
Abstract
The observed long-term decrease in the regional fire activity of Eastern Canada results in excessive accumulation of organic layer on the forest floor of coniferous forests, which may affect climate–growth relationships in canopy trees. To test this hypothesis, we related tree-ring chronologies of black spruce (Picea mariana (Mill.) B.S.P.) to soil organic layer (SOL) depth at the stand scale in the lowland forests of Quebec’s Clay Belt. Late-winter and early-spring temperatures and temperature at the end of the previous year’s growing season were the major monthly level environmental controls of spruce growth. The effect of SOL on climate–growth relationships was moderate and reversed the association between tree growth and summer aridity from a negative to a positive relationship: trees growing on thin organic layers were thus negatively affected by drought, whereas it was the opposite for sites with deep (>20–30 cm) organic layers. This indicates the development of wetter conditions on sites with thicker SOL. Deep SOL were also associated with an increased frequency of negative growth anomalies (pointer years) in tree-ring chronologies. Our results emphasize the presence of nonlinear growth responses to SOL accumulation, suggesting 20–30 cm as a provisional threshold with respect to the effects of SOL on the climate–growth relationship. Given the current climatic conditions characterized by generally low-fire activity and a trend toward accumulation of SOL, the importance of SOL effects in the black spruce ecosystem is expected to increase in the future.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Boreal forests play a significant role in the global carbon cycle because they store about 25% of the terrestrial carbon stocks (Apps and others 1993; Dixon and others 1994). Canadian boreal forests represent about 25% of global boreal forests and account for almost 90% of productive forest area in Canada (Tarnocai 2006). In turn, black spruce (Picea mariana (Mill.) B.S.P.) forests hold a dominant position in the boreal forest of North America (Viereck and Johnston 1990) and are estimated to have the largest total ecosystem carbon content, as compared to other main forest ecosystems within the biome (Gower and others 1997). In this context, understanding climatic controls upon growth of dominant species and future successional trajectories of black spruce forests is crucial for both adequate modeling of ecosystem response to future climatic variability and its resilience under natural and human-related changes in environmental conditions.
Climate change directly affects growth conditions of trees in the boreal zone (Apps and others 1993; Amiro and others 2001) by modifying environmental constraints on their physiological activity, including soil water and temperature regimes, decomposition rates, nutrient cycling (Allison and Treseder 2008), and competitive interactions among trees (Bonn 2000; Johnstone and Chapin 2006). Climatically driven changes in the natural disturbance regimes, for example, in the regional fire cycle (Bergeron and others 2004a), also affect tree growth in the boreal biome by controlling soil organic layer (SOL) thickness, which in turn affects root zone conditions (van Cleve and others 1990; Simard and others 2007). At the millennium time scale, the accumulation of SOL in the boreal forest has been taking place since the last glaciation (Ritchie 2004). At a shorter (centennial) time scale, the SOL accumulation rate is directly affected by climatic variability, whereas the rate of SOL removal from the system is mostly controlled by the natural fire cycle and the amount of SOL removed during single fire events (that is, fire severity) (Conard and others 2002).
The successional development toward paludified forests, reported over large areas of boreal North America, Europe, and Siberia (Crawford and others 2003), calls for a better understanding of the effects of SOL accumulation on tree growth dynamics. Understanding this relationship is critical to quantify both direct and indirect effects of climate change on forest biomass production, carbon sequestration, and ecosystem stability at the regional and global scales (Kurbatova and others 2008; Smith and others 2008). At the stand and regional scales, this knowledge would improve modeling of future stand productivity and its spatio-temporal dynamics (Komarov and Kubasova 2007; Peltoniemi and others 2007), including better parameterization of large-scale carbon models such as Biome-BGC (Kimball and others 1997; Ueyama and others 2009). Although changes in stand productivity along gradients of SOL have been reported earlier (Prescott and others 2000; Simard and others 2007), little is known about annual variation in growth as affected by differences in thickness of the organic horizon. This applies to growth dynamics during both normal and extreme weather conditions.
In the Clay Belt region of Eastern Canada, natural succession in black spruce stands is typically initiated by stand-replacing fires and ultimately proceeds toward dominance of moist black spruce-Sphagnum communities (Lecomte and others 2005; Fenton and Bergeron 2007). The decrease in fire activity since the middle of the nineteenth century reported for the region (Bergeron and others 2001) and, possibly, an increase in growing season precipitation (Archambault and Bergeron 1992), both facilitate the accumulation of SOL. SOL accumulation has been shown to result in expansion of Sphagna, and ultimately, in forest paludification, that is, the successional development toward forested peatlands (Sirén 1955; Fenton and others 2005; Lecomte and others 2006).
It has also been suggested that even without clear signs of paludification, accumulation of organic layer may have negative effects on seed establishment and growth of tree canopy dominants (Greene and others 2007). In black spruce forests, thickening of SOL is associated with the development of adventitious roots (LeBarron 1945; Islam and MacDonald 2004), resulting from a decline in the function of deeper roots. As a result, an increase in SOL depth may affect soil temperature and the availability of nutrients and water (van Cleve and others 1983a), and mediate the trees’ response to annual weather variation (Dang and Lieffers 1989a, b). Influence of SOL on spruce growth may, therefore, be partitioned into (a) effects related to a decrease in the amount of available nutrients along a gradient of increasing SOL, and (b) effects related to modification of physical conditions under which the roots function. In the former case, the effect is due to decreased cationic exchange capacity and available nitrogen and phosphorus in the organic layers compared to the mineral horizons (van Cleve and others 1983a; van Cleve and others 1983b; Simard and others 2007). In the latter case, the effect of SOL is due to changes in temperature and aeration regimes of the root zone (Silins and Rothwell 1999; Laiho 2006). Ultimately, both effects are interrelated because they affect trees’ ability to produce biomass. As no repeated measurements of physical environment or nutrient availability are commonly available, it is difficult to provide conclusive answers about the relative roles of these effects. Analysis of spruce growth variability along a SOL gradient could be the first step in quantifying the role of SOL in black spruce ecosystems.
In this study, we analyzed the effects of SOL thickness on climate–growth relationships in black spruce forests located between 78.4° and 79.7° W, and 48.1° and 50.1° N in the Clay Belt region of Northern Ontario and Quebec. In this part of the eastern North American boreal forest, dendrochronological reconstructions of the regional fire regime (Bergeron and others 2004b) suggest a decrease in fire activity over the last century, facilitating landscape-level accumulation of SOL (Simard and others 2007). By analyzing the effects of SOL on climate–growth relationships at one spatial scale (stand) and two temporal scales (annual versus decadal variation), we formulate the following objectives: (1) identify monthly weather variables that control black spruce growth and (2) evaluate how SOL thickness affects the relationship between weather and spruce growth. The network of sites used in this study covered a wide range of soil conditions with the organic layer depth varying from 0 to 51.7 cm and with the mean tree age in stands ranging from 35 to 250 years. By focusing on the impact of organic layer depth, we address a broader question of the role of site conditions in modifying the weather forcing upon tree growth. This should be helpful in recognizing limits for generalizations concerning the effects of future climate change on boreal forests.
Study Area
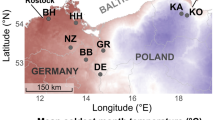
The study area is located in the boreal zone of Western Quebec (Figure 1). In this region, the Precambrian Shield is covered by thick deposits of the Clay Belt, that is, varved clays left by proglacial Lake Barlow-Ojibway. Topography of the area is generally flat, characterized by low elevation hills and a mean altitude between 250 and 400 m above sea level. About half of the area is overlaid by organic deposits of 60 cm thick, residing on clay deposits. The most common soil types on well-drained clay deposits are luvisols, gleysols, and organic soils (Soil Classification Working Group 1998).

Study area and location of sampling sites. Bioclimatic division of the area is based on the Quebec provincial classification system.
The continental climate of the area is characterized by cold winters and warm summers. A cold arctic air mass dominates the area during the cold period and dry tropical air mass tends to dominate during the summer months. The mean annual temperature is −0.7°C, and mean temperatures in January and July are −20.0 and 16.1°C, respectively. The heat sum (degree days above 5°C) reaches 1170, and growing season frosts are common throughout the study area. Total annual precipitation is 905 mm, 35% received during the growing season (Matagami weather station, 49° 84′ N, 77° 84′ W). Annual snowfall is estimated at around 300 mm (Environment Canada 2006).
The northern part of the region is dominated by black spruce (Picea mariana (Mill.) B.S.P.), often mixed with jack pine (Pinus banksiana Lamb.) and trembling aspen (Populus tremuloides Michx.) (Rowe 1972; Gauthier and others 2000). The southern part is dominated by balsam fir (Abies balsamea (L.) Mill.) with paper birch (Betula papyrifera Marsh.), white spruce (Picea glauca Moench), and black spruce. Ericoid shrubs (Vaccinium spp., Rhododendron groenlandicum, Kalmia angustifolia) are dominants of understory vegetation. The moss component of the studied forests is represented by feather mosses (Pleurozium schreberi, Ptilium crista-castrensis, and Hylocomium splendens) and several species of Sphagnum mosses.
The current fire cycle (time required to burn an area equal to the study area) is estimated to be about 400 years, which is significantly longer than those in previous centuries (around 100 years before 1850). The most recent period of increased fire activity was in the beginning of the century (1910–1920) (Bergeron and others 2004b) and intensive forest exploitation started in the region in the 1970s (Bergeron and others 2001). An important factor in spruce population dynamics over most of its distribution range, the spruce budworm (Chorisoneura fumiferana Clem.), has not been of importance for spruce population in the studied region, according to both historical observations (Gray 2008) and dendrochronological reconstructions (Simard and others, unpublished manuscript). The growth pattern of trees in this region may, therefore, provide the opportunity to study the effects of SOL accumulation on tree growth, which may have been little affected by one of the main confounding factors in this part of the North American boreal forest.
Methods
We designed our analyses to test the effects of SOL on climate–growth relationships during the periods dominated by non-anomalous weather conditions and during years with identified regional growth anomalies (pointer years, see below). For the first step of the analyses, we used correlation and response function analyses to identify monthly weather and season-wide variables that were significantly related to spruce growth as depicted by site-level chronologies. Patterns of such relationships were then related to a gradient of SOL. For the second step of the analyses, we studied single-tree chronologies to identify pointer years across all 53 sites included in this study (n trees = 359). The relationship between SOL and growth dynamics during and immediately after such years was analyzed on the subset of sites with developed site chronologies (n sites = 16) using linear and non-linear regression analyses.
Site Selection, Field Sampling, and Climate Data
Two previous studies provided the data for the current analysis (Figure 1). Hofgaard and others (1999) focused primarily on the dendroclimatic response of black spruce on xeric sites with low amounts of SOL, whereas Simard and others (2007) evaluated the impact of increasing organic layer on spruce growth, providing data from sites with SOL typically thicker than 20 cm (Table 1). Stands selected in both studies were naturally established following fire, showed no sign of human influence, and were dominated by black spruce. At each site, 5–29 trees were sampled by taking either two cores at breast height (Hofgaard and others 1999) or a cross-section at the ground level (Simard and others 2007). We considered a potential bias associated with different height of sampling to be minimal and mostly associated with higher ring-width variability in samples measured at ground level. Multiple radii measured in each sample should have minimized a potential variation in ring-width variability due to different sampling heights. Samples from both studies were processed similarly for subsequent analyses. SOL was measured as the distance between the mineral soil and the forest floor surface, and a minimum of 15 such measurements were used to calculate average SOL depth per site.
Monthly climate data (1901–2005) for each site was generated in the ANUSPLIN software package supplied with monthly climate station data for precipitation and minimum and maximum temperature data from the National Climatic Data Center (NCDC) in the U.S. and the Meteorological Service of Canada (MSC) (McKenney and others 2006). The following variables were used: average monthly temperature, sum of monthly precipitation, and growing degree-days (GDD).
Development of Chronologies
Polished and crossdated samples were measured (two radii on cross-sections) using a Velmex micrometer (precision of 0.01 mm) under 40× magnification (Velmex Incorporated, Bloomfield, New York, USA). Occurrence of frost rings was used to aid crossdating of samples prior to their measurement. The dating and homogeneity of tree-ring series were validated with the program COFECHA (Holmes 1999). Two radii measured for each tree were averaged prior to analyses. Crossdated series were detrended in the program ARSTAN (Cook 1985; Cook and Krusic 2005) using a smoothing spline that preserved 50% of the variance at a wavelength of 128 years. Index values for chronologies were computed by division of the original chronology values by the values of the modeled spline. To remove natural persistence (temporal autocorrelation) in the tree-ring growth patterns, each tree-ring curve was modeled as an autoregressive process with the order selected by the first-minimum Akaike Information Criterion (AIC) (Akaike 1974).
The resulting 359 single-tree chronologies from 16 sites were then processed in two different ways. First, for analyses of climate–growth relationships at the stand scale (see “Climate–Growth Relationship, Correlation, and Response Function Analyses” section), we produced 16 residual site chronologies by feeding ARSTAN with all single-tree chronologies from each site. For each site included in the analyses, the number of trees contributing to site chronology varied from 9 to 31 (n trees = 297, Table 2). Second, for pointer year analyses (see “Identification of Pointer Years and Statistical Tests” section), we used all available single-tree chronologies (n trees = 359).
Identification of Pointer Years and Statistical Tests
At first, we used all available single-tree chronologies to identify major positive and negative growth anomalies (pointer years) in black spruce. A pointer year is generally defined as a year with particularly narrow rings in the majority of trees sampled within a site or area (Schweingruber 1996). In this study, a pointer year was defined as a year when ring width was below 5% or above 95% of the ring-width distribution of a single tree. The selection of pointer years was done using the XTRLST routine of the Dendrochronological Program Library (Holmes 1999). Strength (that is, intensity) of a pointer year was assessed by dividing the number of trees expressing the pointer year by the number of trees in the dataset (sample depth) for that year. Pointer years were identified for the period 1901–2003, which corresponds to the instrumental weather data record available for the region. Only pointers, which were present in at least 10% of the trees, were included in the analyses.
Each of the pointer years identified was analyzed for the presence of climatic anomalies in monthly average temperature, monthly total precipitation, and number of GDD, that is, the cumulative number of days during the growing season with a daily temperature average exceeding 5°C. To identify climatic anomalies, 5th and 95th percentiles were calculated for each variable by using normalized monthly data from historical monthly climate models (McKenney and others 2006). As 36 monthly variables in total were analyzed for the presence of climatic anomalies, there was a high probability of finding an association between climatic anomalies and pointer years by chance alone. In fact, given the thresholds used (<5% and >95%), a time span of 100 years, and assuming independence between occurrence of pointer years and climatic anomalies, there was only a 47.8% probability that no anomalies would be associated with four pointer years, even for a single weather variable. To circumvent this problem and test the non-random nature of temporal association between a climatic anomaly and a pointer year, we used superposed epoch analysis (SEA, Haurwitz and Brier 1981). SEA is a statistical method used to solve signal-to-noise problems, which is based on sorting of data into classes dependent on an “event-date” (in our case—a year with climatic anomaly) and on comparing the means of those classes against all possible means. We checked for significant or close-to-significant associations between growth and other years with the same type of weather anomaly. Confidence intervals were generated by resampling the original distribution of growth indices 1,000 times. SEAs were performed in the program EVENT of the Dendrochronological Program Library (Holmes 1999).
Climate–Growth Relationship, Correlation, and Response Function Analyses
Residual chronologies resulting from autoregressive modeling of the 16 site chronologies were used to evaluate climate–growth relationships through correlation and response function analyses (Cook and Kairiukstis 1990). Response function coefficients, in contrast to the common correlation coefficients, are produced using principal component analysis (Fritts and Guiot 1989; Fritts 2001), which transforms original climate data into a set of uncorrelated (orthogonal) components and, therefore, removes intercorrelation present in the climatic data. The coefficients were tested for their statistical significance using the bootstrap method (Efron and Tibshirani 1994). This included random re-sampling of the original dataset with replacement, generating a statistical distribution of a response coefficient, and calculating its 2.5 and 97.5% distribution limits. The values obtained from the original dataset were then compared with the bootstrap-generated distribution limits to assess the significance of the coefficient’s departure from 0 at the 0.05 probability level. We related yearly ring widths to monthly climate variables spanning the period between the month of July of the previous year to the month of August of the current year. In the studied part of black spruce distribution range, the growth typically ceases before the end of the summer. Further, preliminary analyses with varying periods showed that increasing the number of previous season months would have little effect on the outcome of the analyses. Therefore, we attempted to limit the number of months analyzed to ensure statistical stability of the final results. The analyses were performed in the program DendroClim (Biondi and Waikul 2004), using mean monthly temperature and the total sum of monthly precipitation modeled for each site location by historical climate models (McKenney and others 2006). All tests used a significance threshold of 5%.
In addition to the monthly climate variables, we calculated the Seljaninov hydrothermal coefficient (SHC, (Seljaninov 1966), an aridity index that incorporates the influence of both temperature and precipitation variables on growth:
where ΣT and ΣP are the sum of temperature (°C) and precipitation (mm), respectively, for a given time period. We selected the Seljaninov index as a measure of aridity based on its simplicity and clearly defined temporal domain. The index was calculated for the period June–August of the current year growing season.
Finally, we also computed a monthly version of the Drought Code (MDC, Girardin and Wotton 2009). The Drought Code, a component of the Canadian Forest Fire Weather Index, was originally developed to capture moisture content of deep layers of the forest floor (Turner 1972). The numerical value of MDC corresponds to a water holding capacity of 100 mm. For calculations, we utilized the program SimMDC (Girardin and Wotton 2009) and the monthly maximum temperature and total precipitation data from a global 0.5° resolution grid (Mitchell and Jones 2005). MDC values for two grid points nearest to the study area (48.25° N 79.25° W and 48.75° N 78.75° W) were averaged. As monthly MDC values tend to be autocorrelated in time, we extracted the two principal components from six monthly values of MDC (May through October of the current year) to derive independent modes of variation in the MDC values. We used a covariance matrix to center the data around the same mean but to avoid scaling by the standard deviation. To keep axes orthogonal, no axes rotation was performed. We selected only the PC with the respective eigenvalue above 1. We deliberately did not analyze another widely used measure of drought conditions—the Palmer Drought Severity Index (PDSI). By doing so, we wanted to avoid known problems of using PDSI in situations where snow accumulation is a significant contributor to the soil water regime (Akinremi and others 1996). Specifically, calculation of PDSI does not involve the effect of snow accumulation during winter. Further, evapotranspiration taken into account for calculation of PDSI is based exclusively on available soil moisture and no effect of tree-mediated water transport from the soil is considered.
Effect of SOL Accumulation
We assessed the effect of SOL depth on climate–growth relationships (1) using all available years and (2) using only pointer years, which represent periods of regional growth anomalies. In the first case, we simply fit linear or non-linear regressions between SOL thickness and Spearman correlation coefficients reflecting the association between spruce growth and a subset of monthly weather variables used in the correlation and response function analyses (see “Climate–Growth Relationship, Correlation, and Response Function Analyses” section). We used a subset of the monthly temperature averages and total sums of precipitation, which significantly related to black spruce growth in the response function analyses (see the previous sub-section).
In the second case (pointer years), we analyzed two variables as a function of SOL thickness: (a) pointer year strength, expressed as the proportion of trees expressing a particular pointer year in a site, and (b) pointer year effect on radial growth, expressed as the ratio between cumulative increments three years prior and three years following a pointer year. In the second case (b), preliminary analyses with varying timeframes (1–5 years) produced similar results and are not presented here. Linear and non-linear regression models were fitted between SOL and the two above-mentioned variables, using the Gauss–Newton algorithm for nonlinear estimation (Björck 1996). Gauss–Newton is a common algorithm to solve nonlinear least-squares problems. In our case, we expected a nonlinear relationship between the variables, and assumed independence of error from the dependent variable, that is, additive error component.
We did not analyze the water table (WT) dynamics because preliminary analyses with the same set of dependent variables showed that WT depth is an inferior predictive variable as compared to SOL. Previous studies have shown that there is considerable variation in the depth of the WT in spruce stands, both over the course of the year and among years (Simard and others 2007). Another way to describe soil water regime would be to locate the border separating generally non-oxidized and oxidized sediments in the soil profile. Such estimation would provide a more biologically meaningful measure of the WT height (Simard and others 2007; Fenton and others 2006). However, previous studies have shown that oxygen zone position is strongly correlated with thickness of the forest floor (Fenton and others 2006), which supported our choice of SOL as the only variable in the analysis, representing soil water conditions.
Results
The sampled sites showed a wide variation in SOL, ranging from forests with virtually no accumulated organic layer to forests with more than 0.5 m of SOL (Table 1). The 16 site chronologies showed moderate to high levels of growth similarity among trees within a site, with signal-to-noise ratios ranging from 5.4 to 24.0 (Table 2). This pattern was confirmed by the relatively high values (31–61%) of variance explained by the first eigenvector obtained during decomposition of total variance in growth within the site. Variance in the first eigenvector represents the main pattern of growth variability among trees, with high loadings on this eigenvector being indicative of the presence of such a dominant pattern. Alternative representation of this phenomenon is signal strength (SS) statistics (Wigley and others 1984). Only in four cases out of 16, a site chronology did not reach a SS value of 0.90 and in all cases SS values reached 0.80 for the period analyzed.
Correlation and Response Function Analyses
Correlation and response function analyses revealed two periods when black spruce growth was related to the monthly temperature data (Figure 2). The strongest association was a positive effect on growth of February–April temperatures of the current growing season. Growth was also negatively correlated with temperature in August of the previous year. Although we observed five significant correlation coefficients between growth and summer temperatures, only one response function coefficient was significant over that period. As for precipitation, tree growth was negatively correlated to current year January precipitation and was generally positively related to summer precipitation.

Number of significant correlation and response function coefficients between black spruce growth and monthly weather variables for the period ranging from previous year August to current year August in all sites combined (n sites = 16 sites). Turn of the year is indicated by dashed line. Significance of departures from zero at 0.05 level was assessed by generating 1,000 bootstrapped resamples to obtain empirical distribution of respective coefficients.
Pointer Year Analysis
Main pointer years included narrow rings (<5th percentile) in 1956, 1974, 1989, and 1994 (negative growth anomalies), and wide rings (>95th percentile) in 1908, 1924, and 2001 (positive anomalies, Table 3). The earliest negative pointer year (1956) was associated with an unusually cold spring coupled with high late-summer temperatures during the previous year. An exceptionally short growing season, warm August during the previous year, and dry March of the current year were associated with the pointer year 1974. The temperature in both April and May was low and close to the thresholds (−3.0°C, 15th percentile and 4.5°C, 7th percentile, respectively). Abundant winter precipitation and low summer precipitation were observed in 1989. Unusually cold conditions in the winter and previous fall were associated with the most recent negative pointer year (1994). As for the positive pointers (wide rings), no climate anomalies were recorded in the growing season of the two earliest years (1908 and 1924). However, the temperature for the month of August of the year previous to the 1924 growing season was low and close to the 5% threshold (12.7°C, 6th percentile). The latest positive pointer year (2001) was characterized by a generally warm and long growing season.
To assess the statistical significance of the temporal association between weather and growth extremes, we used SEA (Figure 3), assuming that if the association between an extreme weather event and a spruce pointer year was non-random, all growing seasons showing such a weather anomaly should result in consistently narrow (for negative pointer years) or wide (for positive pointer years) rings over the whole period jointly covered by weather and tree-ring data. Out of the 14 unique pairs of climatic anomalies and negative pointer years in spruce (Table 3), only four types of events tended to be linked with negative growth anomalies during the second half of twentieth century. Tree growth was significantly (P < 0.05) lower during years with short growing seasons and low March temperatures (Figure 4A, B). In the case of July precipitation, the ring-width departure value for the focal year as well as for the following year was significant at the 0.1 level (Figure 4C). Finally, unusually higher temperatures during the previous year’s August tended to be associated with negative growth anomalies, the result being significant at the 0.1 level (Figure 4D).

Relationship between occurrence of weather anomalies and tree-ring increment in black spruce—results of superposed epoch analysis for the period 1901–2003. Monthly weather variables were selected from a list of anomalies associated with spruce negative pointer years (Table 3). Only results significant at the 10% level are shown. Bootstrap-derived 95% confidence limits are indicated by dashed lines. Open circles represent growth index average values for the years with climate anomalies (focal years), and filled circles represent averages for the years with one to five years offset from the focal year. In the analyses of number of GDD, March temperature, and July precipitation, the spruce chronology was tested against the list of negative weather anomalies. In the analyses of previous year August temperature, the test was done against a list of positive weather anomalies. List of years with weather anomalies is given for each graph.

Relationship between depth of soil organic layer (SOL) and the strength of growth–climate correlations for 16 sites and six weather variables. Statistical details of the analyses are given in the Table 4. SHC and MDC stand for Seljaninov hydrothermal coefficient and monthly Drought Code, respectively. Lower SHC values are indicative of higher drought conditions.
Effect of SOL Accumulation
Based on the results from the correlation and response function analyses (Figure 2), we selected three monthly variables (January precipitation, average February–March temperature, and average temperature of the previous year’s August) and two drought indices (summer months SHC and MDC) to evaluate if tree growth response to monthly weather varied along a gradient of SOL depth. In the case of MDC, monthly values were first subjected to a principal component analysis to extract two main axes of monthly MDC variability. Cumulatively, the two first principal components accounted for 89.5% of the variability in MDC values (55.9% and 23.6% for the first and the second components, respectively). The first principal component was associated with MDC during mid- and late-summer (loadings for the months of June through October were 0.85, 0.92, 0.87, 0.71 respectively) and the second principal component was associated with early-season MDC (loadings for the months of May and June were 0.79 and 0.65, respectively).
For January precipitation (Figure 4A, Table 4) and summer SHC (Figure 4D, Table 4), an increase in SOL was associated with a transition from a negative to a positive relationship between growth and these weather variables. SOL depth did not affect the relationship between tree growth and previous year’s August temperature (Figure 4B, Table 4), nor between the growth and February–March temperature (Figure 4C, Table 4). Increase in SOL depth was also associated with consistent changes in the MDC–growth relationship (Figure 4E, F, Table 4). We observed a significant trend toward a decrease in the negative association between the main mode of MDC annual variation (June–October period) and growth. A similar pattern was revealed with respect to the MDC variability in spring. In both cases, the inflexion in the regression curves was associated with a threshold of approximately 20–30 cm of SOL.
Expression of the negative pointer years 1956, 1974, 1989 and 1994, represented as the proportion of trees recording that pointer year at each site, increased with increasing depth of SOL (Figure 5, Table 4). The effect of SOL explained the highest amount of variation in the proportion of pointer years during 1974 (41.0%), which was the strongest pointer year in the area (Figure 5B). No relationship was found between the ratio of pre- and post-pointer 3-year cumulative growth increments during any of the pointer years studied.

Relationship between thickness of soil organic layer and expression of four negative pointer years (see Table 3) in 16 site-level chronologies. Expression of a pointer year was partitioned as (1) a proportion of trees recording the pointer year in question at each site (black circles, left OY axis, solid line as fitting curve) or as (2) a ratio between the 3-year cumulative growth increments before and after the pointer year (white circles, right OY axis, dashed line as fitting curve). Significant and close to significant regressions were obtained in the case (1) only. Statistical details of the analyses are given in the Table 4.
Discussion
In this study, we evaluated the possibility of a variation in growth response to annual weather along a gradient in SOL depth in the black spruce forests of the Quebec Clay Belt. Although the impact of site conditions on growth of boreal trees has been thoroughly studied (Schweingruber 1996), to the best of our knowledge, no study has verified these relationships across a gradient of SOL using a large dendrochronological dataset. A novelty of this study is also in the joint analysis of non-anomalous growth through correlation and response function analyses, and of single negative growth anomalies (pointer years). Below, we first discuss the general patterns of spruce growth reaction to annual weather and the temporal pattern of pointer years, which is then followed by the analysis of SOL–growth interactions for non-anomalous weather conditions, and, finally, for years with major growth anomalies.
Growth Response of Black Spruce to Annual Weather
Temperature regime in late winter–early spring appears to be the dominant factor controlling diameter increment of black spruce in the studied region (Figure 2). Particularly, we found a positive association between the increment and the temperatures in February–April, which was consistently replicated across the sites. Influence of late winter–early spring temperatures, also reported in other studies ((Huang and others 2009), is likely related to the dynamics of soil temperature and, particularly, the maximum depth of the zone with temperatures below 0°C. High late-winter temperatures may cause faster snowmelt and faster thawing of soil at the start of the growing season. A positive effect of April temperatures may be also associated with faster snow melting. A longer growing season may promote faster growth and thus greater rates of biomass accumulation in black spruce forests. This pattern may occur more frequently in the future because the start of the growing season in the temperate zone of the Northern Hemisphere has been recently shown to occur earlier in the year (Linderholm 2006; Thompson and Clark 2008).
Surprisingly, positive correlation between growth and summer temperatures, believed to be the main factor driving tree growth dynamics in the boreal zone (Briffa and others 2002), was not observed in our dataset. Out of five significant correlation coefficients between annual growth increment and summer month temperatures, only one was positive, whereas the other four were negative. Similarly to our study, the relationship between black spruce growth and temperature did not reveal a consistent pattern in previous studies where both positive (Dang and Lieffers 1989a; Larsen and MacDonald 1995) and negative (Brooks and others 1998) correlations were reported. We propose that such differences in responses may be related to study site conditions and particularly, to the amount of SOL (see “Effect of SOL on Weather–Growth Relationships—Non-Anomalous Weather” sub-section).
Overall, our results indicate that diameter growth in black spruce is not limited by the temperature during the growing season, but responds primarily to its length, as controlled by the start of physiological activity in spring. Generally, the length of the growing season is considered as a critical variable affecting the response of boreal trees to future climatic changes (Linderholm 2006; Girardin and others 2008). A longer growing season in the boreal zone is commonly associated with increased annual biomass accumulation, but also with higher risks of damage to photosynthetic and conducting tissues during early season cold spells (Hanninen 2006; Gu and others 2008). The almost complete absence of significant relationships between growth and monthly temperatures in the previous fall (September–November) and first two winter months suggests that spruce growth is rather insensitive to the temperature regime of the preceding dormant season.
Well-replicated negative association between previous year August temperatures and current year growth indicates that high evapotranspiration demand of spruce at the end of the previous growing season may interact with translocation of bioassimilates (Sakai and Larcher 1987) and decrease the efficiency of pre-winter hardening, for example, by lowering sugar concentrations in the main stem and the foliage (Strimbeck and others 2008). A similar negative impact of high evapotranspiration demand of previous year growing season was noted in the studies of black spruce in Alberta peatlands (Dang and Lieffers 1989b), white spruce growth in Manitoba (Chhin and others 2004), subalpine fir in the Pacific Northwest (Peterson and others 2002), Douglas-fir in the northwestern United States (Littell and others 2008), trembling aspen in northeastern British Columbia (Leonelli and others 2008), and in the pedunculate oak in Southern Sweden (Drobyshev and others 2008). Previous year conditions are believed to be closely connected to the development of earlywood part of the current year ring primarily due to reliance of cell division and expansion on previous year reserves (Vaganov and Shashkin 2000; Lebourgeois and others 2004).
The effect of precipitation on spruce growth is less pronounced than that of temperature, as suggested by the low number of significant correlation and response function coefficients (Figure 2). Precipitation in January (in the form of snow) was negatively correlated with tree growth in the correlation analysis, but this effect was not statistically significant in the response function analysis. As response function analysis accounts for the intercorrelation among independent variables, this pattern may indicate that the effect of snow accumulation on the growing season onset is effectively overridden by the dynamics of soil temperature regime and possibly, the speed of snow melting, both of which are controlled by late-winter and spring temperatures.
Late-spring and summer precipitation variables had a limited impact on spruce growth (Figure 2). Negative correlations between growth and May precipitation may indicate situations where the soil, still sufficiently filled with snowmelt water, is waterlogged by excessive rains. Later in the season, higher evapotranspiration demand of spruce forest and reduction in soil water reserves during summer months change the sign of this relationship. The observed effect reflects generally high transpiration rates in spruce trees and difficulty in getting access to the water below superficially located root systems (Desplanque and others 1998). Our results, therefore, indicate that even in the area with sufficient amounts of precipitation throughout the year, there are sites probably experiencing moderate summer droughts and positively responding to growing season precipitation. Our conclusions are supported by the results of the CO2 measurement studies, which identified the variation in the onset of photosynthesis in spring as the main factor, and drought-related reduction in physiological activity as one of secondary factors responsible for annual changes in CO2 flux of a mature black spruce forest (McMillan and others 2008).
Pointer Years
Negative growth anomalies of black spruce occurred in 1956, 1974, 1989, and 1994 across the region but were likely caused by different weather conditions, as shown by SEA analyses (Table 3). In three cases out of four results of SEA analyses pointed to a likely factor responsible for the growth anomaly. Despite a number of weather extremes associated with that growing season, none of the main growth anomalies were likely related to a similar set of weather conditions over the course of the twentieth century. We link the 1956 negative growth anomaly to a short growing season (148 GDD, within the lower 9% of respective distribution). This interpretation is supported by the occurrence of growth anomalies during other years with very short growing seasons (Figure 3A) and by the high frequency in 1956 of light rings (M. Simard, unpublished data), which are caused by shortened growing seasons in subarctic Quebec (Yamaguchi and others 1993). An exceptionally low March average temperature could be another factor behind this growth anomaly: the negative anomalies of this variable were consistently related to low spruce tree-width increments (Figure 3B).
Year 1974 was the most prominent pointer year in the region (present in 39% of all analyzed tree chronologies) and could have been caused by a short growing season or a high temperature in the previous year August (Figure 3A, D). The length of the growing season is important for structural ring development in East Siberian coniferous trees (Kirdyanov and others 2003) and controls balsam fir ring growth in northeastern Quebec (Deslauriers and Morin 2005).
Two weather anomalies (high precipitation in January and low precipitation in July) could have caused the 1989 pointer year (Table 1, Figure 2), but SEA suggests that the deficit in summer precipitation of the current year may be the likely factor responsible for the growth anomaly (Figure 3C). This result, together with the moderate effect of monthly precipitation averages for the summer months, indicated nonlinearity in the growth response to water availability.
The 1994 growth anomaly was more difficult to interpret. We speculate that a cold period in January and a limited amount of snow precipitation may have resulted in frost damage to tree root systems. Generally, interpretation of temporal association of spruce negative pointer years was in line with patterns of weather–growth relationships revealed by correlation and response function analyses. This implies that the growth pattern during non-extreme weather conditions and during the periods of weather anomalies is controlled by a similar set of weather variables. These are temperature regimes (1) at the end of the previous growing season and (2) in late winter–early spring of the current growing season, and, to a lesser extent, (3) water deficit in the middle of the summer period. Such similarity in weather controls of tree growth under extreme and non-extreme weather settings, which may facilitate modeling work, is not necessarily present in the pattern of tree growth response to climate. For example, a dendrochronological study of pedunculate oak growth in the Southern Sweden has demonstrated that growth under non-extreme weather conditions was mainly controlled by summer precipitation but that most of the negative pointer years were associated with temperature anomalies (Drobyshev and others 2008).
Two years with positive growth anomalies (1908 and 1924) were not associated with any weather anomalies in the instrumental weather data. This indicated that current weather conditions in the region are generally favorable for black spruce growth. Alternatively, inferior quality of weather data during the early 1900s, primarily due to the limited number of weather stations in the region (McKenney and others 2006) may not have captured more local weather variations.
Effect of SOL on Weather–Growth Relationships—Non-Anomalous Weather
An increase in the amount of SOL affects the growth response of black spruce to summer aridity. Specifically, the negative association between water deficit and tree growth in stands with shallow organic layers changes to a positive one in stands with thicker SOL (>30 cm; Figure 4D). This effect indicates that trees growing in sites with thick SOL were less sensitive to moderate summer droughts but instead may have benefited from higher air temperatures and/or more aerobic conditions in the soil. Similarly, MDC was positively associated with tree growth at the beginning of the growing season in sites with thick SOL (Figure 4F) and was negatively associated with tree growth in the middle and at the end of the growing season in sites with shallower SOL (Figure 4E).
Although additional data are needed to quantify this interaction more precisely, the inflexions in the fitted nonlinear regression curves suggest a threshold value of SOL at around 20–30 cm. This level of SOL thickness corresponds to a significant threshold in the accumulation of SOL at which point the WT moves from the mineral soil to the organic layer, which occurs about 400 years after a high-severity fire (Simard and others 2007). With the current fire cycle, more stands are expected to reach that threshold in SOL thickness, and therefore, to show a positive growth response to summer droughts. However, the rate of SOL removal is also controlled by fire severity, which introduces additional uncertainty in understanding future trends in SOL accumulation (Conard and others 2002; Lecomte and others 2006). The current state of knowledge about future fire activity in the boreal zone makes it difficult to precisely project future trends in SOL accumulation (Bergeron and others 2001; Bergeron and others 2004b). However, the effect of SOL on weather–growth relationships during non-anomalous weather was only moderate and may be difficult to detect, as indicated by the low absolute values of correlation coefficients.
Effect of SOL on Weather–Growth Relationships—Anomalous Weather
The proportion of trees expressing a particular pointer year significantly increased with SOL thickness during three pointer years (Figure 5). The effect was the most pronounced in 1974 and 1989, two major pointer years. In 1974, when an extremely short growing season was the likely cause of the growth anomaly, thicker SOL may have delayed warming of the soil (Simard and others 2007) and onset of root activity at the beginning of the growing season. In 1989, mid-summer water deficit may have been stronger on sites with high SOL, probably due to the fact that low-density organic layers tend to be dryer than underlying mineral soil under conditions of severe water deficit, and due to wetting resistance of peat-rich soils inhibiting water uptake by roots (Schwarzel and others 2002). The opposite conclusion reached for anomalous and non-anomalous weather conditions implies a high degree of nonlinearity in the pattern of climate–growth interactions along the gradient in SOL thickness. Particularly, whereas thick SOL may successfully buffer against low-water availability under non-extreme levels of water deficit, a prolonged mid-season drought may make them even dryer than underlying layers of mineral soil.
Even if the frequency of negative growth anomalies increased on sites with higher amounts of SOL, the long-term effect of such growth reductions appeared limited. SOL was never significantly related to the ratio of 3-year cumulative diameter increments prior to and immediately following the pointer year. Annual growth anomalies in spruce are, therefore, not an indication of growth suppression at longer temporal scales (≤3 years). We suggest that this may be due to the plasticity of black spruce and its tolerance of a wide range of environmental conditions, which even during years with anomalous weather conditions does not reach the levels associated with structural damage to tree tissues.
Possible Effects of Future Climatic Change on Black Spruce Ecosystem Structure and Function
The primary drivers of SOL accumulation are precipitation, which favors SOL accumulation across the landscape, and fire, which removes locally some of the accumulated SOL. Regional climate models for Quebec’s boreal region predict significantly higher summer temperatures but only a slight increase in growing season precipitation (van Bellen and others, unpublished), with changes in seasonal average precipitation staying within 10% of present-day values (Plummer and others 2006). Coarser-scale models predict both decreases and increases in summer precipitation for the eastern Canadian boreal forest (Meehl and others 2007).
In this study, we have shown that accumulation of SOL affected black spruce growth response to climatic variation. Thicker SOL translates into decreased growth sensitivity to average summer aridity, a pattern thought as generally positive for spruce growth. Trees growing in sites with thick SOL were also more affected by extreme climatic events such as short growing seasons, cold springs, and drought. Such negative effects of SOL may be increasingly more important in the future, given the higher frequency of climate anomalies. Direct effects of climate on spruce growth (climate → growth) can, therefore, be strongly modified by the indirect effects of climate through SOL accumulation (climate → SOL → growth), and through interactions with the fire regime (climate → fire regime → SOL → growth). Such a complex structure of interactions suggests strong but complex relationships between SOL effects on growth and ecosystem-level dynamics of black spruce forests.
Finally, continuing SOL accumulation is also expected to have a strong effect on nutrient cycling (Sirén 1955; Brais and others 2000) and on forest productivity. In the Clay Belt, SOL accumulation has been shown to account for 50–80% of the decrease in tree productivity (Simard and others 2007), the amplitude of the effect being dependent on site history and topographic variables. In turn, the role of the observed climate–growth relationships to the dynamics of carbon accumulation in black spruce forests appears limited. This is due to the relatively minor proportion of carbon stored in the canopy strata as compared to carbon in the SOL and in the soil (Lecomte and others 2006) and general decrease in the biomass production with increase in SOL (Simard and others 2007). We speculate that increased SOL could mitigate the negative effect of moderate summer drought on growth, thus maintaining carbon sinks through tree biomass accumulation during such periods. However, an increased SOL can also mean a higher probability of strong growth decline in trees, thus contributing to higher inter-annual variability in the carbon storing capacity of the tree strata.
Conclusion
In this study, we have shown that accumulation of SOL affected the black spruce growth response to climatic variations. Trees growing in sites with thick SOL were less affected by moderate summer drought but were also more affected by extreme climatic events such as short growing seasons, cold springs, and severe drought. The direct effects of climate on spruce growth (climate → growth) can, therefore, be strongly modified by the indirect effects of climate through SOL accumulation (climate → SOL → growth), and through interactions with the fire regime (climate → fire regime → SOL → growth). Such a complex structure of interactions suggests strong but complex relationships between SOL effects on growth and ecosystem-level dynamics of black spruce forests.
References
Akaike H. 1974. A new look at the statistical model identification. IEEE Trans Autom Control 16:716–23.
Akinremi OO, Mcginn SM, Barr AG. 1996. Evaluation of the Palmer Drought index on the Canadian prairies. J Clim 9:897–905.
Allison SD, Treseder KK. 2008. Warming and drying suppress microbial activity and carbon cycling in boreal forest soils. Glob Change Biol 14:2898–909.
Amiro BD, Stocks BJ, Alexander ME, Flannigan MD, Wotton BM. 2001. Fire, climate change, carbon and fuel management in the Canadian boreal forest. Int J Wildland Fire 10:405–13.
Apps MJ, Kurz WA, Luxmoore RJ, Nilsson LO, Sedjo RA, Schmidt R, Simpson LG, Vinson TS. Boreal forests and tundra. 1993. International workshop on terrestrial biospheric carbon fluxes: quantification of sinks and sources of CO2. Bad Harzburg (FRG), 1–5 March 1993.
Archambault S, Bergeron Y. 1992. An 802-year tree-ring chronology from the Quebec boreal forest. Can J For Res 22:674–82.
Bergeron Y, Gauthier S, Kafka V, Lefort P, Lesieur D. 2001. Natural fire frequency for the eastern Canadian boreal forest: consequences for sustainable forestry. Can J For Res 31:384–91.
Bergeron Y, Flannigan M, Gauthier S, Leduc A, Lefort P. 2004a. Past, current and future fire frequency in the Canadian boreal forest: Implications for sustainable forest management. Ambio 33:356–60.
Bergeron Y, Gauthier S, Flannigan M, Kafka V. 2004b. Fire regimes at the transition between mixedwood and coniferous boreal forest in Northwestern Quebec. Ecology 85:1916–32.
Biondi F, Waikul K. 2004. DENDROCLIM2002: a C++ program for statistical calibration of climate signals in tree-ring chronologies. Comput Geosci 30:303–11.
Björck A. 1996. Numerical methods for least squares problems. Philadelphia: SIAM.
Bonn S. 2000. Competition dynamics in mixed beech-oak stands and its modifications expected due to climate changes. Allg For Jagdztg 171:81–8.
Brais S, David P, Ouimet R. 2000. Impacts of wild fire severity and salvage harvesting on the nutrient balance of jack pine and black spruce boreal stands. Forest Ecol Manag 137:231–43.
Briffa KR, Osborn TJ, Schweingruber FH, Jones PD, Shiyatov SG, Vaganov EA. 2002. Tree-ring width and density data around the Northern Hemisphere: part 1, local and regional climate signals. Holocene 12:737–57.
Brooks JR, Flanagan LB, Ehleringer JR. 1998. Responses of boreal conifers to climate fluctuations: indications from tree-ring widths and carbon isotope analyses. Can J For Res 28:524–33.
Chhin S, Wang GG, Tardif J. 2004. Dendroclimatic analysis of white spruce at its southern limit of distribution in the Spruce Woods Provincial Park, Manitoba, Canada. Tree-Ring Res 60:31–43.
Conard SG, Sukhinin AI, Stocks BJ, Cahoon DR, Davidenko EP, Ivanova GA. 2002. Determining effects of area burned and fire severity on carbon cycling and emissions in Siberia. Clim Change 55:197–211.
Cook ER. 1985. A time-series analysis approach to tree ring standardization. PhD dissertation edition, The University of Arizona.
Cook ER, Kairiukstis LA. 1990. Methods of Dendrochronology: applications in the environmental sciences. Dordrecht: Kluwer.
Cook ER, Krusic PJ. 2005. Program ARSTAN. A tree-ring standardization program based on detrending and autoregressive time series modeling, with interactive graphics. Palisades (NY): Tree-Ring Laboratory, Lamont Doherty Earth Observatory of Columbia University.
Crawford R, Jeffree C, Rees W. 2003. Paludification and forest retreat in northern oceanic environments. Ann Bot (Lond) 91:213–26.
Dang QL, Lieffers VJ. 1989a. Assessment of patterns of response of tree-ring growth of black spruce following peatland drainage. Can J For Res 19:924–9.
Dang QL, Lieffers VJ. 1989b. Climate and annual ring growth of black spruce in some Alberta peatlands. Can J Bot 67:1885–9.
Deslauriers A, Morin H. 2005. Intra-annual tracheid production in balsam fir stems and the effect of meteorological variables. Trees-Struct Funct 19:402–8.
Desplanque C, Rolland C, Michalet R. 1998. Comparative dendroecology of the silver fir (Abies alba) and the Norway spruce (Picea abies) in an Alpine valley of France. Can J For Res 28:737–48.
Dixon RK, Brown S, Houghton RA, Solomon AM, Trexler MC, Wisniewski J. 1994. Carbon pools and flux of global forest ecosystems. Science 263:185–90.
Drobyshev I, Niklasson M, Linderson H, Sonesson K. 2008. Influence of annual weather on growth of pedunculate oak in southern Sweden. Ann For Sci 65: doi:10.1051/forest:2008033.
Efron B, Tibshirani RJ. 1994. An introduction to the bootstrap. London: Chapman & Hall.
Environment Canada. 2006. Canadian climate normals 1971–2000. www.climate.weatheroffice.ec.gc.ca.
Fenton NJ, Bergeron Y. 2007. Sphagnum community change after partial harvest in of black spruce boreal forests. For Ecol Manag 242:24–33.
Fenton N, Lecomte N, Legare S, Bergeron Y. 2005. Paludification in black spruce (Picea mariana) forests of eastern Canada: potential factors and management implications. Forest Ecol Manag 213:151–9.
Fenton N, Legare S, Bergeron Y, Pare D. 2006. Soil oxygen within boreal forests across an age gradient. Can J Soil Sci 86:1–9.
Fritts HC. 2001. Tree rings and climate. Caldwell: The Blackburn Press.
Fritts HC, Guiot J. 1989. Methods of calibration, verification, and reconstruction. In: Cook ER, Kairiukstis LA, Eds. Methods of dendrochronology. Application in the environmental sciences. Dordrecht: Kluwer Academic Publishers. p 163–217.
Gauthier S, De Grandpré L, Bergeron Y. 2000. Differences in forest composition in two boreal forest ecoregions of Quebec. J Veg Sci 11:781–90.
Girardin MP, Wotton BM. 2009. Summer moisture and wildfire risks across Canada. J Appl Meteorol Climatol 48:517–33.
Girardin MP, Raulier F, Bernier PY, Tardif JC. 2008. Response of tree growth to a changing climate in boreal central Canada: a comparison of empirical, process-based, and hybrid modelling approaches. Ecol Model 213:209–28.
Gower ST, Vogel J, Stow TK, Norman JM, Steele SJ, Kucharik CJ. 1997. Carbon distribution and above-ground net primary production of upland and lowland boreal forest in Saskatchewan and Manitoba. J Geophys Res 104:29029–41.
Gray DR. 2008. The relationship between climate and outbreak characteristics of the spruce budworm in eastern Canada. Clim Change 87:361–83.
Greene DF, MacDonald SE, Haeussler S, Domenicano S, Noel J, Jayen K, Charron I, Gauthier S, Hunt S, Gielau ET, Bergeron Y, Swift L. 2007. The reduction of organic-layer depth by wildfire in the North American boreal forest and its effect on tree recruitment by seed. Can J For Res 37:1012–23.
Gu L, Hanson PJ, Mac Post W, Kaiser DP, Yang B, Nemani R, Pallardy SG, Meyers T. 2008. The 2007 eastern US spring freezes: Increased cold damage in a warming world? Bioscience 58:253–62.
Hanninen H. 2006. Climate warming and the risk of frost damage to boreal forest trees: identification of critical ecophysiological traits. Tree Physiol 26:889–98.
Haurwitz MW, Brier GW. 1981. A critique of the superposed epoch analysis method: its application to solar-weather relations. Mon Weather Rev 109:2074–9.
Hofgaard A, Tardif J, Bergeron Y. 1999. Dendroclimatic response of Picea mariana and Pinus banksiana along a latitudinal gradient in the eastern Canadian boreal forest. Can J For Res 29:1333–46.
Holmes RL. 1999. Dendrochronological Program Library (DPL). Users manual. http://www.ltrr.arizona.edu/pub/dpl/. Tucson (AZ): Laboratory of Tree-Ring Research, University of Arizona.
Huang J, Tardif JC, Bergeron Y, Denneler B, Berningen F, Girardin M. 2009. Radial growth response of four dominant boreal tree species to climate along a latitudinal gradient in the eastern Canadian boreal forest. Glob Change Biol. doi:10.1111/j.1365-2486.2009.01990.x.
Islam MA, MacDonald SE. 2004. Ecophysiological adaptations of black spruce (Picea mariana) and tamarack (Larix laricina) seedlings to flooding. Trees-Stuct Funct 18:35–42.
Johnstone JF, Chapin FS. 2006. Fire interval effects on successional trajectory in boreal forests of northwest Canada. Ecosystems 9:268–77.
Kimball JS, Thornton PE, White ME, Running SW. 1997. Simulating forest productivity and surface-atmosphere carbon exchange in the BOREAS study region. Tree Physiol 17:589–99.
Kirdyanov A, Hughes M, Vaganov E, Schweingruber F, Silkin P. 2003. The importance of early summer temperature and date of snow melt for tree growth in the Siberian Subarctic. Trees-Stuct Funct 17:61–9.
Komarov AS, Kubasova TS. 2007. Modeling organic matter dynamics in conifer-broadleaf forests in different site types upon fires: a computational experiment. Biol Bull 34:408–16.
Kurbatova J, Li C, Varlagin A, Xiao X, Vygodskaya N. 2008. Modeling carbon dynamics in two adjacent spruce forests with different soil conditions in Russia. Biogeosciences 5:969–80.
Laiho R. 2006. Decomposition in peatlands: reconciling seemingly contrasting results on the impacts of lowered water levels. Soil Biol Biochem 38:2011–24.
Larsen CPS, MacDonald GM. 1995. Relations between tree-ring widths, climate, and annual area burned in the boreal forest of Alberta. Can J For Res 25:1746–55.
LeBarron RK. 1945. Adjustment of black spruce root systems to increasing depth of peat. Ecology 26:309–11.
Lebourgeois F, Cousseau G, Ducos Y. 2004. Climate-tree-growth relationships of Quercus petraea Mill. stand in the Forest of Berce (“Futaie des Clos”, Sarthe, France). Ann For Sci 61:361–72.
Lecomte N, Simard M, Bergeron Y, Larouche A, Asnong H, Richard PJW. 2005. Effects of fire severity and initial tree composition on understorey vegetation dynamics in a boreal landscape inferred from chronosequence and paleoecological data. J Veg Sci 16:665–74.
Lecomte N, Simard M, Fenton N, Bergeron Y. 2006. Fire severity and long-term ecosystem biomass dynamics in coniferous boreal forests of eastern Canada. Ecosystems 9:1215–30.
Leonelli G, Denneler B, Bergeron Y. 2008. Climate sensitivity of trembling aspen radial growth along a productivity gradient in northeastern British Columbia, Canada. Can J For Res 38:1211–22.
Linderholm HW. 2006. Growing season changes in the last century. Agric For Meteorol 137:1–14.
Littell JS, Peterson DL, Tjoelker M. 2008. Douglas-fir growth in mountain ecosystems: water limits tree growth from stand to region. Ecol Monogr 78:349–68.
McKenney DW, Pedlar JH, Papadopol P, Hutchinson MF. 2006. The development of 1901–2000 historical monthly climate models for Canada and the United States. Agric Forest Meteorol 138:69–81.
McMillan AMS, Winston GC, Goulden ML. 2008. Age-dependent response of boreal forest to temperature and rainfall variability. Glob Change Biol 14:1904–16.
Meehl GA, Stocker TF, Collins WD, Friedlingstein P, Gaye AT, Gregory JM, Kitoh A, Knutti R, Murphy JM, Noda A, Raper SCB, Watterson IG, Weaver AJ, Zhao Z-C. 2007. Global climate projections. In: Solomon S, Qin D, Manning M, Chen Z, Marquis M, Averyt KB, Tignor M, Miller HL, Eds. Climate change 2007: the physical science basis. Contribution of Working Group I to the fourth assessment report of the Intergovernmental Panel on climate change. Cambridge, New York (NY): Cambridge University Press.
Mitchell TD, Jones PD. 2005. An improved method of constructing a database of monthly climate observations and associated high-resolution grids. Int J Climatol 25:693–712.
Peltoniemi M, Thurig E, Ogle S, Palosuo T, Schrumpf M, Wutzler T, Butterbach-Bahl K, Chertov O, Komarov A, Mikhailov A, Gardenas A, Perry C, Liski J, Smith P, Makipaa R. 2007. Models in country scale carbon accounting of forest soils. Silva Fenn 41:575–602.
Peterson DW, Peterson DL, Ettl GJ. 2002. Growth responses of subalpine fir to climatic variability in the Pacific Northwest. Can J For Res 32:1503–17.
Plummer DA, Caya D, Frigon A, Cote H, Giguere M, Paquin D, Biner S, Harvey R, De Elia R. 2006. Climate and climate change over North America as simulated by the Canadian RCM. J Clim 19:3112–32.
Prescott CE, Maynard DG, Laiho R. 2000. Humus in northern forests: friend or foe? Forest Ecol Manag 133:23–36.
Ritchie JC. 2004. Post-glacial vegetation of Canada. Cambridge: Cambridge University Press.
Rowe JS. 1972. Forest regions of Canada. Ottawa (ON): Environment Canada.
Sakai A, Larcher W. 1987. Frost survival of plants. Ecological studies 62. Berlin: Springer.
Schwarzel K, Renger M, Sauerbrey R, Wessolek G. 2002. Soil physical characteristics of peat soils. J Plant Nutr Soil Sci—Z Pflanz Bodenkunde 165:479–86.
Schweingruber FH. 1996. Tree rings and environment. Dendroecology. Berne: Paul Haupt Publishers, p 609
Seljaninov GT. 1966. Agroclimatic map of the world. Leningrad: Hydrometeoizdat Publishing House.
Silins U, Rothwell RL. 1999. Spatial patterns of aerobic limit depth and oxygen diffusion rate at two peatlands drained for forestry in Alberta. Can J For Res 29:53–61.
Simard M, Lecomte N, Bergeron Y, Bernier PY, Pare D. 2007. Forest productivity decline caused by successional paludification of boreal soils. Ecol Appl 17:1619–37.
Sirén G. 1955. The development of spruce forest on raw humus sites in northern Finland and its ecology. Acta Forestalia Fennica 62:1–363.
Smith P, Fang CM, Dawson JJC, Moncrieff JB. 2008. Impact of global warming on soil organic carbon. Adv Agron 97:1–43.
Soil Classification Working Group. 1998. The Canadian system of soil classification. 3rd edn. Ottawa (ON): Agriculture and Agri-Food Canada.
Strimbeck GR, Kjellsen TD, Schaberg PG, Murakami PF. 2008. Dynamics of low-temperature acclimation in temperate and boreal conifer foliage in a mild winter climate. Tree Physiol 28:1365–74.
Tarnocai C. 2006. Classification systems: Canadian. In: Lal R, Ed. Encyclopedia of soil science, 2nd edition, Chap. 1:1. Boca Raton (FL): Taylor & Francis. pp 239–244.
Thompson R, Clark RM. 2008. Is spring starting earlier? Holocene 18:95–104.
Turner JA. 1972. The drought code component of the Canadian Forest Fire Behaviour System. Canadian Forest Service Publication 1316. Ottawa (ON): Environment Canada.
Ueyama M, Harazono Y, Kim Y, Tanaka N. 2009. Response of the carbon cycle in sub-arctic black spruce forests to climate change: reduction of a carbon sink related to the sensitivity of heterotrophic respiration. Agric For Meteorol 149:582–602.
Vaganov E, Shashkin AV. 2000. Growth and tree-ring structure of conifers. Novosibirsk: Nauka Publishing House.
van Cleve K, Dyrness CT, Viereck LA, Fox J, Chapin FI, Oechel WC. 1983a. Taiga ecosystems in interior Alaska. Bioscience 33:39–44.
van Cleve K, Viereck LA, Dyrness CT. 1983b. Productivity and nutrient cycling in taiga forest ecosystems. Can J For Res 13:747–66.
van Cleve K, Oechel WC, Hom JL. 1990. Response of Black Spruce Picea-Mariana ecosystems to soil temperature modification in interior Alaska, USA. Can J For Res 20:1530–5.
Viereck LA, Johnston WF. 1990. Picea mariana (Mill.) B.S.P. black spruce. In: Burns RM, Honkala BH, Eds. Silvics of North America: vol 1. Conifers. USDA For. Serv. Agri. Handb. 654, Washington (DC). pp 227–237.
Wigley TML, Briffa KR, Jones PD. 1984. On the average value of correlated time series, with applications in dendroclimatology and hydrometeorology. J Clim Appl Meteorol 23:201–13.
Yamaguchi DK, Filion L, Savage M. 1993. Relationship of temperature and light ring formation at subarctic treeline and implications for climate reconstruction. Quat Res 39:256–62.
Acknowledgements
Financial support for this research was provided by Industrial Chair in Sustainable Forest Management and a NSERC strategic grant to YB. I.D. thanks Martin Girardin and Remi St-Amant, Natural Resources Canada, for the help with MDC calculation. We thank two anonymous reviewers for helpful comments on an earlier version of MS, Mélanie Desrochers, Université du Québec à Montréal for assistance with digital maps, and Dan McKenney, Canadian Forest Service, for help in obtaining climate data.
Author information
Authors and Affiliations
Corresponding author
Additional information
Author Contributions
ID took a leading role in writing of the text and statistical analyses. MS and AH supplied tree-ring data. ID, MS, YV, and AH discussed the study results during the preparation of the paper.
Rights and permissions
About this article
Cite this article
Drobyshev, I., Simard, M., Bergeron, Y. et al. Does Soil Organic Layer Thickness Affect Climate–Growth Relationships in the Black Spruce Boreal Ecosystem?. Ecosystems 13, 556–574 (2010). https://doi.org/10.1007/s10021-010-9340-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10021-010-9340-7