
Abstract
The ruptured anterior cruciate ligament (ACL) does not heal spontaneously. Therefore, the development of new healing techniques employing tissue engineering is vital. One of the aspects related to tissue-engineered artificial ligaments is the type of cell to be used for the artificial ligament. In this study, ligament cells from the ACL and periodontal ligament (PDL) were evaluated. In addition, we prepared highly oriented extracellular matrix (ECM) fiber scaffolds that mimicked the structure of the ligament and examined the cellular responses to these scaffolds. Elastin-A and collagen were used as the ECM proteins. Although the cells from the PDL (PDL fibroblasts [PDLFs]) showed approximately 2.1-fold higher expression of alkaline phosphatase (ALP; marker of osteogenic differentiation) than the ACL cells, the expression of ligament-related genes (for type I collagen, type III collagen, and tenomodulin) did not differ between PDLFs and ACL cells. Furthermore, the cellular responses (expression pattern of ligament-related genes and ALP activity) to the ECM were similar between ACL cells and PDLFs. In particular, elastin-A upregulated ALP and downregulated tenomodulin (TeM; a ligament marker) in ligament cells. In contrast, collagen maintained TeM expression in ligament cells. These results suggest that elastin-A promotes the osteogenic differentiation of ligament cells and that collagen maintains the phenotype of ligament cells.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The anterior cruciate ligament (ACL), which joins the femur to the tibia, is one of the most important ligaments in the knee joint. The ACL is an elastic tissue composed of highly oriented collagen, elastin, and fibroblasts [1–3]. This ligament does not heal itself when ruptured because of its poor vascularization, and surgical reconstruction is the standard treatment in such cases [2, 3]. However, current surgical ACL reconstruction approaches are not ideal. Therefore, the generation of artificial ligaments by tissue engineering is an attractive approach.
One of the issues in the preparation of an artificial ligament for tissue engineering is determining the type of cell to use for the artificial ligament. Although ACL cells may be suitable, other cell sources are typically required. Additionally, the effects of ligament components (elastin and collagen) on ligament cells are not clear. Ligaments join bone to bone; therefore, it is necessary to mimic the interface structure between ligaments and bones.
The periodontal ligament (PDL) is the connective tissue present between the tooth cementum and the alveolar bone. When a patient’s own ACL organization is reduced and extraction is impossible, an extractable PDL is considered as a candidate cell source that can be obtained by tooth extraction. The cellular components of the PDL (PDL fibroblasts: PDLFs) have characteristic properties. For example, PDLFs have the ability to form mineralized tissue and show higher collagen production and alkaline phosphatase (ALP) activity than gingival fibroblasts [4–6].
ACL cells and PDLFs share similar characteristics. For example, both types of cells have spindle-shaped morphology, the ability to differentiate into various cell types such as osteoblasts, and similar surface antigens (positive: cluster of differentiation [CD] 29, CD44, CD73, CD105, and CD166; negative: CD34 and CD45) [7–12]. Therefore, PDLFs could be used as an alternative source for the generation of artificial ligaments. However, the differences between these 2 cell types are unclear. In a previous study [13], we used PDLFs, and demonstrated the interactions between PDLFs and the extracellular matrix (ECM). In this study, ACL cells isolated from the human ACL were used to study these interactions in an environment similar to that of knee joints. The purpose of this study was to examine the differences between ACL cells and PDLFs. Furthermore, we prepared a highly oriented fibrous ECM scaffold that mimicked the interface structure between ligaments and bones and examined the cellular responses between ligament cells and the ECM.
Materials and methods
Preparation of cells and materials
Isolation of ACL cells
Human ACL tissue samples were collected aseptically from adult donors during total knee arthroplasty surgery. All the subjects (n = 11) enrolled in this research gave their informed consent, which was approved by our institutional committee on human research, and the study protocol was found to be acceptable by them.
To prepare the tissue culture, the ACL was washed twice in phosphate-buffered saline (PBS), and the surrounding synovial sheath and fat tissue were carefully removed to obtain the core portion of the ligament. Then, the ACL was cut into small pieces and placed on culture dishes with a minimal amount of culture medium, which consisted of Dulbecco’s modified Eagle medium (DMEM) (Sigma-Aldrich Co., Saint Louis, USA) supplemented with 10 % fetal bovine serum (FBS) and 1 % penicillin/streptomycin (Sigma-Aldrich). After 1 h of incubation at 37 °C in a humidified atmosphere of 5 % CO2 and 95 % air, culture medium was added. The culture medium was exchanged every 3 days. Explant tissues were removed when outgrown cells reached subconfluency, and these cells were referred to as passage 0 cells.
For comparison, we also prepared commercially available human PDLFs (Lonza Walkersville Inc. Walkersville, MD, USA). Subculturing of PDLFs was performed according to the manufacturer’s protocol.
Preparation of insoluble elastin, soluble elastin, and type I collagen
Insoluble elastin was extracted from porcine arteries obtained from a slaughtering center (Matsusaka, Mie, Japan) by using the method reported by Miyamoto et al. [14]. Soluble elastin was then prepared from the insoluble elastin by 0.25 M oxalic acid treatment and fractionated into 3 types of elastin: elastin-A, -B, and -C. Elastin-A [15] was used in this study. Type I atelocollagen derived from porcine arteries was obtained from ECM Laboratory Co. (Mie, Japan).
Preparation of ECM scaffold by electrospinning
An original electrospinning device for preparation of the fibrous ECM scaffold was fabricated in this laboratory, and the ECM scaffolds were prepared using a previously reported method [15]. ECM scaffolds were prepared as follows: 1,300 mg of elastin-A (or 650 mg of collagen) and 40 mg of the cross-linker ethylene glycol di-glysidil ether (EGDE, Wako, Osaka, Japan) were mixed in 5,000 mg of trifluororthanol (Wako). The solution was then placed in a plastic syringe attached to a 21G stainless steel needle (Terumo Corp., Tokyo, Japan). The flow rate of the ECM solution jet from the syringe was 1.2 ml/h and the voltage was 15 kV. The distance between the target electrode and the nozzle was 10 cm. After electrospinning, the obtained ECM scaffold was heated for cross-linking at 121 °C for 20 min.
Cell culture
Cell proliferation assay
The proliferative ability of ACL cells was examined according to methods described elsewhere with minor modifications [7]. Briefly, ACL cells were seeded at 5,000 cells/cm2 on a 35-mm cell culture dish (Sumitomo Bakelite Co. Ltd., Tokyo, Japan) in DMEM containing 10 % FBS. The cells were incubated at 37 °C in a humidified atmosphere of 5 % CO2 and 95 % air and serially passaged every 5 days until senescence occurred. The culture medium was exchanged every 3 days. The cells were counted and replated at the same cellular density. Population doubling (PD) for each passage was calculated according to previously reported methods [7]. PD was determined as follows:
where N c is the number of cells after 5 days and N 0 is the initial cell number.
Expression of ligament cell markers of ACL tissue, ACL cells and PDLFs
Ligament-related gene expression was assayed by reverse transcription-polymerase chain reaction (RT-PCR) as described elsewhere [16] for tenomodulin (TeM), a marker for ligaments and tendons [17, 18]; types I and III collagen, which are the major components of the ligament [1]; and ALP, which is a marker for osteoblasts. The oligonucleotide primer sequences for PCR analysis were as follows: glyceraldehyde-3-phosphate dehydrogenase (GAPDH), 5′-GAT GTC ATC ATA TTT GGC AGG TT-3′ and 5′-CCT GCA CCA CCA ACT GCT TAG CCC-3′; TeM, 5′-CCC AGC AGA AAA GCC TAT TG-3′ and 5′-GCG TGA CGG GTC TTC TCT AC-3′; type I collagen, 5′-TTT GTG GAC CTC CGG CTC-3′ and 5′-AAG CAG AGC ACT CGC CCT-3′; type III collagen, 5′-AAC GGT CTC AGT GGA GAA CG-3′ and 5′-CCA CTC TTG AGT TCA GGA TGG-3′; and ALP, 5′-TGG AGC TTC AGA AGC TCA ACA CCA-3′ and 5′-ATC TCG TTG TCT GAG TAC CAG TCC-3′. GAPDH was amplified for 27 cycles, TeM for 34 cycles, types I and III collagen for 28 cycles, and ALP for 32 cycles. Agarose gel electrophoresis (3 %) was performed with one-fifth of the PCR product and analyzed by densitometry using the FAS Ш system (Toyobo Co. Ltd. Tokyo, Japan). Density values for PCR products were normalized to the values for GAPDH.
To perform RT-PCR, ACL tissue that was attached to bone tissue was divided into 2 regions: the ACL region and the bone region. Total RNA from each region was then extracted using RNA iso Plus (TAKARA BIO Inc. Shiga, Japan) according to the manufacturer’s protocol.
In addition, ACL cells were seeded on a 35-mm cell culture dish at 5,000 cells/cm2 in DMEM containing 10 % FBS. Total RNA was extracted when ACL cells reached subconfluency, and then ligament-related gene expression was analyzed. This experiment was performed on 3 independent cultures by using ACL cells, which had been isolated from 3 patients.
Cellular response to the ECM scaffold
ACL cells and PDLFs between passages 4 and 6 were used for seeding onto the ECM scaffold. The ECM scaffold (2 cm × 2.5 cm × 80 μm) was sterilized at 121 °C for 30 min and placed on a 35-mm cell culture dish. The cells were seeded onto the ECM scaffold at a density of 5 × 104 cells/scaffold and incubated at 37 °C for 1 h. After 1 h, DMEM containing 10 % FBS was added to the culture dish, and the cells were incubated for 1 week. The culture medium was exchanged every 3 days.
Assessment methods
Immunohistofluorescence using frozen sections
To evaluate the components of the ACL, immunofluorescence staining was performed using frozen sections. After removal of extra tissue from ACL samples, they were cut to a suitable size. Then, they were embedded in OCT compound (Sakura Finetek Japan Co. Ltd., Tokyo, Japan), frozen, and sectioned using a microtome (7-μm-thick sections).
Frozen sections were fixed in cold acetone:ethanol (1:1) for 10 min. After washing, the sections were blocked with 1 % bovine serum blocking solution for 30 min and then incubated at 4 °C overnight with a 1:200 dilution of primary antibody (mouse anti-human collagen I [AbD Serotec, Kidlington, UK], collagen III antibody (rabbit) [GeneTex, Inc., San Antonio, TX], and polyclonal antibody to human tropoelastin [Elastin Products Company, Inc., Owensville, Missouri, USA]). The sections were then incubated at room temperature for 90 min with a 1:200 dilution of the secondary antibody (DyLight 649-labeled antibody to rabbit IgG [Kirkegaard and Perry Laboratories, Inc., Gaithersburg, MD] and DyLight 649-labeled antibody to mouse IgG [Kirkegaard and Perry Laboratories, Inc.]). Fluorescent-stained sections were observed using a confocal laser microscope (Fluoview FV 100; Olympus, Tokyo, Japan). The observation conditions were standardized for each protein to evaluate the strength of protein expression. Green auto-fluorescence of the ECM scaffolds was detected, which allowed the properties of the ECM scaffolds to be directly observed.
Scanning electron microscopy
The ECM scaffolds were sputter-coated with Au for 120 s using an ion-sputterer (E-1010; Hitachi, Tokyo, Japan). The Au-coated samples were then examined using a field-emission scanning electron microscope (S-4000; Hitachi) at an acceleration voltage of 20 kV.
Alkaline phosphatase assay
Protein extracts of ligament cells were prepared using 0.1 % Triton X-100/20 mM HEPES–NaOH. The ALP assay was performed using LabAssay ALP (Wako) according to the manufacturer’s protocol. The data obtained were normalized against the amount of protein measured by the bicinchoninic acid method. When the cells on the ECM scaffold were examined, an ECM scaffold without seeded cells was prepared as a negative control.
Data analysis
All data have been expressed as mean value ± standard deviation (SD) values. For the analysis of covariance, the SAS software for statistical analysis (StatView version 5.0; SAS Institute Inc., USA) was used. Dunnett’s test and the Tukey–Kramer test were used. A value of p < 0.05 indicated statistical significance.
Results
Gene expression of ACL tissue and bone tissue
The gene expression in ACL tissue and bone tissue was analyzed by RT-PCR (Fig. 1). Although the expression of type I collagen was similar in ACL and bone tissue, the expression of type III collagen and TeM was 2 and 4 times higher, respectively, in ACL tissue than in bone tissue. In contrast, ALP expression was 5.4 times higher in bone tissue than in ACL tissue.


Characteristics of ACL cells: gene expression in the case of human ACL tissue and bone tissue. a Type I collagen mRNA expression. b Type III collagen mRNA expression. c TeM mRNA expression. d ALP mRNA expression. Fresh tissues were used. The data were normalized to those for GAPDH. ACL tissue, n = 4; bone tissue, n = 3. Characteristics of cells outgrown from the ACL. e Cumulative population doubling of cells outgrown from the ACL. f Type I collagen mRNA expression. g Type III collagen mRNA expression. h TeM mRNA expression. i Phase-contrast microscope image. ACL cells were isolated from 3 different donors (n = 3). Cells were cultured for 5 days, and then their properties were analyzed. The data were normalized to those for GAPDH. Scale bar 250 μm *p < 0.05
Immunohistofluorescence analysis of ACL tissue
Autofluorescence of ACL tissue was not observed under these experimental conditions (Fig. 2a). Elastin, type I collagen, and type III collagen were widely distributed in ACL tissue (Fig. 2c–h). Type I collagen fibers and type III collagen fibers had a dense structure. In contrast, elastin fiber has a sparser structure than collagen fiber, and elastin fiber was observed to fill a gap among collagen fibers.

Immunohistofluorescence analysis of human ACL tissue and ECM scaffolds. a Autofluorescence of ACL. b Differential interference contrast microscope image. c, d Tropoelastin expression. e, f Type I collagen expression. g, h Type III collagen. Scale bars are a–h 250 μm. i, k Elastin-A scaffolds, j, l collagen scaffold. i and j are SEM images of ECM fibers. k and l are autofluorescence of ECM fibers. Scale bars are i 150 μm, j 100 μm, k and l 50 μm
Characteristics of cells outgrown from the ACL
Cells that migrated from the ACL during explant culture demonstrated a fibroblastic and spindle-like shape (Fig. 1i). Cell growth gradually decreased over 75 days (cumulative population doubling [cPD] level, 19). ACL cells showed little proliferative ability at approximately 130 days (cPD level, 23) (Fig. 1e).
ACL cells between passages 1 and 10 (cPD level, from 1.6 to 15) that showed proliferation were used to examine ligament-related gene expression. The expression levels of TeM, type I collagen, and type III collagen remained virtually constant in these cells (Fig. 1f–h).
Comparison of the cellular properties of ACL cells and PDLFs
Ligament-related genes (for TeM, type I collagen, and type III collagen) were almost equally expressed in ACL cells and PDLFs without significant differences (Fig. 3a–c). Type III collagen expression was approximately 2.6 and 3.2 times higher than type I collagen expression in ACL cells and PDLFs, respectively. In contrast, the ALP activity of PDLFs was approximately 2.1 times higher than that of ACL cells (Fig. 3d).

Comparison of the cellular properties of ACL cells and PDLFs. a Type I collagen mRNA expression. b Type III collagen mRNA expression. c TeM mRNA expression. d ALP activity. a, b, c n = 3. Cells were cultured for 5 days. The data were normalized to those for GAPDH. d ACL cells, n = 7; PDLFs, n = 4. *p < 0.05
Structure of the ECM scaffolds
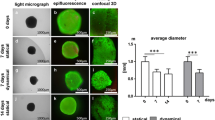
SEM images of the elastin-A scaffold and collagen scaffold are shown in Fig. 2i and j, respectively. We described the average diameter of fibers in the ECM scaffolds in our previous report [13], and the average fiber diameter in the elastin-A and collagen scaffolds were 0.5 and 1.7 μm, respectively. Figure 2k, l show the autofluorescence of the elastin and collagen fibers, and it was observed that their structure was similar to that of the ACL organization and fiber orientation.
Cellular responses to the ECM scaffold
Type I collagen expression from ACL cells on the elastin-A scaffold was slightly higher than that from ACL cells on collagen scaffold (Fig. 4a). Very little difference was detected in the production of type III collagen on the elastin-A scaffold (Fig. 4b). No difference in type I and type III collagen expression from PDLFs was observed between PDLFs on the elastin-A and collagen scaffolds (Fig. 4e, f).

Responses of ACL cells and PDLFs to the ECM scaffold. ACL cells, a type I collagen mRNA expression. b Type III collagen mRNA expression. c TeM expression. d ALP activity. a, b, c Elastin-A scaffold, n = 5; collagen scaffold, n = 8. The data were normalized to those for GAPDH. d Elastin-A scaffold, n = 6; collagen scaffold, n = 7. PDLFs, e type I collagen mRNA expression. f Type III collagen mRNA expression. g TeM expression. h ALP activity. e, f Elastin-A scaffold, n = 11; collagen scaffold, n = 9. g Elastin-A scaffold, n = 9; collagen scaffold, n = 7. The data were normalized to those for GAPDH. h Elastin-A scaffold, n = 9; collagen scaffold, n = 8. ACL and PDLFs were cultured for 7 days on each scaffold type. *p < 0.05
The expression of TeM in ACL cells on an elastin-A scaffold was approximately 70 % lower than that in ACL cells on a collagen scaffold (Fig. 4c). In contrast, the ALP activity of ACL cells on the elastin-A scaffold was 1.8 times higher than that of cells on the collagen scaffold (Fig. 4d).
PDLFs showed cellular responses similar to those of ACL cells (Fig. 4e–h). The TeM expression of PDLFs on the elastin-A scaffold was approximately 93 % lower than that of PDLFs on the collagen scaffold (Fig. 4g). In contrast, PDLFs on the elastin-A scaffold showed 1.8 times higher ALP activity than those on the collagen scaffold (Fig. 4h).
Discussion
Structure of the ECM scaffolds
The value (0.3 MPa) of the elastic modulus of the elastin-A scaffold used in this study was similar to that of neck ligament tissue (0.2 MPa), which is an elastin-rich material described in our previous report [13]. The collagen scaffold, which has an elasticity of 1.2 MPa, has the same strong physical properties as a ligament.
Gene expression in ACL tissue and bone tissue
To determine the gene expression pattern of intact tissue, total RNA was extracted from ACL and bone tissue and measured by RT-PCR (Fig. 1). Higher expression of type III collagen and TeM was observed in ACL tissue than in bone tissue (Fig. 1b, c). Type III collagen is the main component of ligaments, and TeM is a marker of dense connective tissue such as ligaments and tendons. In contrast, the expression of ALP, which is an osteogenic marker, in bone tissue was higher than that in ACL tissue (Fig. 1d). It is therefore confirmed that type III collagen and TeM can be used as markers to evaluate ligament cell phenotype and ALP can be used as a marker to evaluate osteoblast phenotype.
Immunohistofluorescence analysis of ACL tissue
Figure 2 shows that elastin, type I collagen, and type III collagen are widely distributed in ACL tissue. Thus, ligament cells in ACL tissue are likely surrounded by elastin and types I and III collagen. To date, elastin has not been focused on as a material for artificial ligament engineering. However, both elastin and collagen were found to be distributed throughout ACL tissue (Fig. 2c–h). Therefore, it is important to use elastin as well as collagen to prepare artificial ligaments that mimic the ligament structure. Therefore, cellular responses to elastin and type I collagen, which is the main collagen component of ligaments, were examined using ECM-supplemented culture medium and ECM scaffolds in this study.
Characteristics of cells outgrown from ACL
The cell morphological features and cumulative population doubling of outgrown ACL cells agreed with the findings of Cheng et al. [7] and Steinert et al. [8]. The results of gene expression and proliferative assays suggest that isolated ACL cells have a high capacity for proliferation and undergo fewer phenotypic changes during subculture.
Comparison of the cellular properties of ACL cells and PDLFs
PDLFs have higher collagen synthesis ability and ALP activity than other types of fibroblasts, e.g., gingival fibroblasts. In addition, PDLFs have the capacity to form mineralized tissue [4–6]. Figure 3a–c showed that ligament-related gene expression levels were not significantly different between ACL cells and PDLFs. In contrast, the ALP activity of PDLFs was significantly higher than that of ACL cells (Fig. 3d). These results suggest that ACL cells and PDLFs would have similar synthesis ability for matrix protein; however, PDLFs have higher osteoblastic function than ACL cells. The results show that PDLFs have an advantage over ACL cells in the reconstruction of the bone interface because of their osteoblastic functions. Further detailed study is required regarding the osteogenic potential of PDLFs and their ability to synthesize bone-specific proteins.
Cellular responses to the ECM scaffold
A significant difference was observed in the TeM expression in ligament cells on the elastin-A scaffold and ligament cells on the collagen scaffold (Fig. 4c, g). TeM is an antiangiogenic factor that is expressed in dense connective tissues such as ligament and tendon and is known as a ligament marker [18–22]. Figure 4 shows that ligament cells on the elastin-A scaffold hardly showed TeM expression relative to cells on the collagen scaffold. However, ligament cells on the elastin-A scaffold showed significantly higher ALP activity than cells on the collagen scaffold. ALP is known as an osteogenic marker [23]. As previously reported, ACL cells and PDLFs have the potential to differentiate into osteoblasts, and they express ALP in the process of osteogenic differentiation [7–9, 12]. Furthermore, bone tissue showed higher ALP expression than ACL tissue (Fig. 1d). Therefore, ligament cells on an elastin-A scaffold would likely differentiate into osteoblast-like cells. In contrast, like the ACL tissue in Fig. 1c, the ligament cells on the collagen scaffold showed TeM expression. Thus, ligament cells on a collagen scaffold would maintain their phenotype as ligament cells.
Osteogenic induction by the elastin-A scaffold and cell phenotype maintenance by a collagen scaffold were measured in both ACL cells and PDLFs. Ligaments join bone to bone; therefore, it would be useful to mimic the interface structure between ligament and bone by differentiating ligament cells to osteoblasts. The results of this study indicate that the elastin-A scaffold would be useful for reconstructing the interface region. Furthermore, it is considered that elastin is an important factor for obtaining mechanical properties equivalent to those of native ACL.
In general, ligaments play an important role in stabilizing joints and enabling complex movement. Therefore, ligaments possess stiffness and high elasticity. Elastin plays a central role in elasticity in the living body and is widely distributed in ACL tissue (Fig. 2). Thus, it is expected that the mechanical properties of a cell scaffold for an artificial ligament equivalent to the native ACL would be obtained with a combination of collagen scaffolds (stiffness) and elastin-A scaffolds (elasticity).
ACL cells and PDLFs showed similar properties (ligament-related gene expression) and responses to ECM. However, the ALP activity and TeM expression of PDLFs on the elastin-A scaffold was higher and lower than that of ACL cells on the elastin-A scaffold, respectively (Fig. 4). PDLFs show phenotype changes in response to ECM. In particular, it is expected that PDLFs will differentiate into osteoblasts effectively by combining with the elastin-A scaffold (Fig. 4g, h). These results suggest that PDLFs would be useful as the cell source for ACL reconstruction using tissue engineering. Depending on the degree of injury of ACL tissue, ACL cells may not always be available. PDLFs can be isolated from the PDL, and the PDL is easier to harvest than other ligament tissues such as the ACL. Therefore, it is expected that PDLFs will be useful as an alternative to ACL cells.
Conclusion
In this study, ACL cells and PDLFs were found to have similar cellular functions. Since ACL cells and PDLFs can be used almost interchangeably, although ACL cells are important, an important point that should be clarified is that PDLFs can be used as a substitute for the ACL when the ACL cells cannot be used. In addition, with regard to the interaction between ligament cells and ECM, the elastin-A scaffold increased ALP activity and downregulated TeM expression in ligament cells. In contrast, the collagen scaffold maintained TeM expression. In particular, an elastin-A scaffold was suitable for the bone-ligament jointing section, and a collagen scaffold was suitable for the central portion. Therefore, it is expected that the ECM scaffolds prepared in this study will be useful as a cell scaffolds for ligament reconstruction that can promote phenotype alteration of ligament cells.
References
Hadjicostas PT, Soucacos PN, Koleganova N, Krohmer G, Berger I. Comparative and morphological analysis of commonly used autografts for anterior cruciate ligament reconstruction with the native ACL: an electron, microscopic and morphologic study. Knee Surg Sports Traumatol Arthrosc. 2008;16:1099–107.
Duthon VB, Barea C, Abrassart S, Fasel JH, Fritschy D, Menetrey J. Anatomy of the anterior cruciate ligament. Knee Surg Sports Traumatol Arthrosc. 2006;14:204–13.
Laurencin CT, Freeman JW. Ligament tissue engineering: an evolutionary materials science approach. Biomaterials. 2005;26:7530–6.
Somerman MJ, Archer SY, Imm GR, Foster RA. A comparative study of human periodontal ligament cells and gingival fibroblasts in vitro. J Dent Res. 1988;67:66–70.
Lekic P, McCulloch CA. Periodontal ligament cell population: the central role of fibroblasts in creating a unique tissue. Anat Rec. 1996;245:327–41.
Ramakrishnan PR, Lin WL, Sodek J, Cho MI. Synthesis of noncollagenous extracellular matrix proteins during development of mineralized nodules by rat periodontal ligament cells in vitro. Calcif Tissue Int. 1995;57:52–9.
Cheng MT, Yang HW, Chen TH, Lee OK. Isolation and characterization of multipotent stem cells from human cruciate ligaments. Cell Prolif. 2009;42:448–60.
Steinert AF, Kunz M, Prager P, Barthel T, Jakob F, Noth U, Murray MM, Evans CH, Porter RM. Mesenchymal stem cell characteristics of human anterior cruciate ligament outgrowth cells. Tissue Eng Part A. 2011;17:1375–88.
Zhang J, Pan T, Im HJ, Fu FH, Wang JH. Differential properties of human ACL and MCL stem cells may be responsible for their differential healing capacity. BMC Med. 2011;9:68.
Tanaka K, Iwasaki K, Feghali KE, Komaki M, Ishikawa I, Izumi Y. Comparison of characteristics of periodontal ligament cells obtained from outgrowth and enzyme-digested culture methods. Arch Oral Biol. 2011;56:380–8.
Wu LP, Wei X, Ling JQ, Liu L. The surface antigen expression of periodontal ligament cells and dental pulp cells in vitro. Hua Xi Kou Qiang Yi Xue Za Zhi. 2009;27:20–3.
Zhao Y, Wang C, Li S, Song H, Wei F, Pan K, Zhu K, Yang P, Tu Q, Chen J. Expression of Osterix in mechanical stress-induced osteogenic differentiation of periodontal ligament cells in vitro. Eur J Oral Sci. 2008;116:199–206.
Mizutani N, Kawato H, Maeda Y, Takebayashi T, Miyamoto K, Horiuchi T. Multiple-type dynamic culture of highly oriented fiber scaffold for ligament regeneration. J Artif Organs. 2013;16:49–58.
Miyamoto K. Crosslinked elastin and process for producing the same. US Patent 7,125,960 B2 2006-10-24.
Miyamoto K, Atarashi M, Kadozono H, Shibata M, Koyama Y, Okai M, Inakuma A, Kitazono E, Kaneko H, Takebayashi T, Horiuchi T. Creation of cross-linked electrospun isotypic-elastin fibers controlled cell-differentiation with new cross-linker. Int J Biol Macromol. 2009;45:33–41.
Stylianou E, Jenner LA, Davies M, Coles GA, Williams JD. Isolation, culture and characterization of human peritoneal mesothelial cells. Kidney Int. 1990;37:1563–70.
Haddad-Weber M, Prager P, Kunz M, Seefried L, Jakob F, Murray MM, Evans CH, Noth U, Steinert AF. BMP12 and BMP13 gene transfer induce ligamentogenic differentiation in mesenchymal progenitor and anterior cruciate ligament cells. Cytotherapy. 2010;12:505–13.
Itaya T, Kagami H, Okada K, Yamawaki A, Narita Y, Inoue M, Sumita Y, Ueda M. Characteristic changes of periodontal ligament-derived cells during passage. J Periodontal Res. 2009;44:425–33.
Kimura N, Shukunami C, Hakuno D, Yoshioka M, Miura S, Docheva D, Kimura T, Okada Y, Matsumura G, Shin’oka T, Yozu R, Kobayashi J, Ishibashi-Ueda H, Hiraki Y, Fukuda K. Local tenomodulin absence, angiogenesis, and matrix metalloproteinase activation are associated with the rupture of the chordae tendineae cordis. Circulation. 2008;118:1737–47.
Shukunami C, Takimoto A, Miura S, Nishizaki Y, Hiraki Y. Chondromodulin-I and tenomodulin are differentially expressed in the avascular mesenchyme during mouse and chick development. Cell Tissue Res. 2008;332:111–22.
Shukunami C, Hiraki Y. Chondromodulin-I and tenomodulin: the negative control of angiogenesis in connective tissue. Curr Pharm Des. 2007;13:2101–12.
Burgess R, Cserjesi P, Ligon KL, Olson EN. Paraxis: a basic helix-loop-helix protein expressed in paraxial mesoderm and developing somites. Dev Biol. 1995;168:296–306.
Wlodarski KH, Reddi AH. Alkaline phosphatase as a marker of osteoinductive cells. Calcif Tissue Int. 1986;39:382–5.
Acknowledgments
This study was supported by a research grant (Yoshimi Memorial TMP Grant, 2010) from the Japanese Society for Artificial Organs (JSAO).
Conflict of interest
The authors have declared that no conflict of interest exists.
Author information
Authors and Affiliations
Corresponding authors
Rights and permissions
About this article
Cite this article
Mizutani, N., Kageyama, S., Yamada, M. et al. The behavior of ligament cells cultured on elastin and collagen scaffolds. J Artif Organs 17, 50–59 (2014). https://doi.org/10.1007/s10047-013-0736-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10047-013-0736-y