
Abstract
In many terrestrial ecosystems, large amounts of leaf litter are consumed by macroarthropods. Most of it is deposited as faeces that are easily transferred into deeper soil layers. However, the decomposition of this large pool of organic matter remains poorly studied. We addressed the question of how leaf litter transformation into macroarthropod faeces, and their burial in the soil, affect organic matter decomposition in a Mediterranean dry shrubland. We compared mass loss of intact leaf litter of two dominant shrub species (Quercus coccifera, Cistus albidus) with that of leaf litter-specific faeces from the abundant millipede Ommatoiulus sabulosus. Leaf litter and faeces were exposed in the field for 1 year, either on the soil surface or buried at 5 cm soil depth. Chemical and physical quality of faeces differed strongly from that of leaf litter, but distinctively between the two shrub species. On the soil surface, faeces decomposed faster than intact leaf litter in Quercus, but at similar rates in Cistus. When buried in the soil, faeces and leaf litter decomposed at similar rates in either species, but significantly faster compared to the soil surface, most likely because of higher moisture within the soil enhancing microbial activity. The combined effects of leaf litter transformation into faeces and their subsequent burial in the topsoil led to a 1.5-fold increase in the annual mass loss. These direct and indirect macroarthropod effects on ecosystem-scale decomposition are likely more widespread than currently acknowledged, and may play a particularly important role in drought-influenced ecosystems.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The role of climate and plant litter quality in regulating decomposition has been thoroughly assessed in many studies and is reasonably well understood (Couteaux and others 1995; Parton and others 2007; Cornwell and others 2008). On the other hand, the impact of soil fauna is much less studied, despite being a major factor in driving decomposition as recent large-scale studies and meta-analyses suggest (Wall and others 2008; García-Palacios and others 2013; Handa and others 2014). However, a difficulty for the quantification of fauna effects on decomposition is the distinction between litter mass loss due to consumption by detritivores and the amount of organic matter that is actually mineralized. Even though macroarthropods can ingest a large proportion of the annual litter fall (Cárcamo and others 2000; David and Gillon 2002), a substantial amount of the initially consumed litter is returned to the soil as faeces composed of poorly digested litter material (Wolters 2000). As a result, these detritivores convert large quantities of intact leaf litter into faeces without substantially increasing the mineralization of this material. It is critical to understand the fate of faeces in order to correctly assess decomposition processes and how it affects carbon (C) sequestration and mineralization (Prescott 2005), in particular for ecosystems with high macroarthropod abundance.
Few studies have compared the long-term decomposition of macroarthropod faeces with that of intact leaf litter. In the short term (up to 2–3 weeks after egestion), the transformation of litter into faeces can stimulate microbial activity, with higher respiration rates in macroarthropod faeces than in intact leaf litter (Maraun and Scheu 1996; Frouz and Simek 2009). In longer term experiments, however, microbial respiration was found to be similar or even lower in faeces than in intact leaf litter (Scheu and Wolters 1991; Špaldoňová and Frouz 2014). The few field studies explicitly measuring decomposition over a longer period of time were conducted exclusively in temperate ecosystems and reported contrasting results. Two studies with millipedes found no differences between faeces and leaf litter mass loss rates after 1 year (Nicholson and others 1966; Webb 1977), and another study on dipteran larvae (Bibionidae) found a lower mass loss rate in faeces compared to leaf litter after 11 months (Frouz and Simek 2009). After the initial flush of microbial activity, ageing macroarthropod faeces may have lower respiration and mineralization rates than leaf litter, both because of their compaction and as a result of accumulating recalcitrant compounds, such as lignin (Lavelle 1997). However, changes in organic matter quality after gut passage may vary among detritivore species. For example, Zimmer and others (2002) found that different woodlouse species differed in their ability to digest litter phenolic compounds. Additionally, the degree of stimulation of microbial activity in faeces may vary depending on the quality or the species of leaf litter consumed, as suggested by results from several short-term studies (Hassall and others 1987; Maraun and Scheu 1996; Joly and others 2015).
So far, all studies focusing on macroarthropod faeces decomposition were carried out either under controlled laboratory conditions or on the soil surface in the field. However, comparatively small-sized macroarthropod faeces may not stay at the soil surface. Instead, they can easily sediment passively by gravity and in percolating water or even be buried actively by soil animals down to deeper soil horizons (Anderson 1988). In laboratory experiments, juliform millipedes were shown to dig burrows down to 10 cm deep, in which they incorporated faecal pellets (Haacker 1967; Bowen and Hembree 2014). At this depth, microclimatic conditions differ considerably from those on the soil surface. These differences are important for microbial decomposers, and it has previously been shown that buried litter decomposed more rapidly than litter placed on the soil surface (Beare and others 1992; Ghidey and Alberts 1993), probably due to more favourable moisture conditions, and thus, improved microbial decomposition at greater soil depths (Beare and others 1992; Rovira and Vallejo 1997). Similarly, buried faeces are likely to decompose more rapidly than on the soil surface, and Hassall and others (1987) hypothesized that the translocation of faecal material into the soil profile might be the most important indirect contribution that macroarthropods make to decomposition processes. However, this hypothesis has never been tested so far. As there is evidence that the impact of organic matter burial on its decomposition varies inversely with the amount of rainfall (McInerney and Bolger 2000; Powers and others 2009; Lee and others 2014), the incorporation of faeces in deeper and moister microsites could be especially important in water-limited environments such as Mediterranean ecosystems, which are projected to become even drier in the coming decades.
Here we assessed the combined effects of leaf litter transformation into faeces by macroarthropods and burial of these materials on organic matter decomposition in a Mediterranean shrubland of southern France. In this type of ecosystem, the millipede Ommatoiulus sabulosus can be remarkably abundant and was shown to produce large amounts of faeces when feeding on shrub leaf litter (Coulis and others 2013). In a one-year field study, we evaluated annual mass loss for intact leaf litter of Cistus albidus and Quercus coccifera, the two most abundant shrub species in this area, and for faeces produced by Ommatoiulus feeding on each litter type. Moreover, leaf litter and faeces were left to decompose in litterbags either placed on the soil surface or buried to a depth of 5 cm. We hypothesized that decomposition over 1 year proceeds identically in macroarthropod faeces and intact leaf litter, irrespective of plant species identity. Additionally, we hypothesized that faeces and intact leaf litter both decompose faster when buried in the topsoil compared to when they remain on the soil surface. Consequently, at the ecosystem scale, the transformation of leaf litter into faeces, combined with the high probability of transfer of faeces to deeper soil horizons, should increase organic matter decomposition, a mechanism that should be particularly important in our Mediterranean ecosystem experiencing regular drought conditions.
Materials and Methods
Study Site
The study was conducted in a shrubland located at the Chaine de l’Etoile, 5 km northeast of Marseille, southern France (43°22′ N; 5°25′ E; 275 m a.s.l.). The climate is typically Mediterranean with a mean annual temperature of 15.5°C, a mean annual rainfall of 518 mm and a period of pronounced drought in summer (Figure 1). Climatic conditions during our experiment were similar to the long-term average with a mean annual temperature of 15.4°C and mean annual precipitation of 522 mm (Figure 1). The soil is a shallow rendzina on limestone (Montès and others 2008), in which rock fragments and stones represent 59% of the soil volume in the top 20 cm. The vegetation is a typical garrigue with an irregular cover of mostly shrubs less than 1.4 m in height and some perennial and annual herbaceous plants, with a total cover between 25 and 95% of the ground surface. Five plant species dominate the community and account for 97% of total cover: the woody shrubs Q. coccifera (36%), C. albidus (18%), Ulex parviflorus (10%), and Rosmarinus officinalis (9%) and the perennial grass Brachypodium retusum (24%) (N. Rodriguez, unpublished data). In this study, we focused on the two most abundant shrub species Q. coccifera and C. albidus. The saprophagous macrofauna community is dominated by the julid millipede O. sabulosus aimatopodus, a Mediterranean subspecies of O. sabulosus with no dorsal yellow bands and that has a phenology adapted to the Mediterranean climate (David and Coulis 2015). Its population density and biomass, which vary seasonally and annually, were estimated at 164 ± 37 individuals m−2 and 9.2 ± 2 g (live mass) m−2, respectively, in the spring of 2010.

Monthly precipitation during the experimental period (complete seasonal cycle from June 2012 to May 2013) plotted with the long-term means of monthly precipitation, evapotranspiration, and air temperature. Long-term means were calculated from a 30-year recording period (1982–2011). All data are from the meteorological station of Marignane (43°26′N, 5°12′E, 9 m a. s.l.), located 17 km away from the study site.
Leaf Litter Collection and Faeces Production
Quercus and Cistus leaf litter were collected on the ground at the study site in March and April 2011. Freshly fallen leaves were discarded, because litter-feeding macroarthropods generally prefer litter in later decomposition stages (Wolters 2000). Collected leaves were shed mostly during the previous year’s litter fall. This litter cohort, however, still consisted of intact leaves as decomposition proceeds comparatively slowly in this dry ecosystem. Leaf litter was air-dried in the laboratory, sorted into species, and adhering soil particles were brushed off.
Part of this material was used to produce faeces. Three batches of sixty grams of leaf litter each from both species (Quercus and Cistus) and 20 Ommatoiulus individuals (mean individual live weight 103 ± 0.2 mg) per batch and litter type, all collected at the study site, were incubated for 15 days at 20°C in transparent plastic boxes (40 × 33 × 8.5 cm). Every third day, millipedes were removed from the boxes and the material was sieved (2 mm mesh) for faeces collection. Leaf litter and millipedes were then put back in their box and sprayed with water to maintain optimal conditions for millipede activity. Another stock of Quercus and Cistus leaf litter was kept in plastic boxes without fauna under the same conditions as those used for faeces production. After 15 days, we produced a total of 8 and 16 g of faeces from Quercus and Cistus leaf litter, respectively. Leaf litter from the boxes without millipedes and millipede faeces were air-dried and stored at room temperature in the dark until the beginning of the experiment.
Leaf Litter and Faeces Quality
A number of initial quality parameters were determined on leaf litter and faeces materials. Water holding capacity (WHC), expressed as a percentage of dry mass, was determined by soaking samples of leaf litter and faeces for 24 h in distilled water using a mass to volume ratio of 1 g:50 ml. Litter and faeces were drained to remove surface water, weighed moist, and reweighed after drying at 60°C for 48 h. Soaking water was filtered through 0.45-μm cellulose nitrate membrane filters and analysed for dissolved organic carbon (DOC) and total dissolved nitrogen (TDN) using a TOC analyser equipped with a supplementary module for N (CSH E200V, Shimadzu, Kyoto, Japan). The filtrate was also used to determine an index of DOC aromaticity following the protocol of Weishaar and others (2003). Specific UV absorbance at 280 nm (SUVA280) was measured using an UV spectrophotometer (Helios Gamma, Thermospectronic, Cambridge, UK) and calculated as SUVA280 = absorbance at 280 nm/DOC (in L g−1 cm−1).
Further analyses were performed on oven-dry leaf litter and faeces materials, which were both ground to a uniform particle size of 1 mm using a Cyclotec sample mill (Foss Tecator, Höganäs, Sweden). Total C and N concentrations were measured using a flash elemental analyser (EA1112 Series, Thermo Finnigan, Milan, Italy). Ash content was measured gravimetrically after combustion for 3 h at 550°C. Organic matter composition in leaf litter and faeces were assessed by solid-state 13C cross polarization magic-angle spinning (13C CPMAS) NMR spectroscopy (Alarcón-Gutiérrez and others 2009). Chemical shift values were referenced to tetramethylsilane and calibrated with glycine carbonyl signal at 176.05 ppm. The 13C-NMR spectra were divided into the following chemical shift regions (Mathers and others 2007; Alarcón-Gutiérrez and others 2009): alkyl C (10–45 ppm), N-alkyl and methoxyl C (45–60), O-alkyl C (60–110 ppm), aromatic C (110–140 ppm), phenolic C (140–160 ppm), and carboxyl C (160–190 ppm). Interpretation of litter components (for example, polysaccharides and lignin) was based on previous NMR studies (Mathers and others 2007; Preston and others 2009). The alkyl C:O-alkyl C ratio was used as an index of decomposition (Preston and others 2009).
Field Decomposition Experiment
Litterbags (5 × 4 cm) were made of 68 µm nylon mesh (68PES4/135, DIATEX, St-Genis-Laval, France). This mesh size prevents macro- and mesofauna from entering bags and also prevents the loss of very small fragments. Litterbags were filled with either 300 ± 10 mg of leaf litter or 200 ± 10 mg of faeces, which was equivalent to about 22 leaves of Quercus, 13 leaves of Cistus, and 1000 faecal pellets of Ommatoiulus. To compare leaf litter and faeces decomposition rates in the field, 144 litterbags were installed in 18 plots at the study site, all dominated by Quercus and Cistus. In each plot, there were two litterbags per type of substrate (Quercus leaf litter, Quercus faeces, Cistus leaf litter, and Cistus faeces), one on the soil surface and the other at 5 cm depth. The soil was carefully removed to a depth of 5 cm in a 25 × 25 cm area, four litterbags (one of each substrate) were placed horizontally, and the initially removed soil was put back. The four remaining litterbags (one of each substrate) were then placed flat on the soil that was put back within the same 25 × 25 cm area and fixed with stainless steel wire netting (1 cm mesh). All litterbags were installed on 16 May 2012 and removed on 3 June 2013 (that is, after 383 days).
An additional 42 litterbags were installed in six of the total 18 plots to determine the water content of leaf litter and faeces during the experiment. Half of these litterbags were placed 5 cm deep in the top soil and half were placed on the soil surface, in the same way as previously described, but in a different 25 × 25 cm area. They were collected in autumn (25 October 2012), during a period of favourable climatic conditions for decomposition.
After removal, the litterbags were carefully cleaned of adhering soil particles before being enclosed in sealed flasks, taken to the laboratory, weighed fresh, dried at 40°C for 72 h, and reweighed to determine the final dry mass and calculate the water content. The final ash content was determined for each individual litterbag to correct for potential contamination by soil particles. Mass loss was calculated as the difference between initial dry mass corrected for litter-specific ash content (Table 2) and final dry mass corrected for ash content in each litterbag. The mass loss rate is thus the percentage of organic matter lost over the duration of the experiment.
Microclimatic conditions were assessed in all 18 experimental plots from autumn to spring, the most favourable period for decomposition. In each plot, a button-type data logger (Signatrol, Tewkesbury, UK) protected from solar radiation by a custom-made screen, was placed 2 cm above the ground next to the litterbags, and recorded air temperature and relative humidity every second hour. Data were converted into daily averages, maxima, and minima. Soil moisture was also measured in all experimental plots at three dates in April and May 2013. On each occasion, a soil core (diameter 1 cm, length 10 cm) adjacent to the litter bags was sampled. The soil cores were immediately enclosed in sealed flasks, taken to the laboratory, weighed fresh, and reweighed after drying at 105°C to determine the gravimetric soil water content (% of oven-dried mass). Microclimatic data are summarized in Table 1.
Statistical Analyses
Differences in initial quality parameters between leaf litter and faeces were tested using two-way ANOVA with species identity (Quercus vs. Cistus) and substrate type (leaf litter vs. faeces) as fixed factors. Tukey’s HSD test was used for multiple comparisons among pairs of means. Differences in mass loss rates were tested using a mixed model four-way ANOVA. Experimental plots were considered as a random blocking factor, which was crossed with the three fixed factors, litterbag placement (surface vs. buried), substrate type (leaf litter vs. faeces), and species identity (Quercus vs. Cistus). The same ANOVA model was used to test for differences in the water content of leaf litter and faeces in autumn and spring. Relationships between microclimatic variables and mass loss rates were tested using ordinary least square regression. Before ANOVAs and regression analyses, normality of residuals and homogeneity of variances were checked and data were log or power transformed if required. All statistical analyses were performed using R software version 3.0.1 (R Development Core Team 2013).
Results
Initial Quality of Leaf Litter and Faeces
Almost all quality parameters differed significantly between Quercus and Cistus leaf litter (Table 2). The ash content was lower in leaf litter of Quercus compared to that of Cistus. Both carbon (C) and nitrogen (N) concentrations were higher, and C:N ratio lower, in Quercus than in Cistus leaf litter. The WHC was lower in Quercus compared to Cistus. Concentrations of DOC and TDN in leachates were substantially higher in Quercus than in Cistus leaf litter. However, leaf litter of both species had approximately the same aromaticity of DOC (Table 2).
Leaf litter ingestion and gut passage led to a considerably higher ash content in faeces compared to that in initial leaf litter in both species, but the relative change was more important in Quercus compared to Cistus (Table 2). The C concentrations were lower in faeces than in leaf litter in both species. In contrast, the N concentration was higher in faeces compared to that in initial leaf litter in Quercus but not in Cistus (Table 2). In both species, the WHC increased in faeces compared to leaf litter. The DOC and TDN were higher in leachates from Quercus faeces than in its litter, whereas differences between leaf litter and faeces were not significant for Cistus. In both species, but much more so in Quercus, DOC aromaticity increased in leachates of faeces compared to leaf litter.
In accordance with the overall larger differences between leaf litter and faeces characteristics observed in Quercus than in Cistus, 13C-NMR spectra of leaf litter and faeces differed markedly for Quercus, but were much more similar for Cistus (Figure 2). The only region of the Quercus leaf litter spectrum that was little affected by gut passage was the alkyl C region (10–45 ppm) corresponding to lipid, wax, and aliphatic carbons. At 33 ppm, the signal was even stronger in faeces than in leaf litter, while it was clearly the opposite (stronger signal in leaf litter than in faeces) across the rest of the spectrum. The spectral region that was the most affected by gut passage was the O-alkyl C region (60–110 ppm), corresponding to C from structural polysaccharides (for example, cellulose and hemicellulose), which strongly decreased in Quercus faeces compared to leaf litter. The decrease was much less pronounced in Cistus faeces compared to its leaf litter. Likewise, the alkyl C:O-alkyl C ratio was higher in faeces than in leaf litter, but much more for Quercus than for Cistus.

13C CPMAS NMR spectra of Quercus and Cistus leaf litter and of faeces produced by Ommatoiulus feeding on either of these two litter species. Dotted lines show the division into chemical shift regions.
Leaf Litter and Faeces Mass Loss
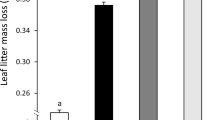
The placement of litterbags strongly influenced decomposition (Table 3), with a 32% higher mass loss in buried materials than on the soil surface (Figure 3). Averaged across the two types of substrates and the two species, annual mass loss was 34.5 ± 0.7% on the soil surface compared to 45.6 ± 1.2% at 5 cm depth. However, the placement effect was more marked in leaf litter than in faeces, resulting in a significant placement × substrate interaction (Table 3).

Mass loss of Quercus leaf litter, Cistus leaf litter, and faeces derived from each litter type placed on the soil surface and buried at 5 cm soil depth (mean ± SE, n = 18). Different letters indicate significant differences within each sub-figure (Tukey HSD test, P > 0.05).
Species identity and substrate type also had a significant effect on decomposition, with a clear interaction between the two factors (Table 3). Overall, Cistus decomposed more rapidly than Quercus and faeces decomposed more rapidly than leaf litter. However, the higher mass loss of Cistus occurred mainly in leaf litter, while the difference between species was much smaller in faeces. Pairwise comparisons showed that mass loss was generally similar between leaf litter and faeces, with one noticeable exception: on the soil surface, Quercus faeces decomposed significantly more rapidly than Quercus leaf litter (P < 0.001) (Figure 3).
Microclimate Effects on Mass Loss
The observed greater mass loss at 5 cm soil depth corresponded with clearly higher water contents of buried materials compared to surface materials, both in autumn 2012 and spring 2013 (Figure 4). ANOVAs showed that most of the variation in leaf litter and faeces water content was explained by litterbag placement (P < 0.001). In addition, at high moisture levels in autumn, species identity and substrate type also significantly influenced the water content, which was 83% higher in Cistus than in Quercus (P < 0.001) and 18% higher in faeces than in leaf litter (P < 0.001) (data not shown).

Mass loss and water content of leaf litter and faeces (averaged across both substrates) placed on the soil surface and at 5 cm soil depth. Whiskers show 1.5 times the interquartile range and outliers are shown as circles.
Microclimatic records collected adjacent to the litterbags showed substantial variations among plots (Table 1). From autumn to spring, the warmest plot was on average 1.4°C warmer than the coolest plot. The most humid plot had a mean relative air humidity of 89.8 versus 81.4% for the driest plot. Soil moisture variations in spring were even more pronounced, with as much as double the moisture in some plots compared to others. However, linear regression analyses showed that leaf litter and faeces mass loss rates were not significantly related to any of these variables, regardless of whether they decomposed on the soil surface (P ≥ 0.06) or at 5 cm depth (P ≥ 0.14) (data not shown).
Discussion
Decomposition of Leaf Litter and Macroarthropod Faeces
Although it is commonly assumed that litter-feeding macroarthropods enhance decomposition by stimulating microbial activity in their faeces, there is little experimental evidence in support of this view (David 2014). A priori predictions based on chemical and physical changes following gut passage are difficult, because macroarthropod faeces, when compared with intact leaf litter, have characteristics that may either stimulate or inhibit decomposition. Specifically, fresh faeces have higher water contents than leaf litter (McBrayer 1973; Suzuki and others 2013), lower tannin contents (Zimmer and others 2002; Coulis and others 2009), and, in some cases, lower C:N ratios (Kautz and others 2002; Bastow 2011), which tends to accelerate decomposition. On the other hand, faeces are poor in compounds that are readily available to microorganisms, such as sugars, and relatively enriched in lignin (Gillon and David 2001; Rawlins and others 2006), which tends to slow decomposition. In the present study, we showed that transformation of Mediterranean shrub litter into faeces by O. sabulosus distinctly affected organic matter quality. The most striking difference between Quercus and Cistus faeces quality changes was a significant increase in N concentration in faeces compared to leaf litter in Quercus, whereas it tended to decrease in Cistus, leading to a much lower C:N ratio in Quercus faeces (C:N of 34) than in Cistus faeces (C:N of 51). Increased faeces N concentration in Quercus was accompanied with a substantially higher total dissolved N concentration in faeces leachates compared to leaf litter leachates, which was six times greater than that in Cistus faeces leachates (Table 2). In addition to these large differences in faeces N quality, Quercus faeces also changed much more in overall C quality than Cistus faeces compared to their respective leaf litter materials, as indicated by the distinct shift in 13C CPMAS NMR and a higher concentration of DOC in leachates from Quercus faeces. The DOC leached from faeces also had a greater aromaticity than DOC leached from leaf litter, which may suggest a higher amount of recalcitrant compounds in faeces. Collectively, our results showed that changes in organic matter quality after gut passage were much more pronounced for Quercus than for Cistus leaf litter, suggesting more important consequences for further decomposition when detritivores transform Quercus leaf litter into faeces.
Accordingly, Ommatoiulus faeces derived from Quercus lost more mass in our one-year field study than Quercus leaf litter when placed on the soil surface. In contrast, faeces derived from Cistus decomposed at approximately the same rate as Cistus leaf litter, regardless whether the material was placed on the soil surface or 5 cm deep in the top soil. These findings are only in part in line with our initial hypothesis stating that leaf litter and faeces produced from the same leaf litter decompose at the same rate. In previous studies conducted on other combinations of leaf litter and macroarthropod species, transient stimulations of microbial activity were sometimes reported in fresh macroarthropod faeces, but so far none of the only few long-term studies reported increased mass loss rates in faeces compared to intact leaf litter (Nicholson and others 1966; Webb 1977; Frouz and others 2015). Our results based on leaf litter from Q. coccifera, the dominant shrub species in our Mediterranean dry shrubland system, indicate that litter-feeding macroarthropods can significantly promote the long-term decomposition of organic matter.
The 32% greater mass loss in Quercus faeces compared to leaf litter decomposing on the soil surface may be explained in several ways. (1) As outlined above, we observed clear changes in chemical composition of faeces compared to intact leaf litter, in particular the lower C:N ratio and the higher DOC and TDN concentrations, which might have stimulated microbial activity. However, the same quality differences between Quercus faeces and leaf litter did not result in the same effect in buried material, perhaps because microbial limitations are different in the litter layer and within the soil (Fanin and others 2012). (2) Another possible explanation may be greater leaching of water-soluble organic matter from Quercus faeces than from leaf litter. Leaching contributes importantly to mass loss during decomposition (Swift and others 1979), and our data showed that the initial amounts of DOC and TDN were larger in faeces than leaf litter in Quercus but not in Cistus. These leachates may contribute to higher mass loss from Quercus faeces and, arguably, leaching is greater for materials exposed to rainfall at the soil surface than for buried materials. (3) Finally, the 42% increase in WHC in Quercus faeces compared to leaf litter also might have had a positive effect on microbial activity, an effect which may be more important at the soil surface than within the overall moister soil. However, a similar increase of 32% in WHC in faeces from Cistus leaf litter did not correspond to a similarly increased faeces decomposition, even though there was a trend of a slightly greater faeces than leaf litter mass loss at the soil surface. At present, it is difficult to identify the overriding factor for a faster faeces than leaf litter decomposition when Quercus material is exposed at the soil surface, and it appears that several chemical and physical characteristics of faeces contribute to this response. Further studies would be needed to more precisely pinpoint the underlying mechanisms.
Effect of Organic Matter Burial on Decomposition
Our data clearly showed that the placement of litterbags (that is, on the soil surface vs within the topsoil) strongly influenced the decomposition of leaf litter and macroarthropod faeces. In line with our initial hypothesis, mass loss was consistently greater in buried materials, irrespective of their nature or initial quality. These results for leaf litter are in line with a previous experiment from a drought-influenced Mediterranean system, reporting higher litter decomposition rates with increasing soil depth (Rovira and Vallejo 1997). It seems reasonable to argue that burial of organic matter within the soil profile buffers moisture deficits and consequently improves conditions for microbial decomposers in drought-influenced ecosystems (Lee and others 2014), except perhaps when physical degradation through UV-radiation makes an important contribution to decomposition in more arid ecosystems (Austin and Vivanco 2006). In fact, several studies showed that greater decomposition rates within the soil were associated with a higher water content in buried than in surface plant litter, especially during dry periods (Beare and others 1992; Rovira and Vallejo 1997; McInerney and Bolger 2000). These results seem to confirm that more favourable moisture conditions within the soil are the driving factor for the positive effect of burial on organic matter decomposition. In line with this interpretation, we measured consistently higher water contents in buried materials than in surface materials, both during a rather wet period in autumn and a relatively dry period in spring.
Intact plant leaf litter, however, is not easily transported down the soil profile, and the previous studies burying leaf litter at different soil depths did not represent the natural conditions of our study system. Apart from chemical changes during the transformation of intact leaf litter into faeces, the physical aspect of the organic material is fundamentally altered. Most obviously, the commonly thin leaves of a relatively large surface area are transformed to small spherical or cylindrical faecal pellets of about 1 mm diameter, which in contrast to intact leaves can move more easily to deeper soil horizons (Anderson 1988). This displacement of organic material in the form of faeces from the soil surface down to deeper soil layers has been argued to be a major mechanism of accelerated decomposition of faeces (Hassall and others 1987). Surprisingly, this mechanism was never tested in the field, despite the potentially large impact on ecosystem C cycling and nutrient turnover. By placing faecal pellets of the dominant macroarthropod at our study site, both at the soil surface and 5 cm deep in the topsoil and following their decomposition, we have tested this hypothesis for the first time. Our results showed that faecal pellets from either of the two shrub species studied, decomposed more rapidly in the soil than at the soil surface, suggesting that faeces burial is an important mechanism by which macroarthropods can indirectly affect decomposition processes.
The chosen soil depth of 5 cm for burying faecal pellets in our study corresponds to quite realistic conditions for millipede faeces. For example, in a tropical dry forest with a thick litter layer, Loranger (2001) found large amounts of faecal pellets of millipedes between 4 and 8 cm below the soil surface. In addition to the passive transfer of faecal pellets that were deposited at the soil surface to deeper soil horizons, soil macroarthropods may also deposit their faeces directly within the soil. Diurnal vertical migrations between the upper litter layers and deeper shelters are common for soil macroarthropods during their activity period (Hassall and others 1987; Hopkin and Read 1992). The depth at which faeces are incorporated depends on the morphology and behaviour of animals, but many species move down to a depth of 5 cm or more, especially the juliform millipedes (Haacker 1967; Bowen and Hembree 2014). At our study site, we often observed Ommatoiulus burrowing at about 5 cm or slightly below in the mineral soil.
The ecosystem-scale consequences of macroarthropod activity can be substantial. On average across the two species, annual mass loss at the soil surface increased from 31 to 38% due to the transformation of leaf litter into faeces, and further increased to 46% as a result of the transfer of faeces to 5 cm soil depth. These values are indicative for the potential range of variation in macroarthropod effects on organic matter decomposition. However, for more realistic estimates, the proportion of litter that is actually consumed and buried by macroarthropods would need to be quantified. Based on the previously determined live biomass of O. sabulosus at our study site of 9.2 g m−2 and their litter consumption rates (Coulis and others 2013), the O. sabulosus population at our study site may consume up to 96 g m−2 of litter dry mass during their 4-month activity period per year (according to field observations). This would be more than half of the total annual litter fall of 162 g m−2 at our study site (M. Santonja, personal communication). These rough estimates indicate that the transformation of leaf litter into macroarthropod faeces is an important process in the studied Mediterranean shrubland ecosystem. Our data suggest that this would affect organic matter cycling even without any burial of faecal pellets, but depending on how much of the faeces will decompose below the soil surface, this indirect fauna-driven effect on decomposition may be augmented.
Conclusions
For a Mediterranean woody shrub-dominated ecosystem, we showed that the abundant macroarthropod O. sabulosus can have a strong impact on organic matter decomposition through the consumption of plant leaf litter and its transformation into faeces. The combined effects of chemical and physical changes of the organic matter when it is transformed from intact leaf litter into faecal pellets, and their subsequent transfer down to deeper soil layers, lead to a 1.5-fold increase in annual mass loss compared to intact leaf litter material decomposing at the soil surface. The transfer of the small faecal pellets to deeper soil layers appears as a particularly important mechanism of how macroarthropods may modify ecosystem C and nutrient cycling. These indirect macroarthropod-driven effects on decomposition are likely playing a much more important role than currently acknowledged, particularly in drought-influenced ecosystems.
References
Alarcón-Gutiérrez E, Floch C, Augur C, Petit JL, Ziarelli F, Criquet S. 2009. Spatial variations of chemical composition, microbial functional diversity, and enzyme activities in a Mediterranean litter (Quercus ilex L.) profile. Pedobiologia 52:387–99.
Anderson JM. 1988. Invertebrate-mediated transport processes in soils. Agric Ecosyst Environ 24:5–19.
Austin AT, Vivanco L. 2006. Plant litter decomposition in a semi-arid ecosystem controlled by photodegradation. Nature 442:555–8.
Bastow JL. 2011. Facilitation and predation structure a grassland detrital food web: the responses of soil nematodes to isopod processing of litter. J Anim Ecol 80:947–57.
Beare MH, Parmelee RW, Hendrix PF, Cheng W, Coleman DC, Crossley DA Jr. 1992. Microbial and faunal interactions and effects on litter nitrogen and decomposition in agroecosystems. Ecol Monogr 62:569–91.
Bowen JJ, Hembree DI. 2014. Neoichnology of two spirobolid millipedes: improving the understanding of the burrows of soil detritivores. Palaeontologia Electronica 17:1–48.
Cárcamo HA, Abe TA, Prescott CE, Holl FB, Chanway CP. 2000. Influence of millipedes on litter decomposition, N mineralization, and microbial communities in a coastal forest in British Columbia, Canada. Can J For Res 30:817–26.
Cornwell WK, Cornelissen JHC, Amatangelo K, Dorrepaal E, Eviner VT, Godoy O, Hobbie SE, Hoorens B, Kurokawa H, Pérez-Harguindeguy N. 2008. Plant species traits are the predominant control on litter decomposition rates within biomes worldwide. Ecol Lett 11:1065–71.
Coulis M, Hättenschwiler S, Fromin N, David JF. 2013. Macroarthropod-microorganism interactions during the decomposition of Mediterranean shrub litter at different moisture levels. Soil Biol Biochem 64:114–21.
Coulis M, Hättenschwiler S, Rapior S, Coq S. 2009. The fate of condensed tannins during litter consumption by soil animals. Soil Biol Biochem 41:2573–8.
Couteaux M-M, Bottner P, Berg B. 1995. Litter decomposition, climate and litter quality. Trends Ecol Evol 10:63–6.
David JF. 2014. The role of litter-feeding macroarthropods in decomposition processes: a reappraisal of common views. Soil Biol Biochem 76:109–18.
David JF, Coulis M. 2015. Millipedes faced with drought: the life cycle of a Mediterranean population of Ommatoiulus sabulosus (Linnaeus) (Diplopoda, Julida, Julidae). Zookeys 510:115–24.
David JF, Gillon D. 2002. Annual feeding rate of the millipede Glomeris marginata on holm oak (Quercus ilex) leaf litter under Mediterranean conditions. Pedobiologia 46:42–52.
Fanin N, Barantal S, Fromin N, Schimann H, Schevin P, Hättenschwiler S. 2012. Distinct microbial limitations in litter and underlying soil revealed by carbon and nutrient fertilization in a tropical rainforest. PLoS One 7:e49990.
Frouz J, Roubíčková A, Heděnec P, Tajovský K. 2015. Do soil fauna really hasten litter decomposition? A meta-analysis of enclosure studies. Eur J Soil Biol 68:18–24.
Frouz J, Simek M. 2009. Short term and long term effects of bibionid (Diptera: Bibionidae) larvae feeding on microbial respiration and alder litter decomposition. Eur J Soil Biol 45:192–7.
García-Palacios P, Maestre FT, Kattge J, Wall DH. 2013. Climate and litter quality differently modulate the effects of soil fauna on litter decomposition across biomes. Ecol Lett 16:1045–53.
Ghidey F, Alberts EE. 1993. Residue type and placement effects on decomposition : field study and model evaluation. Trans ASAE 36:1611–17.
Gillon D, David JF. 2001. The use of near infrared reflectance spectroscopy to study chemical changes in the leaf litter consumed by saprophagous invertebrates. Soil Biol Biochem 33:2159–61.
Haacker U. 1967. Tagesrhythmische Vertikalbewegung bei Tausendfüßlern (Myriapoda, Diplopoda). Naturwissenschaften 54:346–7.
Handa IT, Aerts R, Berendse F, Berg MP, Bruder A, Butenschoen O, Chauvet E, Gessner MO, Jabiol J, Makkonen M, et al. 2014. Consequences of biodiversity loss for litter decomposition across biomes. Nature 509:218–21.
Hassall M, Turner JG, Rands MRW. 1987. Effects of terrestrial isopods on the decomposition of woodland leaf litter. Oecologia 72:597–604.
Hopkin SP, Read HJ. 1992. The biology of millipedes. USA: Oxford University Press.
Joly FX, Coulis M, Gérard A, Fromin N, Hättenschwiler S. 2015. Litter-type specific microbial responses to the transformation of leaf litter into millipede feces. Soil Biol Biochem 86:17–23.
Kautz G, Zimmer M, Topp W. 2002. Does Porcellio scaber (Isopoda: Oniscidea) gain from coprophagy? Soil Biol Biochem 34:1253–9.
Lavelle P. 1997. Faunal activities and soil processes: adaptive strategies that determine ecosystem function. Advances in ecological research, Vol. 27. New York: Academic Press. p 93–132.
Lee H, Fitzgerald J, Hewins DB, McCulley RL, Archer SR, Rahn T, Throop HL. 2014. Soil moisture and soil-litter mixing effects on surface litter decomposition: a controlled environment assessment. Soil Biol Biochem 72:123–32.
Loranger G. 2001. Formes d’humus originales dans une forêt tropicale semi-décidue de la Guadeloupe. Comptes Rendus de l’Académie des Sciences - Series III - Sciences de la Vie 324:725–32.
Maraun M, Scheu S. 1996. Changes in microbial biomass, respiration and nutrient status of beech (Fagus sylvatica) leaf litter processed by millipedes (Glomeris marginata). Oecologia 107:131–40.
Mathers NJ, Jalota RK, Dalal RC, Boyd SE. 2007. 13C-NMR analysis of decomposing litter and fine roots in the semi-arid Mulga Lands of southern Queensland. Soil Biol Biochem 39:993–1006.
McBrayer JF. 1973. Exploitation of deciduous leaf litter by Apheloria montana (Diplopoda: Eurydesmidae). Pedobiologia 13:90–8.
McInerney M, Bolger T. 2000. Decomposition of Quercus petraea litter: influence of burial, comminution and earthworms. Soil Biol Biochem 32:1989–2000.
Montès N, Maestre FT, Ballini C, Baldy V, Gauquelin T, Planquette M, Greff S, Dupouyet S, Perret JB. 2008. On the relative importance of the effects of selection and complementarity as drivers of diversity–productivity relationships in Mediterranean shrublands. Oikos 117:1345–50.
Nicholson PB, Bocock KL, Heal OW. 1966. Studies on the decomposition of the faecal pellets of a millipede (Glomeris marginata (Villers)). J Ecol 54:755–66.
Parton W, Silver WL, Burke IC, Grassens L, Harmon ME, Currie WS, King JY, Adair EC, Brandt LA, Hart SC. 2007. Global-scale similarities in nitrogen release patterns during long-term decomposition. Science 315:361–4.
Powers JS, Montgomery RA, Adair EC, Brearley FQ, DeWalt SJ, Castanho CT, Chave J, Deinert E, Ganzhorn JU, Gilbert ME, et al. 2009. Decomposition in tropical forests: a pan-tropical study of the effects of litter type, litter placement and mesofaunal exclusion across a precipitation gradient. J Ecol 97:801–11.
Prescott CE. 2005. Do rates of litter decomposition tell us anything we really need to know? For Ecol Manag 220:66–74.
Preston CM, Nault JR, Trofymow JA. 2009. Chemical changes during 6 years of decomposition of 11 litters in some Canadian forest sites. Part 2. 13C abundance, solid-state 13C NMR spectroscopy and the meaning of ‘lignin’. Ecosystems 12:1078–102.
Rawlins AJ, Bull ID, Poirier N, Ineson P, Evershed R. 2006. The biochemical transformation of oak (Quercus robur) leaf litter consumed by the pill millipede (Glomeris marginata). Soil Biol Biochem 38:1063–76.
Rovira P, Vallejo VR. 1997. Organic carbon and nitrogen mineralization under Mediterranean climatic conditions: the effects of incubation depth. Soil Biol Biochem 29:1509–20.
Scheu S, Wolters V. 1991. Influence of fragmentation and bioturbation on the decomposition of 14C-labelled beech leaf litter. Soil Biol Biochem 23:1029–34.
Špaldoňová A, Frouz J. 2014. The role of Armadillidium vulgare (Isopoda: Oniscidea) in litter decomposition and soil organic matter stabilization. Appl Soil Ecol 83:186–92.
Suzuki Y, Grayston SJ, Prescott CE. 2013. Effects of leaf litter consumption by millipedes (Harpaphe haydeniana) on subsequent decomposition depends on litter type. Soil Biol Biochem 57:116–23.
Swift MJ, Heal OW, Anderson JM. 1979. Decomposition in terrestrial ecosystems. Berkeley: University of California Press.
Wall DH, Bradford MA, St John MG, Trofymow JA, Behan-pelletier V, Bignell DE, Dangerfield J, Parton WJ, Rusek J, Voigt W. 2008. Global decomposition experiment shows soil animal impacts on decomposition are climate-dependent. Glob Chang Biol 14:2661–77.
Webb DP. 1977. Regulation of deciduous forest litter decomposition by soil arthropod feces. In: Mattson WJ, Ed. The role of arthropods in forest ecosystems. New York: Springer. p 57–69.
Weishaar JL, Aiken GR, Bergamaschi BA, Fram MS, Fujii R, Mopper K. 2003. Evaluation of specific ultraviolet absorbance as an indicator of the chemical composition and reactivity of dissolved organic carbon. Environ Sci Technol 37:4702–8.
Wolters V. 2000. Invertebrate control of soil organic matter stability. Biol Fertil Soils 31:1–19.
Zimmer M, Pennings SC, Buck TL, Carefoot TH. 2002. Species-specific patterns of litter processing by terrestrial isopods (Isopoda: Oniscidea) in high intertidal salt marshes and coastal forests. Funct Ecol 16:596–607.
Acknowledgements
The chemical analyses were performed at the Plateforme d’Analyses Chimiques en Ecologie, a facility of the LabEx Centre Méditerranéen de l’Environnement et de la Biodiversité. We are grateful to Johanne Nahmani, Ammar Shihan and François-Xavier Joly for their help during field work and for helpful discussions. We thank our colleagues at the IMBE, Marseille, for their contributions, in particular Pascal Mirleau for 13C-RMN analyses and Mathieu Santonja and Natalia Rodriguez for the access to unpublished data. We also thank anonymous reviewers for their help to improve the quality of the manuscript. This research is part of the project CLIMED funded through the ANR Grant 09-CEP-007.
Author information
Authors and Affiliations
Corresponding author
Additional information
Author contribution
MC, JFD and SH designed the study. MC performed the research and analyzed data. MC, SC, JFD, and SH wrote the paper.
Rights and permissions
About this article

Cite this article
Coulis, M., Hättenschwiler, S., Coq, S. et al. Leaf Litter Consumption by Macroarthropods and Burial of their Faeces Enhance Decomposition in a Mediterranean Ecosystem. Ecosystems 19, 1104–1115 (2016). https://doi.org/10.1007/s10021-016-9990-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10021-016-9990-1