
Abstract
In grassland ecosystems, where soil water most frequently controls ecosystem processes, expected changes in precipitation and temperature may have dramatic effects on ecosystem dynamics. Previous observational studies have reported that aboveground net primary production (ANPP) in grasslands is very sensitive to changes in precipitation. Yet, we lack experimentally based evidence to support these observations. Further, most of the studies have focused solely on ANPP, neglecting belowground production (BNPP). This is an important gap in our knowledge, as BNPP represents 50% or more of total net primary production (NPP) in grasslands. Here, we present results from a 3-year water manipulation experiment (2008–2010) at two sites in the central grassland region of North America, USA. We were successful in changing the soil water content in our treatments, but these changes resulted in different, but significant responses in ANPP and BNPP at our two sites. At the shortgrass steppe, we found that neither NPP nor ANPP were sensitive to treatment precipitation, and although we found BNPP was sensitive to changes in treatment precipitation, the direction of the response varied between years. In contrast, ANPP was very sensitive to treatment precipitation on the mixed-grass prairie, whereas BNPP was insensitive. Based on our finding that two grassland ecosystems showed dramatically different above and belowground production responses to soil water manipulations, we cannot assume that predicted changes in climate will cause similar above- and belowground production responses. Second, our results demonstrated that sites within the same region may differ markedly in the sensitivity of ANPP to changes in growing season precipitation.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Human activities since the industrial revolution have contributed to regional and global changes in climate, including an increase in global temperature and changes in precipitation patterns (Christensen and others 2007). Global change will not affect the Earth uniformly—temperature and precipitation changes will vary across regions, and responses will depend on an ecosystem’s sensitivity to climate parameters (Knapp and Smith 2001). Thus, it is important to understand how different ecosystems may respond to future climatic perturbations. Predicting the effects of global change in grasslands is especially important, as they represent the potential natural vegetation of over 40% of the Earth’s terrestrial surface (White and others 2000) and include many productive agricultural and rangeland areas. The central grassland region of North America is an ideal ecosystem for studying the effects of global change on grassland communities. The region represents 12.5% of North America and 2% of the Earth’s terrestrial surface (Lauenroth and others 1999). Both temperature and precipitation vary along the north–south and east–west gradients of the region, and this range of climates within the region allows for comparisons with a variety of grasslands worldwide.
Climate models predict that the central grassland region of North America will warm substantially (3–4.4°C) during the twenty-first century (Christensen and others 2007; CCSP 2008). Associated predictions of precipitation changes are less certain and range from small increases to small decreases. It is still not clear how warmer temperatures and associated changes in precipitation regimes will alter ecosystem soil water patterns. However, ecosystems in the region are most frequently water limited (Noy-Meir 1973; Sala and others 1988), and whether the predicted effects of climate changes increase or decrease soil water, any change in soil water will likely have dramatic effects on ecosystem dynamics (Weltzin and others 2003).
A critical ecosystem property that is sensitive to soil water availability is net primary production (NPP), the net amount of carbon removed from the atmosphere by plants. Plant production can be divided into aboveground net primary production (ANPP) and belowground net primary production (BNPP). Quantifying ANPP is a frequent goal of basic and applied ecology (Sala and Austin 2000), as estimates of ANPP are necessary to understand the global carbon balance and trophic interactions. From an applied perspective, estimates of ANPP are used to determine forage availability and stocking rates for livestock and managed wildlife populations in rangelands, and wood yield in forests. Observational studies have found that ANPP is tightly linked to precipitation across the central grassland region (Sala and others 1988), and through time with interannual variability at individual sites (Lauenroth and Sala 1992; Briggs and Knapp 1995). This provides good reason to expect that grasslands will be responsive to predicted future changes in soil water. Indeed, recent predictions of changes in precipitation patterns and temperature have inspired field experiments to examine the effects of altered precipitation on grassland community structure and function with striking results (Grime and others 2000; Fay and others 2003; Zavaleta and others 2003; Yahdjian and Sala 2006; Heisler-White and others 2009; Jentsch and others 2011).
Although predicted changes in soil water will likely have an important impact on ANPP, there is a paucity of studies examining the effect of altered soil water on BNPP. Furthermore, we lack a regional understanding of soil water availability and its impacts on BNPP in the central grassland region (McCulley and others 2005). Yet BNPP is an important portion of total production: it contributes 50% or more to the total NPP of grassland ecosystems (Sims and others 1978; Milchunas and Lauenroth 2001), and represents a large terrestrial carbon sink (Scurlock and Hall 1998). We need to understand how NPP, not just the aboveground component, will respond to changes in soil water to improve models and predictions of future carbon storage and turnover in grassland ecosystems.
Several studies have demonstrated that grasslands are sensitive to variation in precipitation, a surrogate for soil water. In a study spanning a variety of ecosystem types in North America, Knapp and Smith (2001) reported that aboveground production in grassland and old field ecosystems was most sensitive to precipitation variability. In an earlier observational study, Weaver and Albertson (1936) reported large and rapid changes in the species composition and total plant cover at sites in the eastern and central portions of the central grassland region in response to the drought of the 1930s, noting the largest vegetation changes in areas dominated by tallgrass species. More recently, Paruelo and others (1999) found that precipitation use efficiency across the region was maximized at intermediate precipitation values (462 or 491 mm, depending on the data used), not the minimum or maximum represented by the shortgrass steppe or tallgrass prairie. This contrast in aboveground ecosystem response is likely due to differences in plant species traits across the region. In the driest portion of the central grassland region, the dominant species have low relative growth rates and are unable to respond quickly to interannual variation in precipitation. In the wettest portion of the region, the dominant species have higher relative growth rates and can respond more quickly to interannual variation in precipitation, but an increase in biomass or leaf area index will cause limitation by other resources; namely light or nutrients (Knapp and Seastedt 1986; Baer and others 2003). These studies provide observational evidence of differences in sensitivity within grassland types to variations in soil water, yet we lack experimentally based evidence to support these observations. Further, these studies focus solely on aboveground production responses, and lack descriptions of belowground production responses.
Here, we aim to understand how changes in soil water will influence grassland ecosystem function in the driest portions of the central grassland region. We conducted a 3-year water manipulation experiment at two study sites to examine the effects of altered soil water dynamics on NPP. Specifically, we experimentally increased and decreased soil water to determine if there was a difference in the sensitivity of NPP (both the above and belowground components) to changes in soil water between sites. Further, we aimed to determine if above and belowground production respond similarly to manipulations of soil water within and across sites. Based on previous studies, we hypothesized that NPP at the shortgrass steppe site would be less sensitive to changes in soil water than NPP at the mixed-grass prairie site. In addition, we predicted that the belowground production response would mirror the aboveground production response.
Materials and Methods
Study Sites
We conducted this study at two grassland sites located within the central grassland region, which spans a 550-km transect from eastern Colorado to central Kansas. Although the entire region is characterized by a continental climate and similar soil types, strong climatic gradients, including a west to east increasing precipitation gradient and north to south temperature gradient exist. These differences in temperature and precipitation lead to distinct plant communities at different locations within the region. Despite these differences, the grasslands are often lumped together in climate change modeling.
Our first site is located within the semiarid shortgrass steppe at the Central Plains Experimental Range (CPER), 60 km northeast of Fort Collins, Colorado (40°49′N, 104°46′W). The CPER is administered by the USDA Agriculture Research Service and was a National Science Foundation Long Term Ecological Research site. Mean annual precipitation is 341 mm and mean annual temperature is 8.2°C. These climatic conditions lead to a plant community that is dominated by the short-stature C4 grasses. Blue grama (Bouteloua gracilis Willd. ex Kunth Lag. ex Griffiths), which typically accounts for 80–90% of total plant cover in this ecosystem (Milchunas and others 1989) is the dominant species and patches of bare ground are prevalent. The soils are Aridic Argiustolls.
In contrast, the second site is located in the subhumid mixed-grass prairie, further east and south along the climatic gradient of the central grassland region. The site is located at the Fort Hays State University College Farm, in west-central Kansas, 3 km west of Hays (38°52′N, 99°23′W). Mean annual precipitation is 583 mm and mean annual temperature is 12.1°C. The differences in climate result in a very different plant community from that of the shortgrass steppe. The plant community includes a mixture of tall-, mid-, and short-stature C4 grasses, but the clear dominant species at the site is the tall-stature grass little bluestem (Schizachyrium scoparium (Michx.) Nash.). The soils are Typic Argiustolls, with underlying fractured limestone bedrock. Cattle grazing is excluded at both study sites. Species nomenclature follows USDA Plants Database (USDA 2011).
Experimental Protocol
Shortgrass Steppe
In May, 2008, we selected 30 plot locations with similar abundances of the dominant species, B. gracilis, and associated subdominant species. We randomly assigned a treatment (drought or ambient) to each 1.0 m2 plot. We imposed drought by creating 15 1.8 m long × 1.6 m wide rainfall shelters designed to decrease incoming ambient rainfall by 60% (Yahdjian and Sala 2002). The roofs had a 15° inclination with the short side of the shelters oriented west; the dominant direction of incoming rainstorms. The shorter side of the shelters were 0.6 m tall, whereas the taller sides were 1.1 m tall, thus the shelter was always at least about 0.5 m above maximum vegetation height. The roofs were made of 15 cm wide strips of corrugated polycarbonate which transmit greater than 90% PAR (Dynaglass brand). The corrugated strips channel rainfall into gutters that lead water away from the plots. Shelter extended an additional 0.2 m beyond the plot in each direction to help reduce the amount of rain flowing horizontally into the plots from outside, and shelter sides were open to maximize air movement and minimize potential temperature and relative humidity artifacts. To examine the effects of altered growing season precipitation, drought shelters were present on plots during the May–September growing season, and were taken down during the dormant period. We removed any dust or bird droppings from the shelters before reassembling them each May.
In May 2009, we selected 15 additional plots with similar species composition to the existing plots and began a water addition treatment. Throughout the growing season (May–September) we calculated the weekly long term average rainfall for the site, and added the difference between the observed rainfall and double the weekly long term average rainfall once weekly by hand using watering cans. During the 2009 growing season this amounted to an additional 92 mm of water added, and 179 mm in 2010, which is an average increase in precipitation of 71% over mean annual precipitation.
Mixed-Grass Prairie
In late March, 2008, we selected two blocks of nine plots, each block separated by 0.5 km. We randomly assigned each plot to one of three precipitation treatments: drought, ambient, and water addition. Each plot was 2.0 × 8.0 m2; long sides oriented with the slope. The shortest shelter height was approximately 1.5 m above ground level. We constructed six 4.0 × 10.0 m2 rainfall shelters that were designed to intercept 50% of incoming ambient rainfall. Similar to the shortgrass steppe site, a pitched roof of 15 cm wide strips of corrugated polycarbonate (Dynaglass brand) channeled rainfall into gutters and removed water from the plots. Each shelter extended 1.0 m beyond the plots in each direction to help reduce the amount of rain flowing horizontally into the plots from outside, and like the shelters at the shortgrass steppe, shelter sides at this site were open. Drought shelters were permanent and left up year-round. Although it is possible that the differences in experimental set up between sites (growing season rainfall shelters at the shortgrass steppe and year-round rainfall shelters at the mixed-grass prairie) may have influenced our results, the sites both receive the majority of annual precipitation during the summer growing season, when shelters were up at both sites.
We applied water with a pump system from a 5,680-l holding tank connected to a network of drip lines. Once a week throughout the growing season (April–September) we applied the long-term average weekly precipitation, regardless of the actual rainfall received during the week, totaling an addition of 380 mm of water during the growing season each year, which is an average increase in precipitation of 61% over mean annual precipitation. This approach insured a wetter than normal treatment, even if ambient precipitation was below average.
Soil Water
We installed Decagon Devices EC-5 soil water probes in March 2008 at the mixed-grass prairie site at a depth of 5 cm in a subset of plots for each treatment (n = 2 for control, n = 5 for drought, n = 6 for water addition; Decagon Devices Inc., Pullman, WA, USA). In June 2009, we installed ECH2O soil water probes at the shortgrass steppe site at a depth of 10 cm in a subset of plots for each treatment (n = 5 for control, drought, and water addition treatments; Decagon Devices Inc., Pullman, WA, USA). All probes were installed perpendicular to the soil surface and remained in place for the duration of the experiment at each site. Soil water probes measured volumetric soil water content (θ v) every 4 h at each site.
We sampled soil in all plots and found no significant textural differences between treatments at each site (data not shown), so we used the mean absolute difference in soil θ v to quantify how our treatments altered soil θ v during the growing season at each site. We determined these values using daily soil θ v in the control (n = 5 at shortgrass steppe and n = 2 at mixed-grass prairie), drought (n = 5 at shortgrass steppe and n = 5 at mixed-grass prairie), and water addition (n = 5 at shortgrass steppe and n = 6 at mixed-grass prairie) treatments.
Aboveground Net Primary Production
We estimated ANPP nondestructively at both sites to minimize plot disturbance. We used methods that have been proven to be effective in semiarid and subhumid grasslands (Byrne and others 2011; Frank and McNaughton 1990; Paruelo and others 2000; Przeszlowska and others 2009). Because we were interested in within-site differences among treatments and years, we used the most accurate yet least destructive method for measuring ANPP for each site. Therefore our methods differed between sites and additionally, among years at the shortgrass steppe. Thus, the focus of our analysis is on relative differences among treatments within years, not absolute differences between sites or among years. We measured ANPP consistently across plots within years, and because there were no large changes in species composition across time (unpublished data), we feel confident in our abilities to detect treatment differences in ANPP using our non-destructive methods. These non-destructive measurements worked well to preserve plot integrity over our 3-year experiment, but we caution the use of non-destructive estimates in situations where species composition in treatment plots differ from calibration plots where ANPP is destructively harvested.
We estimated ANPP nondestructively on the shortgrass steppe in 2008 using green cover estimates derived from a digital camera (Byrne and others 2011). This method produced variable results (r 2 = 0.23, P < 0.05), however, our 2008 estimates were well within the range of clipped ANPP estimates recorded in 2008 in nearby pastures (SGS LTER, unpublished data). In 2009 and 2010, we estimated ANPP on the shortgrass steppe using both a point-frame and a radiometer technique (Byrne and others 2011). As we were more interested in comparing differences in ANPP within sites than making absolute comparisons between sites, we report estimates of ANPP using the more accurate point-frame technique for the shortgrass steppe site (R 2 = 0.91 and 0.90 for 2009 and 2010, respectively).We used a 62 × 80 cm (0.5 m2) quadrat with 50 equally spaced points inside to estimate cover by functional groups, and also included average vegetation height into our estimate of ANPP (Byrne and others 2011).
At the mixed-grass prairie site, we used a radiometer to nondestructively estimate ANPP for each year of the experiment (Byrne and others 2011). We measured reflectance between 11:00 and 14:00 hours on clear days, at a height of 1.25 m above each plot, which the manufacturer calculated as the distance above the ground necessary to achieve a 0.25-m2 circular sampling area. We took reflectance measurements at eight randomly selected locations within each treatment plot and averaged these measurements to calculate plot-level ANPP estimates.
We performed each nondestructive method on every treatment plot (one location per plot on the shortgrass steppe; eight locations per plot on the mixed-grass prairie) at peak biomass (late July in the mixed-grass prairie; early August on the shortgrass steppe). We selected 15 separate calibration plots in which we performed the nondestructive techniques, then clipped green and recent dead material of grasses and forbs and current year’s green production (excluding woody tissue) for dwarf-shrubs. We clipped biomass at the soil surface, separated by functional group, and placed in paper bags. We dried samples for at least 48 h at 55°C then weighed them. We used total biomass data as an estimate of ANPP (Lauenroth and others 1986; Lauenroth and others 2006). Each year we fit a regression relating the nondestructive methods to our estimate of ANPP (see Byrne and others 2011), and converted our estimates to g m−2 in all cases.
Belowground Net Primary Production
We obtained estimates of BNPP using a modified root ingrowth technique (Vogt and others 1998; McCulley and others 2005; Milchunas 2009). In early May of each year, we excavated 5-cm diameter cores to a depth of 15 cm at the shortgrass steppe and 12 cm at the mixed-grass prairie (due to fractured limestone beginning at 12 cm). We note that this method is not perfect, as roots certainly extend beneath the top 12 or 15 cm of the soil profile, and we missed any changes in BNPP that possibly occurred at depths below our measurements. However, previous work in the shortgrass steppe has shown that over 35% of roots occur in the top 0–10 cm (compared to 10–40 cm) (Milchunas and others 2005b), and Frank and others (2010) reported an exponential decline in root biomass with increasing depth. Worldwide, grasses have an average of 44% of total standing root biomass in the top 10 cm (Jackson and others 1996). Although our method is an underestimate of total belowground production, it captures a large proportion of total root production. One core was excavated in each plot on the shortgrass steppe and five cores were excavated in each plot at the mixed-grass prairie. We placed a mesh cylinder of the same dimensions with 1.0 mm square openings within the excavated core, as 1.0 mm mesh allows the largest diameter roots to pass through (Milchunas and others 2005a; LeCain and others 2006). Excavated cores were refilled with twice sieved root-free soil from the site. In October of each year, ingrowth cores were removed by driving an 8 cm diameter soil corer into the soil surrounding the ingrowth core. Roots and soil on the outside of the mesh cylinders were brushed away, and cores were placed in paper bags and air-dried to minimize decomposition before processing. Once root cores dried, we manually separated out the largest roots (>2 cm), and used a hydropneumatic elutriation system to separate fine roots from the soil. The hydropneumatic elutriator used a combination of pressurized air and water to gently separate roots from soil, then allowed water to wash over the roots and clean them. Once all root samples were cleaned and separated, we dried roots for 48 h at 55°C and weighed samples. We then combusted each sample in a muffle furnace at 500°C for 5 h to determine ash content. We calculated BNPP from the values obtained from the total root sample weight and report BNPP in g m−2 y−1 on an ash-free basis.
Data Analysis
We performed a residual analysis on each year’s estimated ANPP and BNPP for each plot in each treatment and removed any data points that fell more than three SD from the mean. To assess if our treatments had a different effect on ANPP and BNPP at each site between years, we used a mixed model analysis of variance in the Statistical Analysis System (v 9.2, Cary, NC, USA). The fixed effects in the model were year, treatment, and the interaction between those two terms. We included year as a fixed effect to account for any differences between treatments not accounted for by treatment. We included plot (and block for the mixed-grass prairie site) as a random effect. In cases where the treatment effect was significant, we tested for differences among treatments using a Tukey adjustment for multiple comparisons.
To assess if there was a linear relationship between estimated precipitation (mm) received in each treatment and each response variable (ANPP, BNPP, and NPP), we replaced the categorical variable “treatment” from our mixed model analysis of variance above with the continuous variable, estimated precipitation received in each treatment (henceforth “treatment precipitation”). At each site and for each response variable (ANPP, BNPP, and NPP) we created models using the fixed effects that were significant for each site and response variable. The year × treatment precipitation interaction term was only significant for BNPP and NPP at the shortgrass steppe, so the interaction term was only included in the final model for those variables. Values presented are means ± 1 SE and the level of significance for all statistical tests is P < 0.05, unless otherwise noted.
Results
Temperature and Precipitation
The 50-year (1959–2009) mean annual temperature at the shortgrass steppe is 8.2°C, and annual temperature ranged from 7.5 to 8.3°C during the 3-year experiment. The 50-year mean annual temperature at the mixed-grass prairie is 12.1°C, and annual temperature ranged from 11.7 to 12.9°C during the 3-year experiment. Annual precipitation, growing season precipitation, and monthly precipitation were much more variable than temperature throughout the experiment at both sites. Annual precipitation ranged from 330 to 436 mm at the shortgrass steppe, whereas the long term mean precipitation is 341 mm. Long-term mean growing season precipitation (May–September) is 251 mm and ranged from 176 to 282 mm during the experiment (Figure 1A). At the mixed-grass prairie site, the long-term mean annual precipitation is 582 mm. During the experiment, annual precipitation ranged from 552 to 727 mm. Long-term mean growing season (April–September) precipitation is 431 mm, and ranged from 441 to 495 mm during the experiment (Figure 1B).

A April and growing season (May–September) precipitation received at the shortgrass steppe, and B growing season (April–September) precipitation received at the mixed grass prairie, during the experiment years (2008–2010) and long-term monthly growing season precipitation. In individual experiment years, annual precipitation was 331, 436, and 330 mm at the shortgrass steppe and 727, 552, and 600 mm at the mixed-grass prairie in 2008–2010, respectively. Growing season precipitation was 247, 282, and 176 mm at the shortgrass steppe (May–September) and 495, 441, and 489 mm at the mixed-grass prairie (April–September) in 2008–2010, respectively.
Soil Water (θ v)
Shortgrass Steppe
The drought and water addition treatments strongly influenced growing season soil water content. Treatment differences were smallest at the beginning of the growing season, but increased in July and August as soils dried in the drought and control treatments whereas soil water was replenished by weekly water addition in the water addition treatment (Table 1). During the 2009 growing season (soil water probes were not installed until 16 June 2009) in the upper 10 cm of the soil profile, the drought treatment reduced mean soil water content by 24% whereas the water addition treatment increased mean soil water content by 15%. Over the 2010 growing season (1 May–30 Sept.), the drought treatment-reduced mean soil water content by 17% whereas the water addition treatment increased mean soil water content by 13.5%.
Mixed-Grass Prairie
Like the shortgrass steppe site, the imposed water manipulation treatments influenced growing season soil water content, and the largest treatment differences occurred during the months of June–August, whereas the smallest treatment differences occurred toward the beginning and end of the growing season, when temperatures were relatively cooler (Table 2). During the 2008 growing season (1 April–30 Sept), the drought treatment reduced mean soil water content by 37% and the water addition treatment increased mean soil water content by 49%. Over the 2009 growing season, the drought treatment-reduced mean soil water content by 18% and the water addition treatment increased mean soil water content by 82%. Over the 2010 growing season, the drought treatment-reduced mean soil water content by 13% and the water addition treatment increased mean soil water content by 88%.
Shortgrass Steppe Net Primary Production
ANPP
Year, but not treatment, was a factor for ANPP at the shortgrass steppe site (supplementary Table 1). Within years, ANPP was similar among treatments. Only in 2010 was there a difference between treatments, and here the drought treatment had higher ANPP than both the control and water addition treatments. There was not a significant linear relationship between treatment precipitation and ANPP (Table 3). Within each treatment, ANPP was higher in 2009 and 2010 than in 2008, but ANPP was similar between 2009 and 2010, except for the irrigation treatment, which had lower ANPP in 2010 than in 2009 (Figure 2A).

A Shortgrass steppe NPP, and B mixed-grass prairie NPP, in drought, control, and water addition treatments. Uppercase letters represent differences within a given treatment between years, and lowercase letters represent differences between treatments within a given year. Differences significant at P < 0.05, but * indicates significance at P < 0.10. Bars represent standard error. Differences in ANPP are shown above the NPP bars, differences in BNPP are shown below the NPP bars, and differences in NPP are shown within the NPP bars.
BNPP
BNPP at the shortgrass steppe was affected by treatment, year, and the year × treatment interaction (supplementary Table 1). There was no consistent trend in BNPP differences between treatments within years (Figure 2A). In the first treatment year (2008), the drought and control treatments had very similar values for BNPP. In 2009 both the drought and control treatments had higher BNPP than the water addition treatment. In contrast, in 2010 the drought treatment had lower BNPP than the water addition treatment, and the drought treatment had 39% lower BNPP than the control treatment (P = 0.106). There was not a significant linear relationship between treatment precipitation and BNPP (Table 3). In the control and drought treatments, BNPP was higher in 2009 than in 2008 or 2010, but there was no difference in BNPP for the water addition treatment between years.
NPP
Total NPP at the shortgrass steppe was affected by treatment, year, and the year × treatment interaction (supplementary Table 1). The water addition treatment had lower NPP than both the drought and control treatments in 2009, but in 2010 there were no differences in NPP between treatments. Although there was a linear relationship between treatment precipitation and NPP, the slope was very low (Table 3). Within each treatment, there was a difference in NPP between years for the drought and control treatments for all 3 years, but no difference in NPP for the water addition treatment. NPP for the drought and control treatments was remarkably similar throughout the experiment (Figure 2A).
Mixed-Grass Prairie Net Primary Production
ANPP
Treatment and year were factors for ANPP at the mixed-grass prairie site (supplementary Table 1). Throughout the experiment, there was a trend toward reduced ANPP in the drought treatment and increased ANPP in the water addition treatment, with the control treatment falling in the middle. The difference was significant between the drought and water addition treatments for all 3 years, but the difference was never significant between the drought and control treatments. There was a positive linear relationship between treatment precipitation and ANPP (Table 3). Among years, all three treatments had higher ANPP in 2009 and 2010 than in 2008, but only in the water addition treatment was there a difference in ANPP between 2009 and 2010 (Figure 2B).
BNPP
Treatment and year were factors for BNPP at the mixed-grass prairie site (supplementary Table 1). The only differences between treatments occurred in the final treatment year (2010). Here, the control had higher BNPP than both the drought and water addition treatments (Figure 2B). There was not a significant linear relationship between treatment precipitation and BNPP (Table 3). Likewise, there was no difference in BNPP among years for any treatment, except for the control, which had higher BNPP in 2010 than in 2009 (Figure 2B).
NPP
Similar to above and belowground production, treatment, and year were factors for NPP at the mixed-grass prairie site (supplementary Table 1). The differences within years among treatments were similar to the results for ANPP. In 2008 and 2009 the water addition treatment had higher NPP than both the control and drought treatments. In 2010 both the control and water addition treatments had higher NPP than the drought treatment, but the NPP values for the control and water addition treatments were similar (Figure 2B). There was a significant linear relationship between treatment precipitation and NPP (Table 3). There was no difference in NPP among years for the drought treatment. The control treatment had higher NPP in 2009 and 2010 than in 2008, and the water addition treatment had higher NPP in 2009 than in 2008 (Figure 2B).
Discussion
Climate models forecast warming and changes in precipitation in the central grassland region of North America during the twenty-first century (Christensen and others 2007; CCSP 2008). Such predictions represent novel conditions for the region and the effects of these changes remain largely unknown. Our results show that the two grasslands were differentially sensitive to changes in soil water, and that above and belowground responses to our experimental manipulation were contrasting.
Sensitivity of Net Primary Production to Changes in Soil Water Across Sites: ANPP
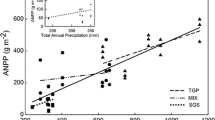
Previous studies have established a tight link between precipitation and ANPP in grassland regions (Rosenzweig 1968; Sala and others 1988; Knapp and Smith 2001). Our ANPP results from the shortgrass steppe were not consistent with this previous research. We found that NPP was not sensitive to changes in treatment precipitation, and ANPP only showed a minimal (P < 0.1) treatment response in the drought treatment in the final treatment year (2010). Although BNPP was responsive to changes in treatment precipitation, the direction of the response was variable among years (Figure 2A; Table 3). In contrast to the shortgrass steppe, our NPP results from the mixed-grass prairie were congruent with previous research. We found a strong-linear relationship between estimated treatment precipitation and both ANPP and NPP, but not BNPP (Figure 2B; Table 3).
An examination of the ecohydrology of the region can partially explain the lack of observable response in NPP at the shortgrass steppe. Potential evapotranspiration (PET) exceeds precipitation by a factor of 3 during the growing season, and the most common state of the top 45 cm of the soil is dry (Lauenroth and Bradford 2006). Dry soil during the growing season is the result of pulse precipitation followed by rapid drying (Sala and Lauenroth 1982). The rapid cycling of soil water conditions between wet and dry is not conducive to plant production. Noy-Meir (1973) suggested that “an effective rain event” is one that activates biological processes (in particular, production and reproduction). Apparently, our water additions in the shortgrass steppe were not large enough to change soil water content long enough to stimulate production. This explanation is consistent with the results of Lauenroth and others (1978) and Heisler-White and others (2008). Both reported an increase in ANPP in response to increased soil water. Yet Lauenroth and others (1978) added much more water than we did: an average of 228 mm of water per growing season whereas we added roughly half as much, an average of 135 mm per growing season. Although Heisler-White and others (2008) did not increase growing season precipitation per se, they increased soil water by manipulating rainfall event size and frequency. These two studies demonstrate that if enough water is added during the growing season, the shortgrass steppe is responsive, yet the amount required to elicit a response in ANPP exceeds current predictions by general circulation models of climate change.
Although our study focused on changes in soil water during the growing season, perhaps future efforts would be better focused on predicted changes in soil water during the dormant period when PET is low at the shortgrass steppe. In an analysis of long-term production at the CPER, Milchunas and others (1994) found that production was very sensitive to changes in cool season (Oct–April) precipitation, but only slightly sensitive to changes in growing season precipitation. Another recent study of net ecosystem production at the CPER concluded that additional precipitation during the July–Oct period would have much less of an impact on net ecosystem production than additional precipitation received during the April–June time period (Parton and others 2011). In addition, a rainfall manipulation experiment in the northern mixed-grass prairie found that severe spring drought (May–June) reduced ANPP, whereas water addition in the summer (July–August) did not significantly increase ANPP compared to a control treatment (Heitschmidt and Vermeire 2006).
In contrast to the semiarid shortgrass steppe, subhumid grasslands experience relatively high soil water availability for a large portion of the growing season, and growing season rainfall events are more likely to maintain ecosystem processes such as photosynthesis in an unstressed state (Knapp and others 1993). Indeed, studies have shown that soil water is typically not the most limiting resource in the tallgrass prairie (Briggs and Knapp 1995). Although the mixed-grass prairie does not receive as much precipitation as the tallgrass prairie, we can estimate the atmospheric demand for water (MAPET) and the ratio of mean annual precipitation (MAP) to MAPET as an indication of the adequacy of the water supply to meet the atmospheric demand for each site using the formula from Lauenroth and Burke (1995);
where MAPET and MAP are measured in cm and MAT (mean annual temperature) is measured in °C. A value at or above 1 indicates that the water supply does adequately meet the atmospheric demand for water. The estimated value at the shortgrass steppe is 0.25, whereas the value at the mixed-grass prairie is 50% higher; 0.38. Although precipitation at the mixed-grass prairie does not completely satisfy the atmospheric demand for water, less additional water is needed to ameliorate water stress and results in an “effective rainfall event” (Noy-Meir 1973). Our irrigation treatment clearly was able to stimulate production at this more mesic site.
The differing sensitivities of ANPP to changes in soil water at our two sites may also be partly explained by vegetational constraints. The shortgrass steppe is dominated by drought resistant species, especially the dominant grass species, B. gracilis (Hyder 1975; Mueller and Weaver 1942). Bouteloua gracilis is adapted to persist below ground in extended periods of low soil water, although the plant’s aboveground leaves may wither and die. The bunchgrass’s lack of rhizomes and stolons make it difficult for individual genets or tillers to respond quickly to favorable water conditions (Mueller 1941). It is likely that B. gracilis and other species had limited responses to the surplus or decrease of soil water in the treatments during our 3-year experiment. A longer study may be needed to see consistent ecosystem responses. Indeed, another study on the shortgrass steppe found that there were no significant changes in total cover in a 50% drought treatment for the first 4 years of a drought manipulation (Evans and others 2011), and other studies in semiarid grasslands have found little reduction in ANPP due to drought (Heitschmidt and others 1999; Cipriotti and others 2008).
In contrast, the vegetation at the mixed-grass prairie is dominated by grass species with a range of traits. These species can adjust total cover or leaf area index faster than the drought tolerant shortgrasses and were therefore able to respond faster to changes in soil water. Previous research by Weaver (1954) during and after the great drought of the 1930s supports these findings: he recorded much greater changes in the plant communities of the tall and mixed-grass prairies as compared to the shortgrass steppe.
Sensitivity of Net Primary Production to Changes in Soil Water Across Sites: BNPP
Our trends in ANPP were consistent with previous soil water manipulation research (Lauenroth and others 1978; Heisler-White and others 2008; Heisler-White and others 2009) and predictions regarding the sensitivity of the central grassland region to variability in precipitation (Paruelo and others 1999; Knapp and Smith 2001), but trends in BNPP were not. Although the between-year patterns were variable for the shortgrass steppe, BNPP responded significantly to soil water manipulation in the second- and third-treatment years (Figure 2A). In contrast, BNPP at the mixed-grass prairie was insensitive to changes in soil water except for the third treatment year, although there was a trend toward reduced BNPP in the drought treatment and increased BNPP in the water addition treatment (Figure 2B).
A few other studies have examined the effect of soil water manipulation on BNPP in grassland ecosystems. In a European grassland experiment, Fiala and others (2009) found that BNPP was reduced with drought and increased with water addition. Two rainfall experiments in Inner Mongolia also reported an increase in BNPP due to water addition, but the studies had no corresponding drought manipulation (Bai and others 2010; Gao and others 2011). In a study spanning the central grassland region, McCulley and others (2005) found that BNPP was virtually identical at the shortgrass steppe in two different rainfall years, whereas BNPP at the mixed-grass prairie was greater in the year with higher growing season rainfall. Although we could not determine a pattern in BNPP response to soil water manipulation at the shortgrass steppe, it is clear that BNPP did exhibit sensitivity to changes in soil water. This result corresponds with previous soil water manipulation studies, whereas our BNPP results at the mixed-grass prairie contradict experiments reporting rapid and significant belowground responses to altered precipitation (Fiala and others 2009; Bai and others 2010; Gao and others 2011). This discrepancy points to the need for additional long-term manipulations to gain a better understanding of both above and belowground plant production responses to predicted changes in climate and soil water.
Are ANPP and BNPP Responses Similar Within and Across Sites?
To our knowledge, ours is the first multiple-year study that examines the response of both ANPP and BNPP in grasslands to both an increase and decrease in soil water across multiple sites. At the shortgrass steppe, we generally found that BNPP, but not ANPP, was sensitive to changes in soil water (Figure 2A). The opposite was true at the more mesic mixed-grass prairie, where ANPP and total NPP were sensitive to changes in soil water, whereas BNPP generally was not (Figure 2B; Table 3). The results of our study indicate that we cannot assume that belowground production will mirror aboveground production in response to predicted climate changes, and highlights the importance of including measurements of belowground net primary production and total net primary production in future climate change experiments and models. Considering the importance of BNPP in contributing to total plant carbon inputs in grassland ecosystems, ignoring grassland BNPP may lead to erroneous predictions if our assumptions about belowground responses to changes in precipitation are incorrect. Additional experimental studies across multiple sites are required to broaden our understanding of the controls of BNPP in grassland ecosystems.
Implications
Our results have several important implications for future climate change research and for predicting the magnitude of temperate grassland responses to these expected climate changes. First, it is clear that we must be careful when we employ previous assumptions about net primary production in our efforts to predict responses to future predicted changes in soil water. We found that two ecosystems within the same grassland biome showed different above and belowground production responses to soil water manipulations. Second, our results indicate that there may be differences in the periods of ecosystem sensitivity among ecosystems within the same biome. The shortgrass steppe was relatively insensitive, whereas the mixed-grass prairie was clearly sensitive to changes in growing season precipitation. Finally, the lack of consistency in our results among years and between sites highlights the need for more multi-site, long-term studies. Given the agricultural importance of grasslands within this region and worldwide, the complexity of both within site and across biome NPP responses to predicted climate changes warrant future experiments to improve our understanding of ecosystem dynamics within the grassland biome.
References
Baer SG, Blair JM, Collins SL, Knapp AK. 2003. Soil resources regulate productivity and diversity in newly established tallgrass prairie. Ecology 84:724–35.
Bai W, Wan S, Niu S et al. 2010. Increased temperature and precipitation interact to affect root production, mortality, and turnover in a temperate steppe: implications for ecosystem C cycling. Glob Chang Biol 16:1306–16.
Briggs JM, Knapp AK. 1995. Interannual variability in primary production in tallgrass prairie: climate, soil moisture, topographic position, and fire as determinants of aboveground biomass. Am J Botany 82:1024–30.
Byrne KM, Lauenroth WK, Adler PB, Byrne CM. 2011. Estimating aboveground net primary production in grasslands: a comparison of nondestructive methods. Rangel Ecol Manage 64:498–505.
CCSP. 2008. The effects of climate change on agriculture, land resources, water resources, and biodiversity. A report by the US Climate Change Science Program and the Subcommittee on Global Change Research. Washington, DC: US Environmental Protection Agency.
Christensen JH, Hewitson B, Busuioc A et al. 2007. Regional climate projections. In: Solomon S, Qin D, Manning M et al., Eds. Climate Change 2007: The physical science basis. Contribution of working Group I to the fourth assessment report of the intergovernmental panel on climate change. Cambridge, UK/New York, NY, USA: Cambridge University Press.
Cipriotti PA, Flombaum P, Sala OE, Aguiar MR. 2008. Does drought control emergence and survival of grass seedlings in semi-arid rangelands? An example with a Patagonian species. J Arid Environ 72:162–74.
Evans SE, Byrne KM, Lauenroth WK, Burke IC. 2011. Defining the limit to resistance in a drought-tolerant grassland: long-term severe drought significantly reduces the dominant species and increases ruderals. J Ecol 99:1500–7.
Fay PA, Carlisle JD, Knapp AK, Blair JM, Collins SL. 2003. Productivity responses to altered rainfall patterns in a C4-dominated grassland. Oecologia 137:245–51.
Fiala K, Tůma I, Holub P. 2009. Effect of manipulated rainfall on root production and plant belowground dry mass of different grassland ecosystems. Ecosystems 12:906–14.
Frank DA, McNaughton SJ. 1990. Aboveground biomass estimation with the canopy intercept method: a plant growth form caveat. Oikos 57:57–60.
Frank DA, Pontes AW, Maine EM, Caruana J, Raina R, Raina S, Fridley JD. 2010. Grassland root communities: species distributions and how they are linked to aboveground abundance. Ecology 91:3201–9.
Gao Y, Chen Q, Lin S, Giese M, Brueck H. 2011. Resource manipulation effects on net primary production, biomass allocation and rain-use efficiency of two semiarid grassland sites in Inner Mongolia, China. Oecologia 165:855–64.
Grime JP, Brown VK, Thompson K et al. 2000. The response of two contrasting limestone grasslands to simulated climate change. Science 289:762–5.
Heisler-White J, Knapp A, Kelly E. 2008. Increasing precipitation event size increases aboveground net primary productivity in a semi-arid grassland. Oecologia 158:129–40.
Heisler-White JL, Blair JM, Kelly EF, Harmoney K, Knapp AK. 2009. Contingent productivity responses to more extreme rainfall regimes across a grassland biome. Glob Chang Biol 15:2894–904.
Heitschmidt RK, Vermeire LT. 2006. Can abundant summer precipitation counter losses in herbage production caused by spring drought? Rangel Ecol Manage 59:392–9.
Heitschmidt RK, Haferkamp MR, Karl MG, Hild AL. 1999. Drought and grazing: I. Effects on quantity of forage produced. J Range Manage 52:440–6.
Hyder DN. 1975. Ecological responses of native plants and guidelines for management of shortgrass range. United States Department of Agriculture Technical Bulletin No. 1503.
Jackson RB, Canadell J, Ehleringer JR, Mooney HA, Sala OE, Schulze ED. 1996. A global analysis of root distributions for terrestrial biomes. Oecologia 108:389–411.
Jentsch A, Kreyling J, Elmer M et al. 2011. Climate extremes initiate ecosystem-regulating functions while maintaining productivity. J Ecol 99:689–702.
Knapp AK, Seastedt TR. 1986. Detritus accumulation limits productivity of tallgrass prairie. Bioscience 36:662–8.
Knapp AK, Smith MD. 2001. Variation among biomes in temporal dynamics of aboveground primary production. Science 291:481–4.
Knapp AK, Fahnestock JT, Hamburg SP, Statland LB, Seastedt TR, Schimel DS. 1993. Landscape patterns in soil–plant water relations and primary production in tallgrass prairie. Ecology 74:549–60.
Lauenroth W, Bradford J. 2006. Ecohydrology and the partitioning AET between transpiration and evaporation in a semiarid steppe. Ecosystems 9:756–67.
Lauenroth WK, Burke IC. 1995. Great plains, climate variability. In: Nierenberg WA, Ed. Encyclopedia of environmental biology. San Diego, CA: Academic Press. p 237–49.
Lauenroth WK, Sala OE. 1992. Long-term forage production of North American shortgrass steppe. Ecol Appl 2:397–403.
Lauenroth WK, Dodd JL, Sims PL. 1978. The effects of water- and nitrogen-induced stresses on plant community structure in a semiarid grassland. Oecologia 36:211–22.
Lauenroth WK, Hunt HW, Swift DM, Singh JS. 1986. Estimating aboveground net primary production in grasslands: a simulation approach. Ecol Model 33:297–314.
Lauenroth WK, Burke IC, Gutmann MP. 1999. The structure and function of ecosystems in the central North American grassland region. Gt Plains Res 9:223–59.
Lauenroth W, Wade A, Williamson M, Ross B, Kumar S, Cariveau D. 2006. Uncertainty in calculations of net primary production for grasslands. Ecosystems 9:843–51.
LeCain D, Morgan J, Milchunas D, Mosier A, Nelson J, Smith D. 2006. Root biomass of individual species, and root size characteristics after five years of CO2 enrichment on native shortgrass steppe. Plant Soil 279:219–28.
McCulley RL, Burke IC, Nelson JA, Lauenroth WK, Knapp AK, Kelly EF. 2005. Regional patterns in carbon cycling across the great plains of North America. Ecosystems 8:106–21.
Milchunas D. 2009. Estimating root production: comparison of 11 methods in shortgrass steppe and review of biases. Ecosystems 12:1381–402.
Milchunas DG, Lauenroth WK. 2001. Belowground primary production by carbon isotope decay and long-term root biomass dynamics. Ecosystems 4:139–50.
Milchunas DG, Forwood JR, Lauenroth WK. 1994. Productivity of long-term grazing treatments in response to seasonal precipitation. J Range Manage 47:133–9.
Milchunas DG, Morgan JA, Mosier AR, Lecain DR. 2005a. Root dynamics and demography in shortgrass steppe under elevated CO2, and comments on minirhizotron methodology. Glob Chang Biol 11:1837–55.
Milchunas DG, Mosier AR, Morgan JA, Lecain DR, King JY, Nelson JA. 2005b. Root production and tissue quality in a shortgrass steppe exposed to elevated CO2: using a new ingrowth method. Plant Soil 268:111–22.
Mueller IM. 1941. An experimental study of rhizomes of certain prairie plants. Ecol Monogr 11:165–88.
Mueller IM, Weaver JE. 1942. Relative drought resistance of seedlings of dominant prairie grasses. Ecology 23:387–98.
Noy-Meir I. 1973. Desert ecosystems: environment and producers. Ann Rev Ecol Syst 4:25–51.
Parton W, Morgan J, Smith D, Del Grosso S, Prihodko L, LeCain D, Kelly R, Lutz S. 2011. Impact of precipitation dynamics on net ecosystem productivity. Glob Chang Biol. doi:10.1111/j.1365-2486.2011.02611.x.
Paruelo JM, Lauenroth WK, Burke IC, Sala OE. 1999. Grassland precipitation: use efficiency varies across a resource gradient. Ecosystems 2:64–8.
Paruelo JM, Lauenroth WK, Roset PA. 2000. Estimating aboveground plant biomass using a photographic technique. J Range Manage 53:190–3.
Przeszlowska A, Trlica MJ, Weltz MA. 2009. Near-ground remote sensing of green area index on the shortgrass prairie. Rangel Ecol Manage 59:422–30.
Rosenzweig ML. 1968. Net primary productivity of terrestrial communities: prediction from climatological data. Am Nat 102:67–74.
Sala OE, Austin AT. 2000. Methods of estimating aboveground net primary productivity. In: Sala OE, Jackson RB, Mooney HA, Howarth RW, Eds. Methods in ecosystem science. New York: Springer-Verlag. p 31–43.
Sala OE, Lauenroth WK. 1982. Small rainfall events: an ecological role in semiarid regions. Oecologia 53:301–4.
Sala OE, Parton WJ, Joyce LA, Lauenroth WK. 1988. Primary production of the central grassland region of the United States. Ecology 69:40–5.
Scurlock JMO, Hall DO. 1998. The global carbon sink: a grassland perspective. Glob Chang Biol 4:229–33.
Sims PL, Singh JS, Lauenroth WK. 1978. The structure and function of ten western North American grasslands: I. abiotic and vegetational characteristics. J Ecol 66:251–85.
USDA, NRCS. 2011. The PLANTS Database (http://plants.usda.gov). National Plant Data Team, Greensboro, NC 27401-4901 USA.
Vogt KA, Vogt DJ, Bloomfield J. 1998. Analysis of some direct and indirect methods for estimating root biomass and production of forests at an ecosystem level. Plant Soil 200:71–89.
Weaver JE. 1954. North American Prairie. Lincoln, NE, USA: Johnsen Publishing Company.
Weaver JE, Albertson FW. 1936. Effects on the great drought on the prairies of Iowa, Nebraska, and Kansas. Ecology 17:567–639.
Weltzin JF, Loik ME, Schwinning S, Williams DG, Fay PA, Haddad BM, Harte J, Huxman TE, Knapp AK, Lin G, Pockman WT, Shaw MR, Small EE, Smith MD, Smith SD, Tissue DT, Zak JC. 2003. Assessing the response of terrestrial ecosystems to potential changes in precipitation. Bioscience 53:941–52.
White RP, Murray S, Rohweder M, Prince SD, Thompson KMJ. 2000. Grassland ecosystems. Washington, DC: World Resources Institute.
Yahdjian L, Sala O. 2002. A rainout shelter design for intercepting different amounts of rainfall. Oecologia 133:95–101.
Yahdjian L, Sala OE. 2006. Vegetation structure constrains primary production response to water availability in the Patagonian steppe. Ecology 87:952–62.
Zavaleta ES, Shaw MR, Chiariello NR, Thomas BD, Cleland EE, Field CB, Mooney HA. 2003. Grassland responses to three years of elevated temperature, CO2, precipitation, and N deposition. Ecol Monogr 73:585–604.
Acknowledgments
This study was conducted in part at the Central Plains Experimental Range (CPER), which is administered by the USDA Agricultural Research Service (ARS) and was a Long Term Ecological Research site (SGS-LTER), funded by the National Science Foundation (Grant No. 1027319). This research was supported in part by the Utah Agricultural Experiment Station (to PBA), Utah State University, and approved as journal paper number 8356. The Nature Conservancy provided partial support for this work through the Nebraska Chapter’s J.E. Weaver Competitive Grants Program (to KMB). We thank J. A. Klein, A. K. Knapp, and two anonymous reviewers for their constructive comments and suggestions on the manuscript. The authors thank J.A. Leiker, M. Lindquist, and the Shortgrass Steppe Field Crew for their assistance designing and constructing the rainout shelters, conducting fieldwork, and applying water to the water addition treatment.
Author information
Authors and Affiliations
Corresponding author
Additional information
Author Contributions
KMB, PBA, and WKL conceived and designed the experiments. KMB and PBA performed the fieldwork. KMB analyzed the data and wrote the manuscript with editorial advice from WKL and PBA.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Byrne, K.M., Lauenroth, W.K. & Adler, P.B. Contrasting Effects of Precipitation Manipulations on Production in Two Sites within the Central Grassland Region, USA. Ecosystems 16, 1039–1051 (2013). https://doi.org/10.1007/s10021-013-9666-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10021-013-9666-z