GLMM was comprised of the data of all three conditions (N = 445 trials) and contained condition (test, C1 or C2), location (PB or OGB), age, sex of the observer, sibling group, nest of origin, the number of distress calls (observer), the number of distress calls (model) and the durations of the observer’s lying down, standing, walking and pacing around as fixed terms. Another GLMM was constructed with data from the test condition only (N = 128 trials) to assess which fixed terms had an influence on the gaze following behaviour of the subjects. Similarly, one GLMM was constructed with data from C1 and C2 (N = 317 trials) only to determine whether there were differences between the two control conditions.
According to standard stepwise model reduction procedures, we sequentially deleted fixed terms in order of decreasing significance, whereby the least significant term was determined after each removal step (Galwey 2006; Garamszegi et al. 2009; Garamszegi 2011). Deletion of fixed terms continued until only terms with a significance value below 0.1 remained. This was then considered the final model. Excluded terms were re-entered one by one into the final model to confirm that they did not explain a significant part of the variation. However, terms were only regarded as being significant if P < 0.05. We present Wald statistics, because the change in deviance when dropping a term from the model approximates a χ2-distribution (Galwey 2006).
Results
The experimental condition significantly influenced the occurrence of look-ups in greylag goslings (Fig. 2). Goslings looked up in 37% of the test trials, but only in 13% (C1) and 10% (C2) of the control trials, respectively (Table 1). Notably, 50% of the goslings looked up in response to their siblings’ gaze cues already in their first test trial at an age of 10–12 days, while only 13% of the goslings looked up in either of the control trials (Fig. 3).

Percentage (mean ± SE) of look-ups during the three experimental conditions in the two test locations PB and OGB

Percentage of individuals looking up during the test (black circle), C1 (white triangle) and C2 (grey triangle) conditions over the course of the experiment. Experimental days represent a time frame of three days, during which each individual was tested once. The first experimental day corresponds to an age of 10–12 days
Additionally, across all conditions, look-ups occurred more frequently in the first test period in PB (23%) than in OGB (15%, Fig. 2), while age per se did not have a significant influence on look-up behaviour. Also, individuals generally looked up less the more they walked or paced around the arena. Neither sex, sibling group, nest of origin nor any of the other investigated behaviours influenced the occurrence of look-ups (Table 2).
Similar to look-ups in general, gaze following in the test condition occurred more frequently in PB (47% of test trials) than in OGB (32% of test trials). Towards the end of the study, the look-up rates increased again, but not significantly so. Gaze following was influenced by the observer’s locomotory state: individuals that paced around more responded less to their siblings’ gaze demonstrations (Table 2). Also in the test condition, age did not have a significant influence on look-up behaviour.
The occurrence of look-ups did not differ between the two control conditions and also decreased during controls the more an individual paced around. However, unlike gaze following, look-ups during controls were not influenced by the location but instead tended to decrease with increasing age (Table 2, Fig. 3).
Also the model’s responsiveness to the laser point changed over the course of the experiments. During the first ten days of testing, models always responded to the laser point cue. The first test trial without a model response to the stimulus was observed on the 11th experimental day and responsiveness steadily decreased thereafter (GLMM, Wald = 26.57, df = 1, P < 0.001). The duration of the models’ gaze cues ranged from 0.7 to 6.0 s (mean ± SD = 2.2 ± 2 s) and also decreased over the experimental period (GLMM, Wald = 5.16, df = 1, P = 0.025).
Discussion
To our knowledge, this is the first experimental demonstration that a precocial bird species visually follows the gaze of conspecifics. This adds to the increasing evidence that gaze following into distant space is a basic cognitive skill common in vertebrates and may be found in reptiles (Wilkinson et al. 2010), mammals (e.g. Kaminski et al. 2005), altricial (Loretto et al. 2010; Schloegl et al. 2008) and precocial birds (this study). Additionally, our study demonstrates that goslings follow other goslings’ gaze already at the earliest tested age of 10 days and that the propensity to follow other’s gaze does not increase thereafter.
The very early gaze following in geese is the earliest known onset of gaze following to date. Two-day-old bobtail quails (Colinus virginianus) responded to a human’s gaze direction by avoiding the area the human was looking at (Jaime et al. 2009), but it remains to be tested whether quail also show gaze following by aligning their own looking direction with that of a human or conspecific.
Our findings are consistent with the prediction that gaze following into distant space may have evolved primarily as a predator avoidance strategy (Bugnyar et al. 2004) and should, therefore, be present in precocial birds earlier than in altricial birds. Gaze following was found in greylag goslings roughly 6 weeks before fledging. In contrast, ravens and rooks do not follow gaze until at least one week post-fledging (Schloegl et al. 2008). However, not only gaze following emerges earlier in geese but they also start to look up by using their monocular field already in the first days after hatching (S. K., personal observation). The first instances of monocular looking in ravens were not observed before they reached 30 days of age (Schloegl et al. 2007), and the first look-up in rooks was observed in a 10- to 14-day-old individual (Schloegl et al. 2008). The first instances of gaze following in geese therefore occurred at an age at which the corvid species only started to open their eyes. These findings make biological sense, as goslings move independently 24 h post-hatching and are subjected to a high predation pressure, whereas under the dense canopy of raven nests, looking up and scanning for aerial predators does not become relevant until fledging. Future tests may elucidate whether geese are also capable of the cognitively more advanced geometrical gaze following (Bugnyar et al. 2004; Loretto et al. 2010) and if so, whether the development of this skill follows comparable developmental trajectories as in corvids.
Juvenile barbary macaques steadily increase their gaze following rates until they are one year old (Teufel et al. 2010), but this does not seem the case in geese. This suggests that geese do not increase their gaze following responses or increases occur outside the time frame of this study. On the contrary, the goslings showed weaker gaze following responses at the second location (OGB) than at the first location (PB); thus, older goslings may be less prone to show gaze following. However, our analysis did not support an effect of age, and ontogenetic changes may have only a minor influence on the observed decrease in look-up behaviour upon change of location. Rather, the goslings may have learned to interpret the model’s behaviour more accurately or they may have been less distracted during the experiments in PB. In OGB, but not in PB, those siblings that did not participate in the experiments were in the observer’s visual field and may have reduced the observer’s attention towards the model. Alternatively, the gaze cue of the model may have been devalued for the observer because none of the other siblings present performed look-ups. Another, mutually not exclusive possibility is the occurrence of habituation to the test situation, which is a well-documented phenomenon in tests of gaze following in monkeys and ravens (Schloegl et al. 2007; Tomasello et al. 2001). The observers could have learned that the gaze cues of the model birds were essentially directed at nothing and ceased to respond. Indeed, during the test condition, we sometimes noticed that the observers stopped moving just after the model reacted to the stimulus, looked in the direction of its sibling, but did not follow its gaze (personal observation). This indicates that they noticed their sibling’s look-up, but may have been habituated to the gaze cues already and thus did not follow the gaze. Aside from the observers, also the model birds habituated to the laser point, which is in concordance with the known rapid habituation to predator models in birds (Schleidt 1961; Schleidt et al. 1983).
The present study is the fourth study using this method for the investigation into gaze following to conspecific models in non-human animals (see also Loretto et al. 2010; Schloegl et al. 2008; Wilkinson et al. 2010). These studies required a human experimenter standing in the proximity of the subjects to provide a gaze cue for the model. Thereby, the humans may have provided involuntary cues (“Clever Hans”-effect), or observers may have looked up to see the experimenter rather than in response to the model. We are confident, however, that this was not the case. First, in C2, the experimenter did not need to project a laser point and consequently was standing further away from the arena than in C1 and the test condition. However, the observers’ rates of look-ups were identical in C1 and C2, but were distinctively different from the test condition. Second, in C1 and in the test condition, the experimenter was positioned in a way to optimally project the laser point onto the barrier. By doing so, the barrier partially blocked the observer’s view to the experimenter, making the perception and response to behavioural cues less likely. Third, if observers were inclined to look at the experimenter, we would assume that this occurred particularly in those cases in which the experimenter was the only social partner in sight (i.e., in condition C1 conducted in location PB), but again this was not the case. Furthermore, in the majority of trials, the goslings looked up laterally and adjusted their looking direction to the behaviour of the models: during the test, 43 of 47 look-ups were directed towards the barrier or directly opposite. Models thus looked to where the sibling looked or moved the head into the same direction as the model’s head, again supporting our interpretation of the goslings’ behaviour as gaze following. In contrast, only 4 of 47 look-ups were directed towards the experimenter, and look-up direction during controls was random. Finally, each sibling group was tested by their respective foster parent. It is likely that potential involuntary cues would differ between the different experimenters, but look-up behaviour did not differ between sibling groups. Taken together, gaze following in response to the model’s gaze cues remains the most parsimonious explanation for the look-up behaviour of the observers.
The gaze following rate of 37% in this study was relatively low compared to mammalian and other bird studies, in which response rates of 60–100% were observed (Kaminski et al. 2005; Loretto et al. 2010; Schloegl et al. 2007; Tomasello et al. 1999). Several not mutually exclusive explanations are possible. First, as outlined earlier, the presence of the subjects’ siblings in the vicinity of the test arena may have distracted the observers. Similarly, the low rates of gaze following may have been a consequence of the separation of the subjects from their siblings and of being restrained from approaching their siblings and foster parents during the experiments. Although the human foster parents remained in the goslings’ visual field, models and observers showed frequent indications of stress. This is supported by our finding that the more the observers were pacing around along the boundaries of the arena, the less they were looking up and the less they followed their siblings’ gaze. Thus, the subjects apparently were less attentive to the experiments, whereas individuals that calmly stood in the arena presumably paid more attention to their surroundings. Furthermore, in big family groups, as found in greylag geese, not every individual may need to be vigilant constantly (Pulliam 1973; Roberts 1995). Greylag goslings grow up in family groups consisting of two parents and a mean number of five to seven siblings (Hemetsberger et al. 2010). Adult geese frequently give alarm calls upon detection of potential danger (Lorenz 1988), and goslings thus may not need to follow the gaze of a family member each time a look-up occurs. Also, we used goslings as models, but adults, and in particular the parents, may be more potent cue-givers than other goslings. In ravens, gaze following occurred in response to both, conspecific and human models (Schloegl et al. 2008), suggesting that both are adequate models, but to our knowledge, influences of the model’s age have never been investigated. Notably, we also observed a large variation in response rates to a sibling’s look-up cues, with some individuals responding in more than 80% of the trials and others not at all. This could indicate that different individuals contribute differently to anti-predator surveillance, which would be worth to investigate in future studies.
In conclusion, this study showed that greylag goslings followed the gaze of conspecifics into distant space and that this behaviour developed shortly after hatching and well before fledging. The early emergence of this behaviour supports the hypothesis that gaze following may have evolved for predator avoidance and demonstrates the commonality of gaze following in socially living vertebrates.
References
Baron-Cohen S (1995) Mindblindness: an essay on autism and theory of mind. MIT Press, Cambridge, Massachusetts
Bugnyar T, Stöwe M, Heinrich B (2004) Ravens, Corvus corax, follow gaze direction of humans around obstacles. Proc R Soc B 271:1331–1336
Bugnyar T, Stöwe M, Heinrich B (2007) The ontogeny of caching in ravens, Corvus corax. Anim Behav 74:757–767. doi:10.1016j.anbehav.2006.08.019
Burger J, Gochfeld M, Murray BG Jr (1991) Role of a predator’s eye size in risk perception by basking black iguana, Ctenosaura similis. Anim Behav 42:471–476
Carter J, Lyons NJ, Cole HL, Goldsmith AR (2008) Subtle cues of predation risk: starlings respond to a predator’s direction of eye-gaze. Proc R Soc B 275:1709–1715. doi:10.1098/rspb.2008.0095
Dawkins MS (2002) What are birds looking at? Head movements and eye use in chickens. Anim Behav 63:991–998
Emery NJ (2000) The eyes have it: the neuroethology, function and evolution of social gaze. Neurosci Biobehav Rev 24:581–604
Ferrari PF, Kohler E, Fogassi L, Gallese V (2000) The ability to follow eye gaze and its emergence during development in macaque monkeys. Proc Natl Acad Sci USA 97:13997–14002
Frigerio D, Weiß BM, Dittami J, Kotrschal K (2003) Social allies modulate corticosterone excretion and increase success in agonistic interactions in juvenile hand-raised geese (Anser anser). Can J Zool 81:1746–1754
Galwey NW (2006) Introduction to mixed modelling. Wiley, Chichester, UK
Garamszegi LZ (2011) Information-theoretic approaches to statistical analysis in behavioural ecology: an introduction. Behav Ecol Sociobiol 65:1–11
Garamszegi LZ, Calhim S, Dochtermann N, Hegyi G, Hurd PL, Jorgensen C, Kutsukake N, Lajeunesse MJ, Pollard KA, Schielzeth H, Symonds MRE, Nakagawa S (2009) Changing philosophies and tools for statistical inferences in behavioral ecology. Behav Ecol 20:1363–1375
Gomez J-C (2005) Species comparative studies and cognitive development. Trends Cog Sci 9:118–125
Hampton RR (1994) Sensitivity to information specifying the line of gaze of humans in sparrows (Passer domesticus). Behaviour 130:41–51
Hemetsberger J (2001) Die Entwicklung der Grünauer Graugansschar seit 1973. In: Kotrschal K, Müller G, Winkler H (eds) Konzepte der Verhaltensforschung. Konrad Lorenz und die Folgen. Filander, Fürth, Germany, pp 249–260
Hemetsberger J (2002) Populationsbiologische Aspekte der Grünauer Graugansschar (Anser anser). Dissertation, University of Vienna
Hemetsberger J, Scheiber IBR, Weiß BM, Frigerio D, Kotrschal K (2010) Influence of socially envolved hand-raising on life history and stress responses in greylag geese (Anser anser). Interact Stud 11(3):380–395
Iwaniuk AN, Hurd PL (2005) The evolution of cerebrotypes in birds. Brain Behav Evol 65:215–230
Jaime M, Lopez JP, Lickliter R (2009) Bobwhite quail (Colinus virginianus) hatchlings track the direction of human gaze. Anim Cog 12:559–565. doi:10.1007/s10071-009-0214-3
Kaminski J, Riedel J, Call J, Tomasello M (2005) Domestic goats, Capra hircus, follow gaze direction and use social cues in an object choice task. Anim Behav 69:11–18
Kotrschal K, Hemetsberger J, Dittami J (1992) Vigilance in a flock of semi-tame Greylag Geese (Anser anser) in response to approaching eagles Haliaeetus albicilla and Aquila chrysaetos. Wildfowl 43:215–219
Kotrschal K, Hemetsberger J, Weiß BM (2006) Homosociality in greylag geese. In: Vasey PL, Sommer V (eds) Homosexual behaviour in animals: an evolutionary perspective. Cambridge University Press, Cambridge, UK
Lorenz K (1988) Hier bin ich—Wo bist du? Die Ethologie der Graugans. Piper, München, Germany
Loretto M-C, Schloegl C, Bugnyar T (2010) Northern bald ibises follow others’ gaze into distant space but not behind barriers. Biol Lett 6:14–17. doi:10.1098/rsbl.2009.0510
Okamoto S, Tomonaga M, Ishii K, Kawai N, Tanaka M, Matsuzawa T (2002) An infant chimpanzee (Pan troglodytes) follows human gaze. Anim Cog 5:107–114
Povinelli DJ, Eddy TJ (1996) Chimpanzees: joint visual attention. Psychol Sci 7:129–135
Pulliam HR (1973) On the advantages of flocking. J Theor Biol 38:419–422
Ristau CA (1991) Aspects of the cognitive ethology of an Injury-Feigning bird, the piping plover. In: Ristau CA (ed) Cognitive ethology: the minds of other animals. Lawrence Erlbaum Associates, Hillsdale, NJ, pp 79–89
Roberts G (1995) A real-time response of vigilance behaviour to changes in group-size. Anim Behav 50:1371–1374
Rutschke E (1982) Stability and dynamics in the social structure of the greylag goose (Anser anser). Aquila 89:39–55
Schleidt WM (1961) Reaktionen von Truthühnern auf fliegende Raubvögel und Versuche zur Analyse ihrer AAM’s. Z Tierpsych 18:534–560
Schleidt WM, Shalter MD, Carawan TC (1983) The effect of spatial context on habituation to a predator model. Z Tierpsych 61:67–70
Schloegl C, Kotrschal K, Bugnyar T (2007) Gaze following in Common Ravens (Corvus corax): ontogeny and habituation. Anim Behav 74:769–778. doi:10.1016/j.anbehav.2006.08.017
Schloegl C, Schmidt J, Scheid C, Kotrschal K, Bugnyar T (2008) Gaze following in non-human animals: the corvid example. In: Columbus F (ed) Animal behaviour: new research. Nova Science Publishers, New York, pp 73–92
Starck JM, Ricklefs RE (1998) Avian growth and development: evolution within the altricial-precocial spectrum. Oxford University Press, Oxford
Teufel C, Gutmann A, Pirow R, Fischer J (2010) Facial expressions modulate the ontogenetic trajectory of gaze-following among monkeys. Dev Sci 13:913–922
Tomasello M, Call J, Hare B (1998) Five primate species follow the visual gaze of conspecifics. Anim Behav 55:1063–1069
Tomasello M, Hare B, Agnetta B (1999) Chimpanzees, Pan troglodytes, follow gaze direction geometrically. Anim Behav 58:769–777
Tomasello M, Hare B, Fogleman T (2001) The ontogeny of gaze following in chimpanzees (Pan troglodytes) and rhesus macaques (Macaca mulatta). Anim Behav 61:335–343
von Bayern AMP, Emery NJ (2009) Jackdaws respond to human attentional states and communicative cues in different contexts. Curr Biol 19:602–606. doi:10.1016/j.cub.2009.02.062
Wilkinson A, Mandl I, Bugnyar T, Huber L (2010) Gaze following in the red-footed tortoise (Geochelone carbonaria). Anim Cogn 13:765–769. doi:10.1007/s10071-010-0320-2
Acknowledgments
We acknowledge the Herzog von Cumberland Stiftung for cooperation and the “Verein der Förderer der Konrad Lorenz Forschungsstelle” for permanent support. The project was funded by FWF project P20538-B17. We thank Kurt Kotrschal and Thomas Bugnyar for helpful discussions on the topic. Special thanks to Sara Sonja Schimming and Noriko Kondo for hand raising and collecting data and Markus and Christine Platzer for allowing us to raise goslings on their property. The conducted experiments comply with current Austrian law.
Conflict of interest
The authors declare that they have no conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Kehmeier, S., Schloegl, C., Scheiber, I.B.R. et al. Early development of gaze following into distant space in juvenile Greylag geese (Anser anser). Anim Cogn 14, 477–485 (2011). https://doi.org/10.1007/s10071-011-0381-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10071-011-0381-x