
Abstract
Extreme drought events challenge ecosystem functioning. Ecological response to drought is studied worldwide in a growing number of field experiments by rain-out shelters. Yet, few meta-analyses face severe challenges in the comparability of studies. This is partly because build-up of drought stress in rain-out shelters is modified by ambient weather conditions. Rain-out shelters can further create confounding effects (radiation, temperature), which may influence plant responses. Yet, a quantification of ecophysiological effects within rain-out shelters under opposing ambient weather conditions and of microclimatological artifacts is missing. Here, we examined phytometers—standardized potted individuals of Plantago lanceolata—under rain-out shelter, rain-out shelter artifact control, and ambient control during opposing outside microclimatological conditions. Furthermore, we tested for artifacts of rain-out shelters on plant responses in a long-term semi-natural grassland experiment. Phytometer plants below the rain-out shelters showed lower stomatal conductance, maximum quantum efficiency, and leaf water potential during warm ambient conditions with high evaporative demand than during cold conditions with low evaporative demand. Plant performance was highly correlated with ambient temperature and vapor pressure deficit (VPD). Rain-out shelter artifacts on plant responses were nonsignificant. Rain-out shelters remain a viable tool for studying ecosystem responses to drought. However, drought manipulations using rain-out shelters are strongly modified by ambient weather conditions. Attributing the results from rain-out shelter studies to drought effects and comparability among studies and study years therefore requires the quantification of the realized drought stress, for example, by relating ecosystem responses to measured microclimatological parameters such as air temperature and VPD.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Extreme climatic events are predicted to increase in frequency and magnitude (Min and others 2011; IPCC 2012) and are expected to have strong ecological implications (Jentsch and others 2007; Knapp and others 2008; Smith 2011; Reichstein and others 2013). Manipulation experiments in the field are powerful tools for the causal understanding of ecological processes in the face of climate change (Beier and others 2012; Smith and others 2014). Ecological response to climatic extremes such as drought events is therefore currently studied worldwide in a growing number of field experiments that predominantly use rain-out shelters (see Supporting information S1).
The quality of an experimental analysis, however, depends on whether the effective independent variables or environmental drivers are really considered (Hurlbert 1984) and on whether the experimental manipulations create hidden treatments (Huston 1997). Rain-out shelters are often criticized for unwanted side-effects on microclimatic conditions such as reduced wind speed, altered radiation, warming, and so on (Fay and others 2000; English and others 2005). Moreover, it can be expected that the manipulation strength of rain-out shelter experiments depends on outside weather conditions, as high air humidity during rainfall events should also affect the manipulated plots. Yet, detailed quantification of such microclimatological artifacts is largely missing and the majority of studies reporting on rain-out shelter effects either do not provide any information on recorded microclimate during drought manipulations (34% of all papers listed in Supporting information S1, n = 56), or measured air temperature (64%), whereas VPD or any other parameter that would reflect air humidity is rarely mentioned in any paper (9%).
To date, there has been one formal meta-analysis published on the results of precipitation manipulations (Wu and others 2011), and this analysis focused on reduced and increased total rainfall amounts without distinguishing between chronic partial rainfall reductions and pulsed full exclusion drought events or the timing and frequency of precipitation. Furthermore, it has recently been shown that soil moisture fluctuations under ambient reference conditions cannot successfully predict soil CO2 efflux in precipitation manipulations across numerous experimental datasets (Vicca and others 2014). Lack of comparability in drought definitions and response parameters among studies are major limiting factors for generalizations across ecosystems (Vicca and others 2012). However, we argue that comparability is further hampered by missing information on atmospheric conditions (that is, weather regimes) during drought manipulations. In general, effective differences between rain-out shelter manipulations and ambient control conditions can only be achieved during rainy periods. These conditions, however, are accompanied by high air humidity which can, for instance, cause opening of stomata (Lange and others 1971) also in the drought manipulation. Evidently, experimental drought manipulation through rain-out shelters can be strongly modified by ambient weather conditions. Effect sizes, however, are yet to be experimentally tested.
Generally, drought manipulations are realized in field experiments either as pulsed full exclusion (complete precipitation withdrawal for defined, rather short periods of time) or permanent partial exclusion (rainfall reduction typically by 30, 50, or 65% over longer times) manipulations. Up to now, the majority of published studies investigated temporally limited full exclusion drought events (42 cases in Table S1) in comparison to permanent partial exclusions (17 cases). Full exclusion drought manipulations are realized by fixed rain-out shelters (for example, Fay and others 2000; 26 cases in S1) or by mobile shelters which automatically close only during rainfall events (for example, Dugas and Upchurch 1984; 15 cases in S1). The latter might appear less sensitive for treatment artifacts, yet both techniques share the major challenge of trying to create drought when ambient conditions are humid. Partial exclusion is realized by partly covering the plots with translucent stripes that take away a defined percentage from any ambient rainfall event by a fixed setting (Yahdjian and Sala 2002) for rather long periods.
Fixed rain-out shelters in particular are criticized for unwanted side-effects such as increased air temperature, decreased solar radiation, wind, and vapor pressure deficit (Dugas and Upchurch 1984; Fay and others 2000; English and others 2005). In addition, rain-out shelters reduce turbulent exchange (that is, turbulent heat flux, transpiration, and carbon dioxide exchange), potentially resulting in increased levels of carbon dioxide under the shelters due to plant dark respiration. Besides this discussion, however, a crucial lack of evidence exists for the quantification of such artifacts on plant performance by the help of additional artifact controls (that is, plots irrigated with ambient precipitation below fully equipped rain-out shelters).
Here, we used two approaches. First, a standardized phytometer approach was applied to track short-term plant ecophysiological responses during standardized drought manipulation but opposing outside ambient weather conditions. Furthermore, we investigated community productivity under three climatic treatments (rain-out shelter manipulation, rain-out shelter artifact control, and ambient control) in an existing long-term climate manipulation experiment (EVENT II, Grant and others 2014) in a semi-natural grassland. We hypothesized that (1) drought stress in rain-out shelter experiments will build up faster during warm outside ambient weather conditions with high evaporative demand than during cold conditions with low evaporative demand. This implies that the realized drought effects are directly related to easily measurable meteorological parameters which will eventually allow for standardizations and comparability between sites and years. Furthermore, we expected that (2) rain-out shelter artifacts of fixed roofs on plant performance are considerable in their effect sizes.
Materials and Methods
Experimental Site
The present research was carried out in the Ecological-Botanical Garden of the University of Bayreuth, Germany (49°55′19″N, 11°34′55″E, 365 m a.s.l.). The short-term phytometer experiment in 2012 was conducted in the existing long-term field experiment Event II (Grant and others 2014). Potential long-term artifacts of rain-out shelters were tested using long-term productivity data from EVENT II. The EVENT II site was covered by a semi-natural grassland consisting of about 13 species m−2 and dominated by tall grasses, particularly Alopecurus pratensis L. (meadow foxtail). The regional climate is temperate and moderately continental, with a mean annual temperature of 8.0°C (1981–2010). The annual precipitation of 756 mm (1981– 2010) has a bimodal distribution with a major peak in June/July and a second peak in December/January (Lüers and others 2014). Following a spring drought in 2012, the precipitation during the measuring period (May to July) was comparable to the climatological mean (see Supporting information S2). Annual mean temperature and precipitation sum of 8.6°C and 698 mm, respectively, represent a moderately humid study year 2012 according to a De Martonne aridity index of 37.5.
Rain-out Shelter Manipulations
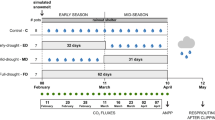
The climate treatments consisted of annually recurrent pulsed full exclusion drought manipulations and controls (treatment levels: rain-out shelter, rain-out shelter artifact control, and ambient control). The climate treatments were realized in a randomized block design with five replications for each treatment (see Figure 3 in Babel and others (2013) for a graphical representation of the experimental design). The ambient control plots remained without climate manipulation throughout the entire manipulation period and received ambient rainfall. Identical rain-out shelters were set up on rain-out shelter artifact controls and rain-out shelter plots during the climate manipulation for 42 days each year. Rain-out shelter plots remained without precipitation but rain-out shelter artifact plots received the same amount of precipitation as ambient control plots, added by irrigation from above the canopy after every rainfall. The shelters were constructed with a steel frame (Hochtunnel, E & R Stolte GmbH, Germany) and covered with a transparent plastic sheet (material: 0.2 mm polyethylene, SPR 5, Hermann Meyer KG, Germany, see Figure 1). Rain-out shelters permitted almost 90% penetration of Photosynthetically Active Radiation (PAR). Greenhouse effects due to rain-out shelters were minimized by having an 80-cm clearance between the lower roof edges and the ground, allowing for near-surface air exchange. Identical or very similar shelters have been used by us for several studies (Jentsch and Beierkuhnlein 2010) and also by other groups (for example, Kahmen and others 2005; Fay and others 2000). Lateral surface flow was avoided by barriers of plastic sheet pilings around all plots reaching down to a depth of 20–25 cm.

The EVENT II experiment with the applied rain-out shelters (photo: C. Schaller).
Phytometer Experiment
We used Plantago lanceolata as phytometer species. It is native to large parts of Eurasia, naturally common at the experimental site, and occurs as nonnative weed in, for example, North America, South America, and Australia (Seipel and others 2012). Moreover, this species has been used as model plant for stomatal conductance measurements by Clark and others (1999) and as phytometer by Temperton and others (2007). P. lanceolata was grown as temporal cohorts, so that they could be transferred to the field site at different dates, while being of identical age. Individuals of P. lanceolata were grown from seeds, using a standardized soil substrate: 20% washed sand, 20% fine lava (sand and lava-steamed at 90°C), and 60% mixture of white and black peat. The plants were germinated and grown in climate chambers under a diurnal rhythm of light (250 µmol m−2 s−1 PAR) at 20°C for 15 h and without light at 10°C for 9 h. All pots were watered on daily basis. After germination of seeds, we transplanted at the 20th day 30 single vigorous individuals into pots (9 × 9 × 9.5 cm3), using sandy loam as soil substrate (82% sand, 13% silt, 5% clay; pH 4.5 and total N = 0.07%). Pot size and soil substrate were selected according to pretrials with the aim of allowing for severe drought stress after seven days under warm and dry conditions. This was necessary to ensure that drought stress could potentially build up and be measured during the exposure of the phytometers. The first cohort of plants was sown on March 22nd, then over the next five weeks another five sets of plants were grown for the 2nd, 3rd, 4th, 5th, and 6th cohorts, each represented by 30 individuals. This sequential sowing ensured that each set of plants exposed to the experimental site was of the same age (62 days). Before exposure in the field, we soaked each pot in water for 2 min, to ensure the moisture balance being equal between pots. The first cohort of phytometers was taken out to the experimental site on 23rd of May, 2012 (see Supporting information S3 for detailed schedule). We used 10 phytometers for each treatment per temporal cohort in a nested block design with five pots placed on level ground in the center of two replications of each treatment block (those that were also covered by the microclimatological measurements; see Babel and others (2013) for full documentation of the spatial arrangements). Existing vegetation was cut to pot height and single pots placed with approximately 20 cm distance to each other.
In summary, contrasting outside ambient weather conditions during the experimental periods (23 May to 03 July, 2012) were evaluated by six temporal cohorts of phytometers, which were set under three climate manipulations (rain-out shelter, rain-out shelter artifact control, and ambient control). Each set of phytometers was exposed to the weather conditions in its respective climate treatments for 7 days.
Response Parameters in the Phytometer Experiment
Stomatal conductance, maximum quantum efficiency of photosystem II (ΔF/Fm), and leaf water potential are frequently studied response parameters relevant to tracking drought stress in plants. Stomatal conductance and ΔF/Fm can respond quickly within hours, whereas leaf water potential reacts more slowly, typically within days.
Stomatal conductance (mmol m−2 s−1) was measured during midday (12:00 to 2:00 pm, maximum stress) at the center of a medium-aged leaf from each plant using a steady-state leaf porometer (SC-1, Decagon Devices, Inc) at the 1st, 2nd, 3rd, and 7th day of exposure per cohort in the rain-out shelters (see Supporting information S3 for actual dates).
Dark-adapted chlorophyll fluorescence was measured at midnight (12:00 to 1:00 am) once per cohort (7th day) at the center of medium-aged leaves using a PAM 2000 portable chlorophyll Fluorometer (Heinz Walz, Germany). The maximum quantum efficiency of photosystem II was quantified as ΔF/F m = (F m − F t)/F m, where F t and F m are the actual and maximal chlorophyll fluorescence under ambient conditions, respectively (Genty and others 1989). ΔF/F m reflects the efficiency of light energy conversion of photosystem II (PS II) which is a sensitive parameter to quantify stress effect and photoinhibition in plants (Bolhar-Nordenkampf and others 1989; Werner and others 2002). Leaves were measured in situ in the natural position with a leaf clip holder.
Leaf water potential (MPa) was measured on the 7th day (maximum stress) with a PMS 600 pressure bomb (PMS Instrument Company, Albany, USA) using the Scholander pressure bomb technique. One medium-aged leaf from each plant was placed inside the closed chamber and pressurized.
Response Parameters in the Long-Term Field Experiments
We investigated annual net primary production (ANPP g m−2) and total cover (%) in ambient control versus rain-out shelter artifact control in the main field experiment (EVENT II, see Grant and others (2014) for details) over four consecutive years (2009–2012) to quantify long-term shelter artifacts.
Ambient Microclimatic Parameters Used to Explain the Drought Effects in Plant Performance
To investigate the relationship between ambient weather conditions outside the rain-out shelters and the degree of drought stress for the six temporal cohorts in the phytometer experiment, we investigated three easily measurable microclimatic parameters, that is, air temperature (°C), vapor pressure deficit (VPD, hPa), and incoming shortwave radiation (R sw, W m−2). Air temperature and humidity were measured by a Frankenberger Psychrometer (Friedrichs & Co., Germany) and radiation measurements (upwelling and downwelling longwave and shortwave components) with a CNR1 net radiometer (Kipp & Zonen, Netherlands). VPD was calculated from the difference between saturation water vapor pressure (hPa) and actual water vapor pressure (hPa). More details about these measurements are available in Babel and others (2013). In addition, soil moisture in the main rooting zone (between 5 and 10 cm depth) was recorded by FD sensors (Echo.EC-5/k; Decagon Devices, Pullman, WA, USA) in the long-term EVENT II experiment.
Data Analysis
Two-factorial linear mixed effects models combined with analysis of variance (ANOVA) were applied to test for significant differences between cohorts and treatments for all parameters in the phytometer experiment. The two fixed factors climate manipulation (rain-out shelter, rain-out shelter artifact control, and ambient control) and outside weather conditions (six cohorts) were tested for their single and interactive effects. The blocked spatial configuration of the experimental design was acknowledged in the mixed models by inserting the row and column of the treatment blocks as random effects. Prior to statistical analysis, data were power or log transformed to improve the homogeneity of variances, or if conditions of normality were not met. Both characteristics were tested by examining the residuals versus fitted plots and the normal qq plots of the linear models. Homogeneous groups of factor combinations were identified by Tukey’s HSD post hoc comparisons. Level of significance was set at p < 0.05.
Correlation analyses between ambient microclimatic parameters outside rain-out shelters and plant responses inside rain-out shelters were conducted in order to link the drought effects to various ambient weather conditions. We used three microclimatic parameters (air temperature, vapor pressure deficit, and incoming shortwave radiation) and three plant responses (stomatal conductance, ΔF/F m of photosystem II, and leaf water potential) for correlation analysis as these parameters were significantly different among the six cohorts, or at least there was a trend in the data (incoming shortwave radiation; Table 1).
Furthermore, we tested for long-term rain-out shelter artifacts on ANPP and cover responses by comparing ambient control and rain-out shelter artifact control over four consecutive years (2009–2012) in a semi-natural temperate grassland (EVENT II, Grant and others 2014). A linear mixed effect model combined with ANOVA was used to test for significant differences between years and treatments. Homogeneous groups of factor combinations were identified by Tukey’s HSD post hoc comparisons.
All analyses were run in R version 3.1.2 (R Core Team 2014) with the additional packages multcomp version 1.3-2 (function glht), nlme version 1.0-6 (function lme), and sciplot version 1.1-0 (function bargraph.CI).
Results
Ambient Weather Conditions Outside Rain-out Shelters Drive Drought Effect
The phytometers inside the rain-out shelters exhibited significantly reduced stomatal conductance (F = 42.2; p < 0.001), maximum quantum efficiency (F = 43.8; p < 0.001), and leaf water potential (F = 26.3; p < 0.001) during warm outside weather conditions with high evaporative demand as compared to those during cool conditions with low evaporative demand (Figure 2). Drought compared to ambient control significantly reduced plant stomatal conductance, ΔF/F m of photosystem II, and leaf water potential only when outside ambient weather conditions were warm and dry (Cohorts 4, 5, 6; Figure 3). The rain-out shelter manipulation did not significantly hamper the three plant response parameters compared to control in cool cohorts with low evaporative demand (in particular Cohort 2, same trend in Cohort 3; Figure 3). The clear pattern of drought effects under the rain-out shelters developing only under warm ambient weather conditions with high evaporative demand (Figure 2), however, is also confounded by the fact that significant treatment effects (the comparison between rain-out shelter manipulation and ambient control) further depend on the amount of precipitation taken away by the shelters. The pattern of drought treatment effects in Figure 3 therefore also depends on sufficient precipitation in the ambient controls to create any difference in moisture availability among the treatments. High temperatures combined with high evaporative demand cannot result in a significant difference among the rain-out shelter manipulation and the ambient control if there is no effective difference in precipitation, as clearly shown by cohort 1 (Figure 3).

Ecophysiological responses of standardized phytometer plants to exposure under rain-out shelters for seven days under different outside weather conditions (as summarized below the graph, daily mean of 30-min values during daytime). A Stomatal conductance (g s), B maximum quantum efficiency of photosystem II (ΔF/F m), and C leaf water potential (Ψ). Seventh day mean and standard error of 10 replicates per cohort are shown. Same letters indicate homogeneous groups according to Tukey’s HSD post hoc comparisons.

Relative differences in performance of standardized phytometer plants between ambient control and the rain-out shelter manipulation (∆d) after 7 days of exposure for A stomatal conductance (g s), B maximum quantum efficiency of photosystem II (ΔF/F m), and C leaf water potential (Ψ) for six temporal cohorts differing in outside weather conditions (as summarized below the graph, daily mean of 30-min values during daytime). Significant results (p < 0.05) of the corresponding two-factorial ANOVA with cohort, treatment, and their interaction as fixed effects are marked in bold and significant pairwise comparisons between ambient control and drought in each cohort according to Tukey’s HSD post hoc comparisons are indicated by asterisks. “Coh” = Cohort; “Treat” = Treatment. Daily means of air temperature (“Temp”), vapor pressure deficit (“VPD”), and sums of precipitation (“Prec”) are given per cohort.
The drought effects as observed in the ecophysiological plant responses were strongly correlated with the ambient microclimate if considered over several days (Table 2): Stomatal conductance was significantly correlated with the microclimate during phytometer exposure from the second day onwards, while it was not significantly related to any microclimatic parameter after exposure for only one day (Table 2). Likewise, maximum quantum efficiency of photosystem II (ΔF/F m) was significantly correlated with outside ambient microclimate conditions. Again, this relationship was stronger if considered over longer time spans (climate during the full cohort exposure versus climate of the final day of exposure only, Table 2). ΔF/F m and also leaf water potential were not significantly correlated with incoming shortwave radiation over the full cohort exposure. However, water potential was strongly related to the microclimate during the full exposure time. In general, multiple regressions, in particular those using temperature and VPD resulted in the strongest correlations across all response parameters (Table 2).
Rain-out Shelter Artifacts
Microclimatic parameters were considerably altered by the rain-out shelters with mean midday shortwave radiation being reduced by 22% and net radiation by 18% (Table 3). Mean summer temperatures were less strongly influenced. Still, mean daytime air temperature was reduced by 1.1°C, while nighttime temperature increased by 0.4°C (soil temperatures indicating comparable effects, Table 4). Soil moisture showed no consistent rain-out shelter artifact (slight increases or decreases in mean manipulation time soil moisture among years between artifact control and ambient control, Table 4). Minimum temperatures, however, were considerably higher under the shelters than in ambient control conditions (except midday, see Table 5).
Stomatal conductance, ΔF/F m, and leaf water potential in the rain-out shelter artifact treatment did not significantly differ from the ambient control (Figure 4). Furthermore, ANPP and aboveground plant cover of ambient control vs. rain-out shelter artifact control did not significantly differ and no significant interaction between treatment and year was observed in the long-term field experiment (EVENT II, Figure 5).

Relative differences in performance of standardized phytometer plants between ambient control and rain-out shelter artifact control (∆r) after 7 days of exposure for A stomatal conductance (g s), B maximum quantum efficiency of photosystem II (ΔF/F m), and C leaf water potential (Ψ) during five temporal cohorts differing in outside weather conditions (as summarized below the graph, daily mean of each 30-min values during daytime). Note that the sixth cohort is not displayed due to failed irrigation in the rain-out shelter artifact control. Significant results (p < 0.05) of the corresponding two-factorial ANOVA with cohort, treatment, and their interaction as fixed effects are marked in bold. Note that ambient control and rain-out shelter artifact control did not significantly differ in any response parameter (Treatment in ANOVA tables) and that furthermore no significant interaction between treatment and cohort was observed. “Coh” = Cohort; “Treat” = Treatment; “Temp” = Air temperature; “VPD” = Vapor pressure deficit.

Relative differences in plant responses between ambient control and rain-out shelter artifact control over four years (2009–2012) in a semi-natural temperate grassland (EVENT II, n = 5 except for ANPP responses in 2012 where n = 4). Significant results (p < 0.05) are marked in bold. Ambient daytime mean temperature (Temp) and vapor pressure deficit (VDP) over the experimental run (42 days during May to July each year) are given here.
Discussion
Ambient Weather Conditions Outside Rain-out Shelters Drive Drought Manipulation Effects
Drought stress inside rain-out shelters built up only during dry outside conditions with high evaporative demand, not during cold outside conditions with low evaporative demand in our experiment. This dependence of drought manipulation effects on ambient weather conditions implies that insights from different experimental sites and even from different years within the same experiment are hardly comparable, as they depend on uncontrollable weather patterns. Generalizations of the effects of full exclusion drought events just based on drought length of rain-out shelter studies are therefore elusive. Field-scale precipitation manipulation experiments remain, however, a crucial tool for the causal understanding of ecosystem responses to climate change (Beier and others 2012) and are applied in increasing numbers (Supporting information S1).
Temperature and VPD Scales’ Drought Effects on Plant Performance
The strong correlation between the realized ecophysiological drought effects and the ambient microclimate may offer options for generalization: At least for our experiment, drought effects can be standardized by microclimate during the drought manipulations and are then comparable among years. This relation needs to be tested under various climatic settings in other locations in order to test if it may offer a general option for facilitating comparability among drought manipulations. Up to now, the vast majority of rain-out shelter studies report no microclimatological characterization or air temperature only during their drought manipulations. VPD is rarely mentioned in any paper. VPD, which basically measures the dryness of the air, has been shown repeatedly to be closely correlated with plant productivity in systems ranging from tropical rainforests (Brando and others 2010) to maize monocultures (Kiniry and others 1998) or the understory of temperate forests (Leuschner 2002). Meta-analyses will benefit from the reporting of ambient weather conditions such as temperature and VPD as they need to explore responses to more complex and more realistic scenarios (Kreyling and Beier 2013) than simple annual reduction of rainfall (Wu and others 2011). Ultimately, observed ecological effects (or the absence thereof) can be attributed to the applied rain-out shelter treatment without any further characterization of microclimate. Interpreting such effects as drought effects, however, requires the confirmation (and quantification) of a substantial difference between drought manipulation and control that would then result in drought stress in plants. So we feel that the characterization of ambient environmental conditions during drought manipulations and their effect on the severity of a given treatment is not only an add-on which facilitates comparability across studies, but rather is a prerequisite for a sound interpretation of the results as drought effects (or the absence thereof).
The ecophysiological parameters measured in our short-term phytometer study capture the important aspects for ecosystem functioning; for example, transpiration and water potential have implications for water cycling and photosynthesis has implications for carbon cycling and productivity. Such ecophysiological data also represent fundamental inputs into many process-based ecosystem or biogeochemical models (for example, Hickler and others 2004), which are built for the purpose of scaling up to the response of ecosystems to environmental change. Differences in ecophysiology underlie any changes in ecosystem functioning as the ecosystem response is the result of the responses of all individual organisms (including their interactions). Based on this, our phytometer study was able to causally link ecophysiological responses to differences in ambient weather conditions by keeping all other conditions constant, such as the phenology and ontogeny of the individual plants and their potential rooting volume and available soil water. Therefore, we demonstrate the potential importance of ambient weather conditions for the interpretation of rain-out shelter manipulations on ecological functioning despite being unable to quantify the long-term effects on ecosystem functioning directly.
The effect of ambient weather conditions on plant performance under rain-out shelters in full precipitation exclusion experiments depends, of course, on water availability and therefore on soil water status. We favor quantifying easily measurable microclimatic parameters such as air temperature and VPD, in combination with recording rainfall, for comparisons among and within studies. Soil water status depends on a multitude of parameters (Hickler and others 2009) which can be assessed precisely only with considerable effort. Soil water status depends strongly on soil type, soil structure, soil depth, plant identity, and plant performance, just to name a few important interacting variables. All of these parameters are spatially highly heterogeneous and temporally highly dynamic (Bogner and others 2008; Glaser and others 2013). In particular, in the light of the current effort for coordinated distributed experiments with low maintenance effort and high replication across large spatial scales (Knapp and others 2012; Fraser and others 2013), such detailed quantification of soil water status is hardly feasible. We therefore recommend the quantification of ambient microclimate, which might even be taken a posteriori from nearby weather stations, in order to compare and generalize from a multitude of studies. Still, comprehensive interpretation of drought manipulations will require both the quantification of soil water availability and the quantification of microclimatological water demand because water stress will ultimately only occur when both conditions act together.
Here, we only considered a full exclusion pulsed drought manipulation experiment, that is, the complete withdrawal of precipitation over rather short periods of time (for example, Jentsch and others 2007, 2011). Yet, also the magnitude of manipulation effects in partial rainfall exclusions with rain-out shelters (Yahdjian and Sala 2002) depends strongly on outside conditions: during wet years, their effect may equal an average year, whereas during a dry year they create extreme conditions. Meaningful manipulation periods therefore require decades rather than single years in partial rainfall exclusion experiments (Kreyling and Beier 2013). Taken together, full and partial rainfall exclusions are not experimental alternatives, but rather different aspects of a larger picture. To enable comparability between full and partial rainfall exclusions and their effectiveness, we recommend precisely recording the ambient microclimate with adequate temporal (for example, daily) resolution.
Rain-out Shelter Artifacts on Plant Responses
We found no significant rain-out shelter artifacts either in short-term ecophysiological responses or in long-term productivity and plant cover when comparing artifact treatments with ambient conditions (Figures 4, 5) despite considerable effects on microclimate (Tables 3, 4, 5). For short-term full rainfall exclusion manipulations, this finding implies that fixed shelters can be used without creating strong artifacts with respect to the response parameters investigated in this study. Furthermore, the obvious microclimatological effects are in the direction of natural drought events, which are commonly accompanied by increased air temperatures (De Boeck and others 2010). It needs to be kept in mind, though, that the elimination of early-morning late frost events by the capture of longwave outgoing radiation might create a hidden treatment sensu Huston (1997). Such frost events in the early phase of the vegetation period can eventually cause drastic declines in grassland productivity (Kreyling and others 2012). As late frost did not occur during the presented experiment of this study, possible artifacts related to this cannot be evaluated here.
Movable shelters which automatically close only during rainfall events are a technically advanced solution which can reduce artifacts outside rainfall events (Beier and others 2004). Investment costs and the need for power supply, however, oppose regular application. Furthermore, these shelters usually close quite slowly and often after the onset of rainfall events and are not always functional during stormy conditions (Beier and others 2004). A complete withdrawal of water input is therefore hardly achieved, particularly in the face of short and intense rainfall events such as thunderstorms which typically carry most of the water input during growing seasons in temperate climates.
Partially covered rain-out shelters as commonly used for the manipulation of chronic rainfall reductions can be expected to cause similar, yet weaker microclimatic artifacts (Yahdjian and Sala 2002). The typical duration of manipulations for several years, however, raise concerns that even small microclimatic artifacts such as shading and warming might affect the ecological processes in the long run. Applying the same shelters with stripes turned upside down and therefore discharging into the shelters for control plots can circumvent these artifacts and allow for sound comparisons within studies. However, caution is required when the results are upscaled to real-world conditions in which dry periods are rather accompanied by increased incoming radiation than by shading (for example, Brando and others 2010).
We conclude that the effectiveness of drought simulations, that is, causing water stress, by rain-out shelters depends strongly on ambient weather conditions. Drought stress is only realized during warm and dry ambient atmospheric conditions. This finding limits the comparability of rain-out shelter studies in space and time. The close relation between drought stress and easily measurable microclimatic variables such as air temperature and vapor pressure deficit at our study site, however, promotes approaches to standardize studies and years, respectively. Yet, this relationship needs to be tested at other locations and climatic conditions.
Astonishingly, fixed rain-out shelters used for short-term complete rainfall exclusion were found to create less impact (artifact) on the plant responses than previously thought. Fixed shelters therefore remain a valid experimental tool and an alternative to sophisticated and costly automatic shelters powered by electric engines. Rain-out shelters are and will remain a major tool in ecological climate impact research. The precise measurement of microclimate and an improved understanding of microclimatic artifacts and hidden treatments will secure a sound interpretation of results and support comparability among studies, upscaling, and generalization.
References
Babel W, Schaller C, Eigenmann R, Foken T, Hübner J, Jentsch A, Kreyling J, Sultana F, Zhao P. 2013. Documentation of the EVENT-HMMS Experiment 2012—Microclimatological effects of rain-out shelters within EVENT II. Arbeitsergebnisse, Universität Bayreuth, Abteilung Mikrometeorologie, ISSN 1614-8916. 55:43 pp.
Beier C, Beierkuhnlein C, Wohlgemuth T, Penuelas J, Emmett B, Körner C, de Boeck H, Christensen JH, Leuzinger S, Janssens IA, Hansen K. 2012. Precipitation manipulation experiments—challenges and recommendations for the future. Ecol Lett 15:899–911.
Beier C, Emmett B, Gundersen P, Tietema A, Penuelas J, Estiarte M, Gordon C, Gorissen A, Llorens L, Roda F, Williams D. 2004. Novel approaches to study climate change effects on terrestrial ecosystems in the field: drought and passive nighttime warming. Ecosystems 7:583–97.
Bogner C, Wolf B, Schlather M, Huwe B. 2008. Analysing flow patterns from dye tracer experiments in a forest soil using extreme value statistics. Eur J Soil Sci 59:103–13.
Bolhar-Nordenkampf HR, Long SP, Baker NR, Oquist G, Schreiber U, Lechner EG. 1989. Chlorophyll fluorescence as a probe of the photosynthetic competence of leaves in the field—a review of current instrumentation. Funct Ecol 3:497–514.
Brando PM, Goetz SJ, Baccini A, Nepstad DC, Beck Pieter S A, Christman MC. 2010. Seasonal and interannual variability of climate and vegetation indices across the Amazon. PNAS 107:14685–90.
Clark H, Newton PC, Barker DJ. 1999. Physiological and morphological responses to elevated CO2 and a soil moisture deficit of temperate pasture species growing in an established plant community. J Exp Bot 50:233–42.
De Boeck HJ, Dreesen FE, Janssens IA, Nijs I. 2010. Climatic characteristics of heat waves and their simulation in plant experiments. Glob Change Biol 16:1992–2000.
Dugas WA, Upchurch DR. 1984. Microclimate of a rainfall shelter. Agron J 76:867–71.
English NB, Weltzin JF, Fravolini A, Thomas L, Williams DG. 2005. The influence of soil texture and vegetation on soil moisture under rainout shelters in a semi-desert grassland. J Arid Environ 63:324–43.
Fay PA, Carlisle JD, Knapp AK, Blair JM, Collins SL. 2000. Altering rainfall timing and quantity in a mesic grassland ecosystem: design and performance of rainfall manipulation shelters. Ecosystems 3:308–19.
Fraser LH, Henry HAL, Carlyle D, White S, Beierkuhnlein C, Cahill J, Casper BB, Cleland EE, Collins SL, Dukes JS, Knapp AK, Lind E, Long R, Smith MD, Sternberg M, Turkington R. 2013. Coordinated distributed experiments: an emerging tool for testing global hypotheses in ecology and environmental science. Front Ecol Environ 11:147–55.
Genty B, Briantais JM, Baker NR. 1989. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim Biophys Acta 990:87–92.
Glaser B, Jentsch A, Kreyling J, Beierkuhnlein C. 2013. Soil-moisture change caused by experimental extreme summer drought is similar to natural inter-annual variation in a loamy sand in Central Europe. J Plant Nutr Soil Sci 176:27–34.
Grant K, Kreyling J, Dienstbach LF, Beierkuhnlein C, Jentsch A. 2014. Water stress due to increased intra-annual precipitation variability reduced forage yield but raised forage quality of a temperate grassland. Agr Ecosys Environ 186:11–22.
Hickler T, Smith B, Sykes MT, Davis MB, Sugita S, Walker K. 2004. Using a generalized vegetation model to simulate vegetation dynamics in northeastern USA. Ecology 85:519–30.
Hickler T, Fronzek S, Araujo MB, Schweiger O, Thuiller W, Sykes MT. 2009. An ecosystem model-based estimate of changes in water availability differs from water proxies that are commonly used in species distribution models. Global Ecol Biogeogr 18:304–13.
Hurlbert SH. 1984. Pseudoreplication and the design of ecological field experiments. Ecol Monogr 54:178–211.
Huston MA. 1997. Hidden treatments in ecological experiments: re-evaluating the ecosystem function of biodiversity. Oecologia 110:449–60.
IPCC. 2012. Summary for Policymakers. In: Field CB, Barros V, Stocker TF et al., Eds. Managing the risks of extreme events and disasters to advance climate change adaptation: A Special Report of Working Groups I and II of the Intergovernmental Panel on Climate Change. Cambridge: Cambridge University Press. p 1–20.
Jentsch A, Beierkuhnlein C. 2010. Simulating the future—responses of ecosystems, key species and European provenances to expected climatic trends and events. Nova Acta Leopoldina 112:89–98.
Jentsch A, Kreyling J, Beierkuhnlein C. 2007. A new generation of climate change experiments: events, not trends. Front Ecol Environ 5:365–74.
Jentsch A, Kreyling J, Elmer M, Gellesch E, Glaser B, Grant K, Hein R, Lara M, Mirzae H, Nadler SE, Nagy L, Otieno D, Pritsch K, Rascher U, Schaedler M, Schloter M, Singh BK, Stadler J, Walter J, Wellstein C, Woellecke J, Beierkuhnlein C. 2011. Climate extremes initiate ecosystem-regulating functions while maintaining productivity. J Ecol 99:689–702.
Kahmen A, Perner J, Buchmann N. 2005. Diversity-dependent productivity in semi-natural grasslands following climate perturbations. Funct Ecol 19:594–601.
Kiniry JR, Landivar JA, Witt M, Gerik TJ, Cavero J, Wade LJ. 1998. Radiation-use efficiency response to vapor pressure deficit for maize and sorghum. Field Crops Res 56:265–70.
Knapp AK, Beier C, Briske DD, Classen AT, Luo Y, Reichstein M, Smith MD, Smith SD, Bell JE, Fay PA, Heisler JL, Leavitt SW, Sherry R, Smith B, Weng E. 2008. Consequences of more extreme precipitation regimes for terrestrial ecosystems. Bioscience 58:811–21.
Knapp AK, Smith MD, Hobbie SE, Collins SL, Fahey TJ, Hansen GJA, Landis DA, La Pierre KJ, Melillo JM, Seastedt TR, Shaver GR, Webster JR. 2012. Past, present, and future roles of long-term experiments in the LTER network. Bioscience 62:377–89.
Kreyling J, Beier C. 2013. Complexity in climate change manipulation experiments. Bioscience 63:763–7.
Kreyling J, Thiel D, Simmnacher K, Willner E, Jentsch A, Beierkuhnlein C. 2012. Geographic origin and past climatic experience influence the response to late spring frost in four common grass species in central Europe. Ecography 35:268–75.
Lange OL, Lösch R, Schulze ED, Kappen L. 1971. Responses of stomata to changes in humidity. Planta 100:76–86.
Leuschner C. 2002. Air humidity as an ecological factor for woodland herbs: leaf water status, nutrient uptake, leaf anatomy, and productivity of eight species grown at low or high vpd levels. Flora 197:262–74.
Lüers J, Soldner M, Olesch J, Foken T. 2014. 160 Jahre Bayreuther Klimazeitreihe, Homogenisierung der Bayreuther Lufttemperatur- und Niederschlagsdaten. Arbeitsergebnisse Universität Bayreuth, Abteilung Mikrometeorologie, ISSN 1614-8916. 56:52 pp.
Min S, Zhang X, Zwiers FW, Hegerl GC. 2011. Human contribution to more-intense precipitation extremes. Nature 470:378–81.
R Core Team. 2014. R: A Language and Environment for Statistical Computing. R version 3.1.2. Vienna, Austria: R Foundation for Statistical Computing. ISBN 3-900051-07-0, http://www.R-project.org.
Reichstein M, Bahn M, Ciais P, Frank D, Mahecha MD, Seneviratne SI, Zscheischler J, Beer C, Buchmann N, Frank DC, Papale D, Rammig A, Smith P, Thonicke K, van der Velde M, Vicca S, Walz A, Wattenbach M. 2013. Climate extremes and the carbon cycle. Nature 500:287–95.
Seipel T, Kueffer C, Rew LJ, Daehler CC, Pauchard A, Naylor BJ, Alexander JM, Edwards PJ, Parks CG, Arevalo JR, Cavieres LA, Dietz H, Jakobs G, McDougall K, Otto R, Walsh N. 2012. Processes at multiple scales affect richness and similarity of non-native plant species in mountains around the world. Global Ecol Biogeogr 21:236–46.
Smith MD. 2011. An ecological perspective on extreme climatic events: a synthetic definition and framework to guide future research. J Ecol 99:656–63.
Smith MD, Rodgers VL, Brzostek E, Kulmatiski A, Avolio ML, Koerner SE, Hoover D, Grant K, Jentsch A, Fatichi S, Niyogi D. 2014. Towards a better integration of biogeochemical and ecological data from precipitation manipulation experiments into land surface models. Rev Geophys. doi:10.1002/2014RG000458.
Temperton VM, Mwangi PN, Scherer-Lorenzen M, Schmid B, Buchmann N. 2007. Positive interactions between nitrogen-fixing legumes and four different neighbouring species in a biodiversity experiment. Oecologia 151:190–205.
Vicca S, Bahn M, Estiarte M, van Loon EE, Vargas R, Alberti G, Ambus P, Arain MA, Beier C, Bentley LP, Borken W, Buchmann N, Collins SL, de Dato G, Dukes JS, Escolar C, Fay P, Guidolotti G, Hanson PJ, Kahmen A, Kroel-Dulay G, Ladreiter-Knauss T, Larsen KS, Lellei-Kovacs E, Lebrija-Trejos E, Maestre FT, Marhan S, Marshall M, Meir P, Miao Y, Muhr J, Niklaus PA, Ogaya R, Penuelas J, Poll C, Rustad LE, Savage K, Schindlbacher A, Schmidt IK, Smith AR, Sotta ED, Suseela V, Tietema A, van Gestel N, van Straaten O, Wan S, Weber U, Janssens IA. 2014. Can current moisture responses predict soil CO2 efflux under altered precipitation regimes? A synthesis of manipulation experiments. Biogeosciences 11:2991–3013.
Vicca S, Gilgen AK, Serrano MC, Dreesen FE, Dukes JS, Estiarte M, Gray SB, Guidolotti G, Hoeppner SS, Leakey ADB, Ogaya R, Ort DR, Ostrogovic MZ, Rambal S, Sardans J, Schmitt M, Siebers M, van der Linden L, van Straaten O, Granier A. 2012. Urgent need for a common metric to make precipitation manipulation experiments comparable. New Phytol 195:518–22.
Werner C, Correia O, Beyschlag W. 2002. Characteristic patterns of chronic and dynamic photoinhibition of different functional groups in a Mediterranean ecosystem. Funct Plant Biol 29:999–1011.
Wu Z, Dijkstra P, Koch GW, Penuelas J, Hungate BA. 2011. Responses of terrestrial ecosystems to temperature and precipitation change: a meta-analysis of experimental manipulation. Glob Change Biol 17:927–42.
Yahdjian L, Sala OE. 2002. A rainout shelter design for intercepting different amounts of rainfall. Oecologia 133:95–101.
Acknowledgments
Mohammed A.S. Arfin Khan and F. Sultana were financially supported by the German Academic Exchange Service (DAAD) while at the Department of Disturbance Ecology, University of Bayreuth. The financial support for the set-up of the EVENT II experiment was provided by a research grant of the German Science Foundation (DFG JE 282/6-1). We thank the Ecological-Botanical Gardens for providing the required experimental sites.
Author information
Authors and Affiliations
Corresponding author
Additional information
Author Contributions
AJ, JK, TF, and CB designed the study; FS, JK, and JW performed research; MAK, JK, and WB analyzed the data; JK and MAK wrote the paper with substantial input from all the authors.
Juergen Kreyling and Mohammed A. S. Arfin Khan have contributed equally to this work.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Kreyling, J., Arfin Khan, M.A.S., Sultana, F. et al. Drought Effects in Climate Change Manipulation Experiments: Quantifying the Influence of Ambient Weather Conditions and Rain-out Shelter Artifacts. Ecosystems 20, 301–315 (2017). https://doi.org/10.1007/s10021-016-0025-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10021-016-0025-8