
Abstract
Riparian ecosystems are recognized as sinks for inorganic nitrogen (N). Denitrification, a heterotrophic microbial process, often accounts for a significant fraction of the N removed. Characteristics of both riparian soils and hydrologic vectors may constrain the locations where denitrification can occur within riparian ecosystems by influencing the distribution of substrates, water, and suitable redox conditions. We employed spatially explicit methods to quantify heterogeneity of soil characteristics and potential rate of denitrification in semi-arid riparian ecosystems. These results allow us to evaluate the relative contributions of hydrologic vectors (water courses that convey materials) and soil resources (materials required by biota) to spatial heterogeneity of denitrification. During dry and monsoon seasons we contrasted a mesic site, characterized by shallow groundwater and annual inundation by floods, with a xeric site that is inundated less often and has a deeper water table. Potential denitrification was detected throughout the mesic floodplain and the average rate of denitrification was greater at the mesic site than at the xeric site, indicating the influence of water availability on denitrification. At the xeric reach, sharp declines in pools of soil resources and rate of denitrification occurred away from the stream, demonstrating the importance of the stream in determining spatial patterns. Using geographically weighted regression analysis, we determined that soil organic matter and soil nitrate were significant predictors of denitrification at the xeric site, but that factors influencing denitrification varied spatially. Spatial heterogeneity of carbon (C) and N substrates in soils likely influenced spatial patterns of denitrification, but distribution of C and N substrates was ultimately organized by hydrologic vectors. Droughts will increase the abundance of reaches with hydrogeomorphic templates similar to the xeric reach studied here. Consequences of such a transition may include a reduced rate of denitrification and patchy distribution of denitrification in floodplain soils, which will decrease the contribution of riparian ecosystems to N removal.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Denitrification, the conversion of nitrate (NO −3 ) to gaseous N2 or N2O by anaerobic heterotrophic bacteria, contributes significantly to maintaining water quality (Seitzinger and others 2006). Riparian ecosystems foster removal of nitrogen (N) from hydrologic flowpaths that connect uplands with downstream ecosystems (Lowrance and others 1984), and denitrification often accounts for a substantial fraction of the N removed (Hanson and others 1994; Vidon and Hill 2004). The interplay of riparian soils with hydrologic vectors influences availability of substrates and redox conditions required for denitrification and may constrain N removal by generating spatial and temporal patterns in the distribution of denitrification activity (Groffman and others 1992; Bruland and others 2006). Improved understanding of interactions among soil characteristics, hydrologic vectors, and denitrification is therefore necessary to estimate the contribution of riparian zones to catchment-scale N budgets, and to design or maintain riparian ecosystems to mitigate N loading. Here we use spatially explicit analyses of riparian floodplain sites that contrast in hydrology to determine how reach-scale hydrology and soil characteristics interact to influence the spatial patterns of denitrification.
Spatial heterogeneity in denitrification may simply be a function of the distribution of available organic C and NO −3 substrates in soils. Localized stores of resources form patches, which are defined as areas that are homogeneous in resource content relative to the surrounding matrix. In contrast, gradual spatial variation in resource content forms gradients. Soil N or organic C may be concentrated near vegetation (Schade and Hobbie 2005), with larger plants corresponding to greater stores of soil organic matter (Sponseller and Fisher 2006), or associated with organic debris (Pettit and Naiman 2005). Vegetation may also contribute to patchiness in soil moisture by promoting infiltration and preventing evaporative water losses (D’Odorico and others 2007). Both of these mechanisms would contribute to patchiness in denitrification. Soil texture may determine capacity for soils to retain C and N (Pinay and others 2000), and spatial variation in texture of riparian soils often occurs along topographic gradients (Bruland and Richardson 2004; Bechtold and Naiman 2006), which would then produce gradual spatial variation in denitrification. Denitrification is also dependent upon redox conditions, which vary spatially in riparian ecosystems (Gallardo 2003) and are related to soil texture, organic matter, or moisture. Therefore, if spatial heterogeneity in substrate availability or redox conditions is the primary influence on spatial patterns in denitrification, then rate of denitrification should be correlated with soil resources in a spatially explicit manner. The nature and strength of such relationships should apply across a range of hydrologic regimes.
The hypothesis that soil characteristics determine the spatial locations of denitrification assumes that all in situ stores of soil resources are accessible, whereas hydrologic vectors may determine where and when soils are biologically active by influencing the distribution of water or by importing missing reagents. Hydrologic vectors are defined here as water courses that convey materials, including water, through an ecosystem. The sum of the characteristics of all hydrologic vectors in an ecosystem defines its hydrologic regime. Hydrologic vectors deliver or redistribute substrates that are required for denitrification (Clement and others 2003; Meixner and others 2007) and may promote other microbial processes (for example, nitrification or decomposition) that generate substrates for denitrification (Seitzinger and others 2006). Inundation of soils by hydrologic vectors influences the oxygen content of soils, promoting the low redox potential required for denitrification (Lamontagne and others 2006). Fluvial sorting of sediments by size may also indirectly influence spatial patterns of denitrification by generating gradients of redox potential and substrate availability in soils (Pinay and others 2000). Hydrologic vectors may thus either increase or decrease spatial heterogeneity of denitrification. We hypothesize that homogenization occurs when hydrologic vectors inundate the whole floodplain or when inundating flows occur frequently. In contrast, hydrologic vectors may contribute to spatial heterogeneity when they are infrequent or unevenly distributed in space. If the hydrologic regime drives spatial distribution of denitrification in floodplains, we predict that spatial patterns will differ between sites that have contrasting hydrologic regimes. In deserts, sharp declines in soil resources and denitrification should occur away from the stream at sites where water tables are deep and floods infrequent, indicating the role of water in facilitating biological activity and promoting biological accumulation of C and N. In contrast, average patch sizes of soils fostering denitrification should be larger where floodplain inundation is more frequent and spatially extensive, or where shallow groundwater is available, reflecting homogenization of soil resources by hydrologic vectors.
Here we report on a study designed to determine the ways in which hydrologic vectors, soil resources, and their interactions influence spatial patterns in denitrification of desert floodplains. We compared spatial patterns along two stream reaches with contrasting hydrologic regimes and assessed relationships among spatial patterns in denitrification, soil resources, and hydrologic vectors to evaluate the hypotheses that (a) spatial patterns in soil resources or (b) characteristics of hydrologic vectors cause variation in the distribution of denitrification in desert floodplains. By observing patterns in dry and monsoon seasons, we were also able to evaluate interactions among spatial patterns in soil resources and hydrologic vectors. We studied a mesic site situated along a gaining reach, with groundwater flow oriented toward the stream channel via shallow flowpaths, that is regularly inundated by floods during the summer monsoon season (Figure 1). A contrasting xeric site was characterized by losing hydrology, with subsurface water routed through deeper flowpaths and overbank floods that regularly inundate only near-stream locations. In dry and monsoon seasons, we evaluated spatial trends with respect to distance from the stream channel, employed geostatistical methods to quantify patch sizes of resource availability and denitrification rate, and quantified the effects of soil resources on rate of denitrification using geographically weighted regression.

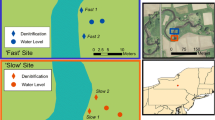
Location of study sites within the upper basin of the San Pedro River. White rectangles highlight locations of the (20 m × 100 m) sampling grids. Arrows indicate flow direction.
Methods
Study Site
The study was conducted within the upper basin (catchment area ∼7,600 km2) of the San Pedro River in southeastern Arizona, USA (Figure 1). The basin is drained by an unregulated stream located near the Sonoran-Chihuahuan desert transition. The study sites lie at approximately 1,200 m elevation and average annual precipitation in the basin ranges 300–750 mm, largely confined to two distinct seasons: winter rains (December–March) and summer monsoon storms (July–September). Weather preceding the monsoon storms is hot and dry (May–July). Two reaches were selected that are bordered by wide (>100 m) floodplains contrasting in hydrologic characteristics. The stream at the xeric site is characterized by losing hydrology and at the mesic site by gaining hydrology (Pool and Coes 1999; Baillie and others 2007). Because stream channels of losing reaches are higher in elevation than the regional groundwater table, water discharges from the stream to the aquifer. The xeric site is thus characterized by ephemeral discharge and dry periods occur in summer prior to onset of floods during the monsoon season, whereas the stream at the mesic site is perennial. During the period of this study, overbank floods did not inundate the entire floodplain at the xeric site. Median discharge at the xeric site was 0 m3/s during the dry season and 1.95 m3/s in the monsoon season (USGS gage # 09471550, ∼7 km downstream of the study site). At the mesic site, median discharge was 0.04 m3/s during the dry season and 2.46 m3/s in the monsoon season (USGS gage # 09471000, ∼10.5 km downstream from the study site).
Both the floodplains are bounded by a mesquite (Prosopis velutina)-dominated terrace situated 3–5 m above the riparian zone. Vegetative communities of the floodplains are dominated by cottonwood (Populus fremontii) and mesquite trees, seepwillow shrubs (Baccharis salicifolia and B. emoryi), native bunchgrass (Sporobolus wrightii), and Johnson grass (Sorghum halepense), a non-native grass. The xeric site also contains rabbitbrush (Ericameria nauseosa), a perennial shrub. Compared to the mesic site, the xeric site supports sparse vegetation cover with fewer cottonwood trees, and obligate wetland species (for example, seepwillow) are confined to near-stream locations. Topography of the xeric floodplain is undulating whereas elevation increases gradually away from the stream at the mesic site (Figure 2). However, absolute change in elevation is similar between the two sites.

Physical and vegetative characteristics of the study sites. Soil texture was measured in the dry season.
Field Sampling
We established one 20 m × 100 m study plot along each reach. Plots were located adjacent and perpendicular to the active channel, extending 100 m into the floodplain (Figure 1). To facilitate geostatistical analysis, a regular grid was overlain with random points to yield 175 sampling locations per reach. The regular grid was spaced at 5 m intervals on the axis parallel to the stream, and random points were established at a density of one point/25 m2. Spacing of the regular grid was chosen to reflect average spacing of perennial trees and shrubs. Soils were sampled twice at each point, once during the dry season and once during the monsoon season.
Soil Analyses
Analysis of soils included available inorganic N, moisture, organic matter, texture, and potential rate of denitrification. Inorganic N in soil was estimated as the ion-exchangeable fraction, which provides an index of available N averaged through time, rather than an instantaneous pool size given by traditional bulk extraction methods (Lajtha 1988). Resin bags were constructed of 4 g mixed-bed ion-exchange resin enclosed in a nylon bag. In May 2006 we buried two replicate resin bags to 5 cm depth at each sampling point. Following 30 days of incubation, we retrieved the resin bags to individual plastic bags and collected one soil sample from each point. Soils were collected manually from within a copper ring (8 cm diameter) inserted to 8 cm depth. Soils and resin bags were transported on ice to the laboratory. Resin bags were then frozen until extraction. Except for texture, analyses of bulk soils were completed within 3 days of collection. Resin bags were again deployed during the monsoon season. Bags were deployed at the mesic site in August and two weeks later at the xeric site. Floods prevented access to the xeric site until this date. Floods again prevented access to both sites until mid-September when resin bags and soil samples were collected. Deposition of sediment at the mesic site resulted in significant loss of resin bags during the monsoon season (∼60%).
Mass of nutrients adsorbed to resins was measured by extraction of resins in 2 M KCl with vigorous shaking for 1 h. Resulting extracts were filtered through pre-leached, ashless Whatman 42 filter papers and frozen until analysis for NH +4 -N (phenol–hypochlorite method) and NO −3 -N (cadmium reduction method) on a Lachat flow-injection autoanalyzer. Analyses of bulk soil were conducted on the smaller-than-2mm fraction. Soil moisture was determined gravimetrically by drying subsamples at 105°C for 48 h. Organic matter content was determined as ash-free dry mass by combustion of subsamples at 550°C for 4 h. The density hydrometer method was used to determine soil texture following dispersion of clay in sodium hexametaphosphate (Robertson and others 1999) and sand content was determined gravimetrically by rinsing subsamples through a 53-μm sieve. Soil texture was analyzed only during the dry season.
Potential rate of denitrification was assayed using the acetylene-block technique (Yoshinari and others 1977). Soils were amended with a solution of dextrose (100 mg dextrose-C l−1) and nitrate (100 mg NO −3 -N l−1). Chloramphenicol (10 mg l−1) was added to prevent new protein synthesis. Resulting slurries were made anoxic by addition of N2 and acetylene was added to approximately 10% v/v headspace. Bottles were then vented to bring headspace equal to atmospheric pressure, shaken manually until all particles were suspended, and initial headspace gas samples were collected into evacuated vials. A final headspace sample was collected after 4 h of incubation, again following vigorous shaking. Resulting gas samples were analyzed for N2O on a gas chromatograph equipped with an electron-capture detector and Porapak-Q columns. Temperature-specific Bunsen coefficients were used to account for N2O in the aqueous portion of slurries.
Vegetation and Topography
Microtopography, canopy height, and canopy intensity were measured by airborne light detection and ranging (LiDAR), a technique that uses travel times of light passing between a source and a target to measure distance. Data were collected via an aircraft in June 2003 at a density of approximately 2–4 points/m2. Points were summarized to produce a grid with a grain size of 1 m2 (Farid and others 2006). Vertical accuracy for elevation and canopy height is 0.15 m. Canopy intensity is described by the normalized return intensity of the LiDAR pulse and may be related to leaf area index (Farid and others 2008). Larger values of return intensity correspond to less light interception by leaves, and presumably smaller leaf area. Values from these raster data were extracted for points at which soil properties were measured.
Statistical Analysis
We used the coefficient of variation (standard deviation/mean) as an index of spatial variability to compare across sites and seasons, interpreting values greater than 100% as indicators of spatial variability. We quantified spatial autocorrelation using analysis of semi-variance. Variograms summarize variance between pairs of points as a function of the separation distances (lags) among measured points. The resulting curve contains information about spatial relationships of the measured points through the nugget (c0), the sill (c), and the range. The nugget, defined as the semi-variance measured at a lag of 0, exists due to residual variance (that is, spatially uncorrelated errors at the measured grain size plus measurement error). The sill occurs when the measured values are no longer spatially correlated and is identified as the semi-variance at which the curve levels off. The range is the lag value associated with the start of the sill and is interpreted here as the mean distance between patch centers, due to periodic nature of the variograms (Radeloff and others 2000). We used the range to compare the spatial distribution of patches supporting denitrification with distribution of soil resources. We quantified the strength of spatial dependence using the ratio of the sill (c) and nugget (c0).
We evaluated the fits of multiple semi-variance models to the data. Akaike’s information criterion (AIC) was used to select the best-fit model. We evaluated models for a lag size of 5 m, across 15 lags. The lag size corresponds to the spacing of the regular grid, and the largest separation distance contains greater than 30 pairs of data points. When necessary, data were logarithmically or arcsine-square root transformed to meet the assumption of normality before fitting semi-variance models. During the dry season, many values of 0 occurred for soil moisture and potential rate of denitrification at the xeric site, so models fit to these data must be interpreted with caution. For all variables, the spherical or Gaussian model best fit the data (Figure 3). The spherical model is defined by

Example variograms fit with spherical (top panel, dry season) and Gaussian models (bottom panel, monsoon season). Data are ln-transformed soil organic matter from the xeric site.
where γ = semi-variance, h = lag size, and a = range. The Gaussian model describes a more gradual rise to the sill, and is defined by
Finally, we used geographically weighted regression (GWR) to determine spatial patterns in the factors that predict potential rate of denitrification. GWR is a spatially explicit regression technique that results in unique regression parameters at each measured location, and significant predictors can be identified for different regions of the study area (Fotheringham and others 2002). Spatial autocorrelation within a dataset is used to determine a bandwidth, the distance over which surrounding locations contribute to the regression function for a given location (here 175 points), and a weighting function determines the contribution of surrounding points to the regression equation. Bandwidth selection was completed by AIC minimization and we applied a fixed bandwidth to all measured locations. Predictor variables included soil moisture, organic matter, texture, and resin-exchangeable nitrate; surface elevation, canopy height, and canopy intensity were also considered. We assessed correlation among predictor variables prior to regression analyses and did not include collinear predictors in the same model (correlation coefficient >0.4). We used normal probability plots of residuals to assess normality and maps of residuals to assess spatial constancy of variance. Potential rate of denitrification was logarithmically transformed to meet the assumption of normality. We compared the ordinary least squares (OLS) model (that is, global model) with each corresponding GWR model (that is, local model) using an F-test of error variance. We included only those GWR models that explained significantly greater variation than the OLS models as candidate models. We used a Monte Carlo approach to determine whether predictors of denitrification were significantly heterogeneous in space. The spatial coordinates of the variance of each partial regression coefficient were randomly permuted to generate a distribution of variance estimates. Significance was determined by comparing the actual variance against the distribution of permuted estimates. Finally, we use plots of the t-values corresponding to partial regression coefficients to visualize spatial patterns in predictors of denitrification. Because we use these plots to assess spatial patterns in important predictors rather than to make statistical inferences about any particular location, we present t-values that are uncorrected for multiple tests. All regressions were completed using GWR 3.0 software (Fotheringham and others 2002, National University of Ireland).
For each season, candidate GWR models were ranked based on AIC weights (w i ), which are interpreted as the probability that a given model is the best fit of the models considered. A 95% confidence set of models was constructed by summing w i across models to total 0.95 (Burnham and Anderson 2002). Weights were calculated using the difference between the AIC value of each model and the model with the smallest AIC (Δ i ), normalized by the sum of Δ i across the set of candidate models (Burnham and Anderson 2002).
Results
Seasonal and Site Comparisons
Soils of the mesic site supported greater potential denitrification and tended to be more spatially homogeneous than those of the xeric site. At the mesic site, soils were finer in texture, containing no more than 50% sand; in contrast, the coarser soils of the xeric site contained no less than 50% sand (Table 1). Soils of the mesic site contained several-fold more moisture and organic matter than those of the xeric site whereas the xeric site contained greater inorganic N (Table 1). Median potential rate of denitrification was an order of magnitude greater at the mesic compared to xeric site in both seasons and denitrification was more spatially variable at the xeric site, with many points that showed no denitrification activity (Table 1).
Soil moisture and potential rate of denitrification increased at both sites following monsoon storms. Median soil moisture content and potential rate of denitrification increased 100-fold at the xeric site and by several fold at the mesic site following monsoon season floods. At the mesic site, soil organic matter content decreased following the monsoon season and we observed significant deposits of sediment throughout the site (∼20 cm depth, on average). Declines in soil organic matter content following monsoon floods occurred only within 10 m of the stream channel at the xeric site. Total pool sizes of resin-available inorganic N were similar before and after the monsoon season at the xeric site, although some locations exhibited 10-fold increases in NO −3 . Ammonium remained the dominant form of inorganic N following the monsoon season at the xeric site. Total inorganic N declined at the mesic site, and NO −3 became the dominant form.
Distance from Stream Patterns
Soil and vegetation characteristics exhibited spatial patterns with respect to distance from the stream at the xeric, but not the mesic site. At the xeric site, canopy height (Figure 2), moisture, and potential rate of denitrification (Figures 4 and 5) were highest near the stream and moderate values occurred near the border with the upland terrace. For most characteristics, a region of low values occurred approximately 50–70 m from the stream at the xeric site, corresponding with soils of nearly 100% sand (Figure 2). These patterns were apparent during both seasons. Exchangeable NO −3 also showed a similar spatial pattern during the monsoon season (Figure 5). Spatial patterns in soil characteristics were not observed relative to the stream at the mesic site (Figures 6 and 7).

Spatial arrangement of soil characteristics at the xeric site measured in the dry season.

Spatial arrangement of soil characteristics at the xeric site measured in the monsoon season.

Spatial arrangement of soil characteristics at the mesic site measured in the dry season.

Spatial arrangement of soil characteristics at the mesic site measured in the monsoon season. Fewer replicates are available for soil NO –3 due to resin bag loss in monsoon season floods.
Spatial Autocorrelation
A substantial fraction of variation in soil characteristics was explained by spatial autocorrelation at the xeric site, as shown by the nugget:sill ratio (Table 2), with the exception of inorganic N. At the xeric site, soil characteristics that could be modeled were best fit by a spherical model during the dry season. Following the monsoon season, spatial variation in organic matter and denitrification was best fit by a Gaussian model, which describes a smoother spatial pattern than the spherical model (Figure 3). Values of the range parameter (that is, the distance between patches) for denitrification were intermediate to range parameters for soil characteristics in both seasons (Table 2). Soils at the mesic site were spatially homogeneous and could not be modeled using analysis of semi-variance.
Spatial Heterogeneity in Predictors of Denitrification
At the xeric site, predictors of potential rate of denitrification were spatially and seasonally variable (Table 3; Figure 8). In the dry season, only soil organic matter was significantly related to denitrification, and this relationship occurred for several distinct portions of the study area (Figure 8). In the monsoon season, the 95% confidence set contained several GWR models. Organic matter was a significant predictor, but available NO −3 , canopy elevation, canopy intensity, and topography also explained significant portions of the variation in rate of denitrification (Table 3, Figure 8). All GWR models for the xeric site explained significantly more variation than OLS counterparts, indicating that factors influencing denitrification were spatially heterogeneous. At the mesic site, OLS models explained no more than 10% of the variation in potential rate of denitrification. Further, GWR models did not explain more variation in denitrification than corresponding OLS models and regression results are thus not presented for the mesic site.

T-values of partial regression coefficients in geographically weighted regression models at the xeric site. Best-fit models for the dry season (left panel) and monsoon season (right panel) are shown. Maps are oriented with the stream near the bottom edge of the figure, and the terrace near the top. NS designates no significant predictors.
Discussion
Using comparisons of two representative desert floodplains, we found support for the hypothesis that soil resources determine the spatial locations of denitrification activity. We also found support for the hypotheses that the hydrologic regime influences distribution of denitrification activity directly by constraining water availability and indirectly by distributing substrates. The mesic floodplain, which is more frequently inundated by floods and has shallower groundwater, contained greater soil moisture, organic matter, vegetation, and higher potential rate of denitrification than the xeric site, indicating a direct influence of water on soil resources and denitrification. However, floodplain soils of the mesic site showed no spatial pattern at the scales measured, suggesting that overbank floods and availability of shallow groundwater result in homogenization of resource pools. In contrast, distribution of resources and denitrification activity were spatially heterogeneous at the xeric site, and spatial patterns were likely due to existing hydrologic vectors and the legacies of past floods.
Whole Reach Patterns
The direct effect of water availability on potential rate of denitrification at the reach scale was evidenced by seasonal changes within the sites, and by contrasts between the mesic and xeric sites. Median potential rate of denitrification increased 10-fold at both reaches following the monsoon season. Soil moisture, organic matter, canopy height, and potential rate of denitrification were also several-fold greater at the mesic compared to the xeric site. Results emphasizing the strong influence of water availability on denitrification are certainly not unique. For example, Pinay and others (2007) determined that across floodplains of Europe, soil moisture was the strongest predictor of denitrification. In the semi-arid basin studied here, potential rate of denitrification responded to patterns in water availability caused by both seasonality and the reach-scale hydrologic regime, supporting the hypothesis that hydrologic vectors cause variation in denitrification via influences on water availability.
Within-Site Heterogeneity
We predicted that patch size of soil characteristics would be larger at the mesic compared to xeric site due to the homogenizing effects of overbank floods. However, at the grain size of our sampling, we could not detect patches at the mesic site. Soils showed no clear trends in any characteristic with distance from the stream, and soil characteristics produced non-significant variograms, indicating lack of spatial autocorrelation. Lack of spatial pattern at the mesic site supports the hypothesis that hydrologic vectors influence denitrification by distributing resources, and is likely due to the large spatial extent of inundation by floods, high recurrence interval of floods, and a shallow water table. The notion that the mesic site is homogenized due to continual reworking by floods is supported by seasonal changes in soil organic matter content. We observed a decrease in soil organic matter content at the mesic site following inundation by monsoon floods. Concurrent with our field observations of sediment deposition, this result suggests that materials of low organic content were deposited during floods. Repeated inundation over the course of a season increases the likelihood that a particular location will receive flood deposits, which could contribute to the spatial homogeneity observed here. Indeed, comparison of plots of organic matter with distance from the stream for both seasons show that decreases in soil organic matter content occurred throughout the mesic site (Figures 6, 7).
Observations in other floodplains have shown that the effects of flooding may supercede the effects of local soil characteristics on microbial processes (Johnston and others 2001) and that recurrent floods may increase the distance over which spatial dependence is observed (Gallardo 2003). The size of dominant plants is often positively correlated with patch sizes of soil characteristics (Gross and others 1995; Gallardo and others 2000), but at the mesic site, large cottonwoods form a closed canopy over a large portion of the site. This likely results in broad spatial distribution of plant-derived substrates, and may have contributed to homogeneity at the grain and extent at which we sampled. Thus floodplains with high flood frequency and vegetation cover may be generally associated with broadly distributed denitrification activity. Finally, spatial homogeneity at the grain and extent measured does not preclude heterogeneity at finer scales. In estuarine stream banks, microbial processes that are influenced by drainage and redox potential varied at a smaller scale than processes that were more strongly influenced by distance from the stream (Franklin and others 2002), suggesting that if distance from the stream is not influencing spatial trends at the mesic site, perhaps spatial autocorrelation in denitrification occurs in accordance with microsite-scale variation in soil redox potential.
At the xeric site, spatial patterns in potential rate of denitrification were correlated with soil and vegetation characteristics, but hydrologic vectors played a dominant role in determining capacity for denitrification. Analysis of semi-variance indicated that the mean distance between patches supporting denitrification activity was intermediate to the distance between patches of high relative soil moisture and organic matter during both seasons, and similar to that for fine particles measured in the dry season. Moreover, correlations between patch attributes and potential rate of denitrification were spatially explicit, indicating heterogeneity in the relationships among denitrification and patch attributes or their common drivers. Given the demonstrated significance of hydrology at seasonal, reach, and within-reach scales, it is surprising that soil moisture was not a significant predictor of potential rate of denitrification. This may be a result of assessing denitrification activity in saturated slurries. Alternatively, it indicates that at the scales considered, hydrologic vectors influence denitrification primarily by establishing patterns in availability of nitrate and organic C substrates, rather than by the direct influence of water on redox potential of soils. Spatially heterogeneous correlations of denitrification and predictor variables suggest that use of easily measured or remotely sensed attributes to predict and scale up floodplain denitrification capacity must account for spatial patterns. Aerial imagery may be used to delineate regions of study sites that vary in flood history or vegetation cover, with these data used to guide spatially stratified soil sampling.
Soil moisture, vegetation characteristics, and potential rate of denitrification declined with distance from the stream at the xeric site, indicating the link between floodplain soils and stream water availability and supporting the hypothesis that hydrologic vectors influence spatial patterns of denitrification. A seasonal shift in the model best capturing semi-variance at the xeric site further implicates the contributions of hydrologic processes to spatial patterns in the floodplain. All soil characteristics modeled during the dry season were best fit by a spherical model, which describes a rapid decline in spatial autocorrelation over increasing lag distances, indicating patchiness. Following the monsoon season, spatial patterns of organic matter and rate of denitrification were best fit by a Gaussian model, which describes a more gradual pattern. Redistribution of resources by fluvial processes in the near-stream zone, the only portion of the xeric floodplain inundated by floods during the monsoon season, may explain the seasonal transition from patchy to smooth variation.
The gradient pattern associated with location of the channel was accompanied by a patchy spatial pattern that encompassed the entire study area. A distinctive band of coarse-textured soils (nearly 100% sand) occupied a zone 50–70 m from the stream, and was indicative of alluvial deposits. This zone was characterized by sparse vegetation, the lowest soil organic matter content observed at the site, and was also a denitrification “cold spot”, with rates substantially lower than surrounding areas of the floodplain. We observed an increase in soil resources, potential rate of denitrification, and canopy height, and a decrease in soil particle size on both sides of this zone. Analysis of semi-variance validated existence of two distinct patches, with periodic variograms resulting from all measured characteristics. Periodic variograms result where only a few patches occupy a study area, such as the two patches observed here (Radeloff and others 2000). The range parameter by definition corresponds to the coarsest scale comprising a spatial pattern (Meisel and Turner 1998), giving us confidence that our analysis captured the dominant spatial pattern. Locations of the two patches supporting denitrification activity in this desert floodplain are consistent with observations in mesic regions (Schipper and others 1993; McClain and others 1994). Stream-riparian and upland-riparian interfaces are thought to support denitrification by providing complementary overlap in the conditions supporting this process, including redox gradients (Hedin and others 1998) and supplies of NO –3 or organic C (Devito and others 2000).
Feedbacks Between Hydrology and Spatial Patchiness
Feedbacks between water, vegetation, and sediment transport may explain how observed patchiness in denitrification is created and maintained. In ecosystems subject to overland flow, vegetation traps fine particles transported by hydrologic vectors, entrainment results in increased nutrient and water retention, and these resources in turn promote plant recruitment and growth, creating a positive feedback (Corenblit and others 2007). The legacy of past floods may have greater influence on rates of denitrification at the xeric site because legacy effects are often strongest in landscapes that are spatially heterogeneous or when frequency of disturbance is low (Peterson 2002). In the absence of floods, shallow groundwater is accessible to most plants only near the stream channel along losing reaches, and supports recruitment and growth of cottonwood trees (Amlin and Rood 2002; Snyder and Williams 2007). Trees positively influence soil moisture via shading and production of abundant organic material. In addition, mesquite, a tree that can access deep groundwater, was abundant near the terrace at the xeric site. Presence of mesquite may contribute to the slight increase in resource availability near the terrace via similar mechanisms as cottonwoods near the stream, but mesquite trees additionally perform hydraulic lift during droughts, which may result in an additional source of water to surface soils in this portion of the floodplain during the dry season (Scott and others 2008). Stands of cottonwood and mesquite trap fine sediments during floods (Hupp and Osterkamp 1996), but require periods of low flow to become established (Stromberg and others 2007). Thus when floods are infrequent, spatial patterns in recruitment of vegetation resulting from a flood could lead to patterns in productivity and soil resources if large floods are absent in subsequent years, allowing plants to establish. Maintenance or amplification of spatial patterns may result from subsequent floods by positive feedback whereby collection of fine particles in vegetated areas improves conditions for growth. In contrast, at the mesic site, two hydrologic vectors—high frequency overbank floods and shallow groundwater—transport resources throughout the floodplain and likely support dense, productive vegetation. Recurrence of multiple overbank floods each year and shallow groundwater that is accessible to plants decreases the probability that spatial patterns created by a single event will persist. Such rapid, small-scale changes in the locations that can support denitrification diminish the appearance of spatial patterns and may be common in floodplains that are inundated over relatively long time periods (Orr and others 2007).
We conclude that for these representative floodplains of the San Pedro River, soil resources may be the proximate drivers of spatial pattern in rate of denitrification, but that spatial patterns are ultimately created and maintained by hydrologic vectors. Understanding how hydrology influences spatial heterogeneity of denitrification within riparian ecosystems is important for predicting changes in ecosystem function under altered climatic conditions and scenarios of human water use. Our observations suggest that temporal patterns in discharge, flood history, and vegetative cover may be used to indicate degree of spatial heterogeneity and guide efforts to characterize denitrifying capacity of floodplains. Arid regions of the world are increasing in area (Dai and others 2004) and depth to groundwater, instream flows, and high flow events are decreasing in many regions (Jackson and others 2001; Poff and others 2006) due to droughts and water withdrawals. Such hydrologic changes will reduce water availability and substrate delivery to riparian ecosystems, and may cause declines in abundance of vegetation (Stromberg and others 2007; Rood and others 2008). Furthermore, climate change models for the Southwest point to an increasingly warm and dry future (Seager and others 2007). If decreases in water availability to floodplains result in riparian ecosystems that function more like the xeric than the mesic site studied here, the result may be a lower average rate of denitrification and confinement of denitrification to a fraction of total riparian spatial extent, reducing the overall capacity for floodplains to contribute to N removal and maintenance of water quality.
References
Amlin N, S Rood. 2002. Comparative tolerances of riparian willows and cottonwoods to water-table decline. Wetlands 22:338–46.
Baillie MN, Hogan JF, Ekwurzel B, Wahi AK, Eastoe CJ. 2007. Quantifying water sources to a semiarid riparian ecosystem, San Pedro River, Arizona. J Geophys Res-Biogeosci 112:G03S02. doi:10.1029-2006JG000263
Bechtold JS, RJ Naiman. 2006. Soil texture and nitrogen mineralization potential across a riparian toposequence in a semi-arid savanna. Soil Biol Biochem 38:1325–33.
Bruland GL, CJ Richardson. 2004. A spatially explicit investigation of phosphorus sorption and related soil properties in two riparian wetlands. J Environ Qual 33:785–94.
Bruland GL, CJ Richardson, SC Whalen. 2006. Spatial variability of denitrification potential and related soil properties in created, restored, and paired natural wetlands. Wetlands 26:1042–56.
Burnham KP, DR Anderson. 2002. Model selection and multimodel inference: a practical information-theoretical approach. 2nd edn. New York: Springer-Verlag.
Clement J-C, L Aquilina, O Bour, K Plaine, TP Burt, G Pinay. 2003. Hydrological flowpaths and nitrate removal rates within a riparian floodplain along a fourth-order stream in Brittany (France). Hydrol Process 17:1177–95.
Corenblit D, E Tabacchi, J Steiger, AM Gurnell. 2007. Reciprocal interactions and adjustments between fluvial landforms and vegetation dynamics in river corridors: a review of complementary approaches. Earth Sci Rev 84:56–86.
Dai A, K Trenberth, T Qian. 2004. A global dataset of Palmer drought severity index for 1870–2002: relationship with soil moisture and effects of surface warming. J Hydrometeorol 5:1117–30.
Devito KJ, D Fitzgerald, AR Hill, R Aravena. 2000. Nitrate dynamics in relation to lithology and hydrologic flow path in a river riparian zone. J Environ Qual 29:1075–84.
D’Odorico P, Caylor K, Okin GS, Scanlon TM. 2007. On soil moisture-vegetation feedbacks and their possible effects on the dynamics of dryland ecosystems. J Geophys Res Biogeosci 112:G04010. doi:10.1029/2006JG000379
Farid A, DC Goodrich, S Sorooshian. 2006. Using airborne lidar to discern age classes of cottonwood trees in a riparian area. West J Appl For 21:149–58.
Farid A, DC Goodrich, R Bryant, S Sorooshian. 2008. Using airborne lidar to predict leaf area index in cottonwood trees and refine riparian water-use estimates. J Arid Environ 72:1–15.
Fotheringham AS, C Brunsdon, M Charlton. 2002. Geographically weighted regression. West Sussex, England: Wiley.
Franklin RB, LK Blum, AC McComb, AL Mills. 2002. A geostatistical analysis of small-scale spatial variability in bacterial abundance and community structure in salt marsh creek bank sediments. FEMS Microbiol Ecol 42:71–80.
Gallardo A. 2003. Spatial variability of soil properties in a floodplain forest in Northwest Spain. Ecosystems 6:564–76.
Gallardo A, JJ Rodriguez-Saucedo, F Covelo, R Fernandez-Ales. 2000. Soil nitrogen heterogeneity in a Dehesa ecosystem. Plant Soil 222:71–82.
Groffman PM, AJ Gold, RC Simmons. 1992. Nitrate dynamics in riparian forests: microbial studies. J Environ Qual 21:666–71.
Gross KL, KS Pregitzer, AJ Burton. 1995. Spatial variation in nitrogen availability in 3 successional plant-communities. J Ecol 83:357–67.
Hanson GC, PM Groffman, AJ Gold. 1994. Denitrification in riparian wetlands receiving high and low groundwater nitrate inputs. J Environ Qual 23:917–22.
Hedin LO, JC von Fischer, NE Ostrom, BP Kennedy, MG Brown, GP Robertson. 1998. Thermodynamic constraints on nitrogen transformations and other biogeochemical processes at soil-stream interfaces. Ecology 79:684–703.
Hupp C, W Osterkamp. 1996. Riparian vegetation and fluvial geomorphic processes. Geomorphology 14:277–95.
Jackson RB, SR Carpenter, CN Dahm, DM McKnight, RJ Naiman, SL Postel, SW Running. 2001. Water in a changing world. Ecol Appl 11:1027–45.
Johnston CA, SD Bridgham, JP Schubauer-Berigan. 2001. Nutrient dynamics in relation to geomorphology of riverine wetlands. Soil Sci Soc Am J 65:557–77.
Lajtha K. 1988. The use of ion-exchange resin bags for measuring nutrient availability in an arid ecosystem. Plant Soil 105:105–11.
Lamontagne S, FW Leaney, AL Herczeg. 2006. Patterns in groundwater nitrogen concentration in the riparian zone of a large semi-arid river (River Murray, Australia). River Res Appl 22:39–54.
Lowrance R, R Todd, J Fail, O Hendrickson, R Leonard, L Asmussen. 1984. Riparian forests as nutrient filters in agricultural watersheds. Bioscience 34:374–7.
McClain M, J Richey, T Pimentel. 1994. Groundwater nitrogen dynamics at the terrestrial-lotic interface of a small catchment in the central Amazon Basin. Biogeochemistry 27:113–27.
Meisel JE, MG Turner. 1998. Scale detection in real and artificial landscapes using semivariance analysis. Landsc Ecol 13:347–62.
Meixner T, Huth AK, Brooks PD, Conklin MH, Grimm NB, Bales RC, Haas PA, Petti JR. 2007. Influence of shifting flow paths on nitrogen concentrations during monsoon floods, San Pedro River, Arizona. J Geophys Res Biogeosci 112:G03S03
Orr CH, E Stanley, K Wilson, J Finlay. 2007. Effects of restoration and reflooding on soil denitrification in a leveed midwestern floodplain. Ecol Appl 17:2365–76.
Peterson G. 2002. Contagious disturbance, ecological memory, and the emergence of landscape pattern. Ecosystems 5:329–38
Pettit N, R Naiman. 2005. Flood-deposited wood debris and its contribution to heterogeneity and regeneration in a semi-arid riparian landscape. Oecologia 145:434–44.
Pinay G, VJ Black, AM Planty-Tabacchi, B Gumiero, H Decamps. 2000. Geomorphic control of denitrification in large river floodplain soils. Biogeochemistry 50:163–82.
Pinay G, B Gumiero, E Tabacchi, O Gimenez, AM Tabacchi-Planty, MM Hefting, TP Burt, VA Black, C Nilsson, V Iordache, F Bureau, L Vought, GE Petts, H Decamps. 2007. Patterns of denitrification rates in European alluvial soils under various hydrological regimes. Freshw Biol 52:252–66.
Poff N, B Bledsoe, C Cuhaciyan. 2006. Hydrologic variation with land use across the contiguous United States: geomorphic and ecological consequences for stream ecosystems. Geomorphology 79:264–85.
Pool DR, Coes AL. 1999. Hydrogeologic investigations of the Sierra Vista subwatershed of the upper San Pedro basin, Cochise County, southeast Arizona. In: USGS Water Resources Investigations Report
Radeloff VC, TF Miller, HS He, DJ Mladenoff. 2000. Periodicity in spatial data and geostatistical models: autocorrelation between patches. Ecography 23:81–91.
Robertson GP, Coleman D, Bledsoe C, Sollins P. 1999. Standard soil methods for long-term ecological research. USA: Oxford University Press
Rood S, J Pan, K Gill, C Franks, G Samuelson, A Shepherd. 2008. Declining summer flows of Rocky Mountain rivers: changing seasonal hydrology and probable impacts on floodplain forests. J Hydrol 349:397–410.
Schade JD, SE Hobbie. 2005. Spatial and temporal variation in islands of fertility in the Sonoran Desert. Biogeochemistry 73:541–53.
Schipper LA, AB Cooper, CG Harfoot, WJ Dyck. 1993. Regulators of denitrification in an organic riparian soil. Soil Biol Biochem 25:925–33.
Scott RL, WL Cable, KR Hultine. 2008. The ecohydrologic significance of hydraulic redistribution in a semiarid savanna. Water Resour Res 44:W02440.
Seager R, MF Ting, I Held, Y Kushnir, J Lu, G Vecchi, HP Huang, N Harnik, A Leetmaa, NC Lau, CH Li, J Velez, N Naik. 2007. Model projections of an imminent transition to a more arid climate in southwestern North America. Science 316:1181–4.
Seitzinger S, JA Harrison, JK Bohlke, AF Bouwman, R Lowrance, B Peterson, C Tobias, G Van Drecht. 2006. Denitrification across landscapes and waterscapes: a synthesis. Ecol Appl 16:2064–90.
Snyder K, D Williams. 2007. Root allocation and water uptake patterns in riparian tree saplings: responses to irrigation and defoliation. For Ecol Manage 246:222–31.
Sponseller R, S Fisher. 2006. Drainage size, stream intermittency, and ecosystem function in a Sonoran Desert landscape. Ecosystems 9:344–56.
Stromberg JC, VB Beauchamp, MD Dixon, SJ Lite, C Paradzick. 2007. Importance of low-flow and high-flow characteristics to restoration of riparian vegetation along rivers in and south-western United States. Freshw Biol 52:651–79.
Vidon P, AR Hill. 2004. Denitrification and patterns of electron donors and acceptors in eight riparian zones with contrasting hydrogeology. Biogeochemistry 71:259–83.
Yoshinari T, R Hynes, R Knowles. 1977. Acetylene inhibition of nitrous-oxide reduction and measurement of denitrification and nitrogen-fixation in soil. Soil Biol Biochem 9:177–83.
Acknowledgments
We thank R. Davies, R. Martin, M. McCrackin, and S. Norlin for assistance with data collection. LiDAR data were provided by the USDA-ARS, Tucson, AZ, and the University of Florida with funding assistance from the EPA, and the U.S. Dept. of Defense Legacy Program. A. Farid supplied processed LiDAR data and A. Buyantuyev assisted with GIS. Comments from J. Elser, S. Fisher, S. Hall, J. Sabo, and two anonymous reviewers improved the quality of this manuscript. An Ariel Appleton research grant from the Research Ranch Foundation (to TKH) supported this study.
Author information
Authors and Affiliations
Corresponding author
Additional information
TKH designed and completed the study and wrote the paper; EAW contributed methods and edited the paper; NBG designed the study and edited the paper.
Rights and permissions
About this article
Cite this article
Harms, T.K., Wentz, E.A. & Grimm, N.B. Spatial Heterogeneity of Denitrification in Semi-Arid Floodplains. Ecosystems 12, 129–143 (2009). https://doi.org/10.1007/s10021-008-9212-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10021-008-9212-6