
Abstract
In this study we examined how social learning of feeding preferences by zebra finches was affected by the identity of different demonstrators. We presented adult zebra finches with two demonstrators, one male and one female, that exhibited different food choices, and we recorded their subsequent preference when given a choice between the two food types. Previously it was found that young zebra finches' patterns of social learning are affected by the sex of the individual demonstrating a feeding behaviour. This result could be explained by the lack of exposure these animals had to the opposite sex, or by their mating status. Therefore, we investigated the social learning preferences of adult mated zebra finches. We found the same pattern of directed social learning of a different type of feeding behaviour (food colour): female zebra finches preferred the colour of food eaten by male demonstrators, whereas male zebra finches showed little evidence of any preference for the colour of food eaten by female demonstrators. Furthermore, we found that female observers' preferences were biased by demonstrators' relative feeding activity: the female demonstrator was only ever preferred if it ate less than its male counterpart.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
There is increasing evidence that social learning might be an important mechanism through which individuals obtain adaptive information about their environment, in a wide range of species (Avital and Jablonka 2000), allowing individuals to obtain locally adaptive information more quickly than if they had relied on other sources of information (Boyd and Richerson 1985). To assess these ideas, it is necessary to understand the paths by which information spreads through a group. Many previous theoretical studies of cultural evolution have assumed random transmission of behaviour through a population (see Cavalli-Sforza and Feldman 1981; Boyd and Richerson 1985). However, there is observational (Coussi-Korbel and Fragaszy 1995) and experimental (e.g. Nicol and Pope 1999; Swaney et al. 2001) evidence that such transmission is not random in animal populations but is, in fact, biased by social relationships and preferences.
Non-random transmission of socially learned information might arise because the quality of information possessed by different individuals may vary. For example, a conformist preference—a preference for learning the behaviour of the majority of demonstrators—is often found (e.g. Lachlan et al. 1998). Other studies have found that familiar (e.g. Swaney et al. 2001) or dominant (Nicol and Pope 1999) demonstrators are preferred. In all of these cases, we might predict that the information used by familiar individuals or the majority might be more reliable; however, the actual adaptive value of these preferences has not been measured. The most in-depth study of adaptive social learning biases in animals was carried out by Dolman et al. (1996). They found that preferences for conspecific versus heterospecific demonstrators was linked to the social organisation of zenaida dove (Zenaida auriata) populations: populations in which conspecifics foraged together preferred conspecifics demonstrators.
An alternative explanation for biases in social learning preferences is that they are side effects of the different levels of attention individuals pay to different stimuli. In this scenario, social learning preferences might even be maladaptive if attention is attracted for another reason. For example, Coussi-Korbel and Fragaszy (1995) review primatological studies that have found that adult individuals are often less likely to acquire useful information if it is displayed only by younger group members.
The sex of demonstrators is another categorisation that appears to influence social learning by animals. Previous experiments have produced varying results: Nicol and Pope (1999) found that domestic hens avoided learning food preferences from roosters. Benskin et al. (2003) found that female zebra finches preferred learning from male demonstrators. Both adaptive and epiphenomenal hypotheses can be developed about why individuals might prefer learning from the same or different sex. If sexes have different nutritional needs (e.g. minerals for the development of bird eggs), then it might pay to learn from one's own sex. Alternatively, a preference for the opposite sex might be a method for females to assess the foraging ability of potential mates—by sampling the same food, females may learn how good a forager a male is. However, attention may well be directed towards the opposite sex simply as a result of sexual attraction, and a bias for learning from the opposite sex may result without it serving any adaptive function.
In this study we report the results of experiments investigating the feeding preferences of zebra finches, and how they are affected by the sex of demonstrators. Zebra finches are a granivorous Australian estrildid finch species. Their food resources are patchily located, in both time and space (Zann 1996)—factors that might make social learning a particularly adaptive strategy. Although many aspects of their behaviour have been studied in the laboratory, there have been relatively few studies of their feeding behaviour. Initial studies suggested that the presence of conspecifics reduced the rate of learning about food. Beauchamp and Kacelnik (1991) placed pairs of zebra finches in a cage with a food source that was presented in conjunction with a light. They found that if one of the finches had previously learned about the relationship between light and food, this inhibited the other from learning the relationship. Similarly, Coleman and Mellgren (1997) found that putting a foraging partner in a cage next to a zebra finch inhibited learning about a novel food source. However, they also demonstrated social learning: after one of the pair was removed, the other was more likely to choose the food type that the first had been feeding on.
This article reports a follow-up experiment to that carried out by Benskin et al. (2003). They found that domesticated zebra finches did indeed socially learn feeding preferences. Moreover, they found that females preferred the choices of males, whereas males did not show a preference when presented with male and female demonstrators. Such a sex-dependent learning bias could be due to females simply paying more attention to males than to other females, as a result of sexual attraction, and therefore as a side effect, also paying more attention to males' feeding behaviour. Since females rather than males choose sexual partners in this species (Zann 1996), this might explain why Benskin et al. found that only females showed a bias for learning from the opposite sex. However, Benskin et al.'s study was carried out on unmated, young individuals that had had little exposure to the opposite sex prior to the experiment, and therefore an alternative explanation for the results is that females may have simply paid more attention to novel stimuli. Moreover, the lack of preference by males could reflect that they had not reached breeding condition and so were not interested in females. We were interested in how this behavioural preference might have changed in older individuals that were already mated. The primary aim of this experiment was therefore to examine whether demonstrator preferences in mated, mature pairs of zebra finches were the same as those in unmated, young adult zebra finches.
Methods
Twenty-four mated pairs of domesticated wild-type finches were used. Prior to the experiment, they had been housed in pairs for at least 6 months in a breeding colony and had raised at least one clutch successfully. They had been kept in visual but not acoustic isolation from all other pairs, experienced a 14 h:10 h light:dark cycle under fluorescent lighting, and were provided with food ad libitum. During each experimental trial, one pair of zebra finches first observed a second pair feeding on two different food types; they were subsequently given a choice themselves between the two food types.
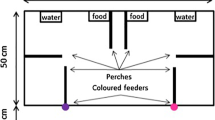
The experiments were carried out in a separate experimental room. Figure 1 shows the experimental set-up. In each trial there were two demonstrators: a male and female from the same mated pair, and two observers from a second mated pair. At any one time, three trials were carried out in parallel, so there were 12 individuals in the experimental room, although individuals were only in visual contact with those in their own trial. Having served as observers in one trial, each pair of zebra finches were used as demonstrators on the following day in the subsequent trial. Once individuals had served as demonstrators, they were returned to the stock room.

Schematic diagram of the experimental set-up. The black and open circles represent the locations of the red and green feeders—the demonstrators are only able to feed at one of these feeders
Normal food was removed from the demonstrators' cage and replaced with two feeders, one containing red and the other green seed (coloured using food colouring), by 12 noon on the day preceding the trial. At 5 p.m. that day, this food was also removed, as well as any spilled seed on the cage floor. At the same time, the observer pair was also moved into the experimental room: their cage was placed opposite that of the demonstrators (Fig. 1), and their food also removed. The aim of this was to ensure that the motivation to feed of observers was equal, since previous work has found that this can affect the tendency to learn socially (Dorrance and Zentall 2001). Also at this time, the demonstrators' cage was split into two equal-sized compartments by inserting an opaque separator that insured that the two demonstrators were visually isolated from one another during the trial. One hour before the beginning of the trial, the observers' cage was also divided by a separator. The trials began between 10.30 and 11.30 a.m. Thus, during the trials, each observer could only see the two demonstrators, and each demonstrator could only see the two observers.
Each trial began with two 15-min observational periods (each followed by a 30-min pause) in which the demonstrators were provided with coloured food, in the presence of the observers. Each demonstrator was provided with two feeders, one containing red food, and the other green. However, one of the feeders was blocked by placing a pellet of paper in its entrance, such that the demonstrator could, in fact, only feed at the other feeder. We arranged the feeders such that one of the demonstrators fed on red food, and the other on green.
Following this was a test phase, in which the two observers (while still separated) were simultaneously provided with two feeders, one containing red seed, and the other containing green seed, to measure their food colour preference. In preliminary trials, the number of pecking movements made towards the seed ("number of pecks") was found to be highly correlated with other possible indications of feeding preferences (time spent feeding, and volume of seed removed from the feeder were also considered). The number of pecks made by both observers towards each colour was recorded, as was the number of pecks made by each of the demonstrators during the observational phases. These records were all taken by one of us (MK). The pecking rate during the trials was low enough to be easily counted accurately. Moreover, during pilot studies involving different individuals, but using identical methods, interobserver reliability checks were carried out, during which identical totals were reached.
The trials were arranged to balance for possible side effects of both demonstrators and observers, as well as to balance for colour preferences for the seed (i.e. 50% of the trials were conducted with the male demonstrator eating red seed, 50% with green seed). Table 1 contains more details of this factorial design.
We did not measure the significance of individual preferences, and we used binomial and non-parametric statistics to examine the existence of feeding preferences in our sample. This was because the numbers of pecks at each feeder were not independent of each other. If an observer pecked at the green feeder, it would be more likely to peck next at the same feeder not just because of any preference it had for the green food, but also because it was already positioned next to the green feeder. Moreover, we considered that the total amount of seed eaten by the observers reflected how hungry they were, rather than the strength of their preference. Therefore, we analysed the proportion of each colour being eaten, rather than simply taking the difference between them.
Results
Results were collected for 23 pairs of individuals (due to the death of 1 individual during the penultimate trial). Table 2 shows the results of the experiment. Most observers ate one food colour much more than the other: 20 female and 18 male observers ate one food type at least twice as much as the other. Similarly, in most observation trials, observers pecked at least 30 times (2 females and 3 males pecked less than 30 times), suggesting that observers had a high motivation to feed. Table 3 summarises the binomial statistical analysis. The only significant effect in the analysis was that female observers pecked more at the food colour eaten by the male demonstrator than that eaten by the female tutor (Fig. 2). This effect remained significant even if the three trials in which the observers did not eat one food colour more than twice as much as the other were excluded from the analysis (two-tailed binomial test, n=20, q=5, P=0.041), or if the two trials in which female observers pecked less than 30 times were excluded (two-tailed binomial test, n=21, q=5, P=0.027). There was no evidence that males preferred pecking at the food colour eaten by the female or male tutors. There was no significant effect for either sex on the observers' preferences of whether the demonstrator was on the left or right side of the cage, or whether it was on the same or opposite side as itself, or for either food colour.

Relation between food choice and the sex of the demonstrator. The y-axis indicates the proportion of pecks made by the observer at the food colour eaten by the female demonstrator (i.e. a score of 0.5 indicates that the observer made an equal number of pecks at both food colours). The x-axis represents the different trials, ordered according to the strength of the preference by the observers. The open bars show the results for the male observers, and the black bars the results for the female observers
We did not find that observers showed any overall preference for the demonstrator that pecked the most or the least (Table 3). However, when we examined how the observers' preferences were affected by the relative amount the male demonstrators ate, compared to how much the females ate, a pattern did emerge (Fig. 3). If female demonstrators ate more than male demonstrators, female observers showed a strong preference for the food eaten by the male demonstrators (Fig. 3a). This correlation was found to be significant (Spearman's rho: 0.42, P<0.03). Only when male demonstrators ate more than the female demonstrators did some female observers eat more of the food eaten by the female observers. Four of the five trials in which female observers chose the female demonstrators, and all three trials in which no strong preference was shown by the observers, occurred when the male demonstrators ate more than the female demonstrators. In these seven trials, the male demonstrators did peck more than during the other trials (mean of 211 pecks during the seven trials; 152 in the others), but a more marked trend was that the female demonstrators ate a lot less during these trials, compared to the others (mean of 86 pecks during the seven trials; 193 in the others). Male observers showed a similar trend to that shown by females (Fig. 3b), but the correlation was not significant (Spearman's rho: 0.18, P>0.1).

Relation between how much the demonstrators ate relative to each other and observer preference. Graphs show on the x-axis how much the female demonstrator ate as a proportion of the total number of pecks made by the demonstrators in that trial (i.e. a score of 0.5 indicates that both demonstrators made the same number of pecks). The y-axis indicates the proportion of pecks made by the observer at the food colour eaten by the female demonstrator (i.e. a score of 0.5 indicates that the observer made an equal number of pecks at both food colours). a Results for the female observers; b results for the male observers
We could not control how much the demonstrators ate during the demonstration phase of the experiment. Therefore it is plausible that the preference of females for male demonstrators was due to an overall difference in how much male and female demonstrators ate. However, the number of pecks made by male and female demonstrators were not noticeably different (mean no. pecks by males: 170; by females: 161. In 12 trials, males pecked more than females; in 11 trials females pecked more than males). Similarly, demonstrators did not show any bias for red or green food. In 10 of the 23 trials, green food was pecked more than red food, and the average number of pecks for the red food was 132, compared to 102 for the green food.
One possible explanation for the difference in preferences between male and female observers is that they differed in their motivation to feed, and this affected their social learning (Dorrance and Zentall 2001). In support of this, we found that in most trials, male observers pecked less than their female counterparts (two-tailed binomial test, n=23, q=5, P=0.01). Although we found no evidence that those male observers that pecked more were more likely to prefer males or females (by recalculating the results after excluding trials in which male observers pecked less than x times, and examining various values of x), we certainly cannot exclude a difference in motivation as an explanation for our results.
Discussion
We found that zebra finches socially learn feeding preferences, in this case the colour of a novel food source, from each other, after two 15-min trials. In contrast to many studies of social learning (e.g. Beauchamp and Kacelnik 1991) in which an individual socially learns from one demonstrator, learning was demonstrated as a preference for one of two food types after both were seen to be eaten at the same time by different demonstrators. This design may be a way to explore how a potentially important decision for social animals is made, since social animals may often be presented with several individuals feeding at the same time from different food sources.
Female zebra finches expressed a preference for the feeding preferences of male demonstrators, whereas males showed no clear preference. This finding was very similar to that found by Benskin et al. (2003), who studied young (3–4 months old), unpaired zebra finches. In their experiment, individuals had been housed in single-sex flocks prior to the experiments, so the preference of females could have resulted from the attractiveness of a novel stimulus (males). Our result suggests that this is not the case, and that the preference of females does not change with mating status. Since the mated pairs were visually separated during our experiments, it could be argued that perhaps the females were behaving as if they were unpaired. However, the subjects were certainly in acoustic contact during the experiment, since in all trials calling interactions between the subjects were observed. As Benskin et al. (2003) argued, a likely explanation for our result is that females paid more attention to the male demonstrator than to the female demonstrator. However, this does not explain why female observers showed a bias and male observers did not. A priori, one might expect at least as strong an opposite-sex bias in males as in females. One possibility is that if female zebra finches are more discriminating in mate choice, then maybe they also devote more time to observing the opposite sex. A more speculative explanation is that by following their food choice, females are able to learn about the foraging prowess of potential mates (in a similar way to courtship feeding), information that might be particularly relevant in a species with biparental care. The lack of selectivity by males may have other explanations, however. Dolman et al. (1996) found that the tendency of doves to learn from conspecifics was related to whether they aggressively defended territories or not. Individuals from populations that defended territories tended not to learn from conspecifics. Male zebra finches are much more aggressive than female zebra finches (Zann 1996). It could be that increased competition over feeding sites between males leads to them paying less attention to the feeding habits of conspecifics in general.
Our second finding was that demonstrator behaviour did appear to affect the feeding choice of female observers. Female observers were more likely to choose the female as a demonstrator if the female demonstrator did not eat as much as her male counterpart. It is important to note that this result cannot explain our finding that female finches preferred male demonstrators, because we found no evidence that male and female demonstrators ate different amounts of food during the demonstration trials. However, this bias appeared to be caused more by the female demonstrator eating less than the male demonstrator eating more. There are two obvious explanations for this bias. Firstly, the observers might perceive more competition for a food type if more of it is being eaten by a demonstrator. Alternatively, data from a similar pilot experiment suggest that the rate of calling by demonstrators is negatively correlated with the rate of pecking (R.F. Lachlan, unpublished data). If so, female demonstrators that call less could be communicating with observers more, and consequently attracting more attention.
Our results provide more evidence for the non-random pattern of social transmission. In this case, it seems likely that biases are caused by social factors unrelated to the information that is socially learned. We would predict that they might influence the spread of a novel feeding behaviour through a group. Moreover, since the biases are unrelated to the quality of information, they could act to hinder the spread of useful information: an innovation made by a female is less likely to be learned by others than one made by a male.
References
Avital E, Jablonka E (2000) Animal traditions: behavioural inheritance in evolution. Cambridge University Press, Cambridge
Beauchamp G, Kacelnik A (1991) Effects of the knowledge of partners on learning rates in zebra finches (Taeniopygia guttata). Anim Behav 41:247–253
Benskin CMH, Mann NI, Lachlan RF, Slater PJB (2003) Social learning directs feeding in the zebra finch (Taeniopygia guttata). Anim Behav (in press)
Boyd R, Richerson PJ (1985) Culture and the evolutionary process. Chicago University Press, Chicago
Cavalli-Sforza LL, Feldman MW (1981) Cultural transmission and evolution: a quantitative approach. Princeton University Press, Princeton, N.J.
Coleman SL, Mellgren RL (1997) Social enhancement and interference of food finding in zebra finches (Taenopygia guttata). J Comp Psychol 111:242–250
Coussi-Korbel S, Fragaszy DM (1995) On the relation between social dynamics and social learning. Anim Behav 50:1441–1453
Dolman CS, Templeton J, Lefebvre L (1996) Mode of foraging is related to tutor preference in Zenaida aurita. J Comp Psychol 110:45–54
Dorrance BR, Zentall TR (2001) Imitative learning in Japanese quail (Coturnix japonica) depends on the motivational state of the observer quail at the time of observation. J Comp Psychol 115:62–67
Lachlan RF, Crooks L, Laland KN (1998) Who follows whom? Shoaling preferences and social learning of foraging information in guppies. Anim Behav 56:181–190
Nicol CJ, Pope SJ (1999) The effects of demonstrator social status and prior foraging success on social learning in laying hens. Anim Behav 57:163–171
Swaney W, Kendal J, Capon H, Brown C, Laland KN (2001) Familiarity facilitates social learning of foraging behaviour in the guppy. Anim Behav 62:591–598
Zann RA (1996) The zebra finch: a synthesis of field and laboratory studies. Oxford University Press, Oxford
Acknowledgements
RFL would like to acknowledge the help of the students of the second-year ethology course, 2000 and 2001, who helped with pilot studies of this project, and Hans Dudart for help with animal care. Funding was provided by a Marie Curie Fellowship to RFL. This research complied with the laws of the Netherlands and was carried out with the approval of the Leiden University Animal Welfare Commission (UDEC).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Katz, M., Lachlan, R.F. Social learning of food types in zebra finches (Taenopygia guttata) is directed by demonstrator sex and feeding activity. Anim Cogn 6, 11–16 (2003). https://doi.org/10.1007/s10071-003-0158-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10071-003-0158-y