
Abstract
Wildfire can influence a variety of stream ecosystem properties. We studied stream temperatures in relation to wildfire in small streams in the Boise River Basin, located in central Idaho, USA. To examine the spatio-temporal aspects of temperature in relation to wildfire, we employed three approaches: a pre–post fire comparison of temperatures between two sites (one from a burned stream and one unburned) over 13 years, a short-term (3 year) pre–post fire comparison of a burned and unburned stream with spatially extensive data, and a short-term (1 year) comparative study of spatial variability in temperatures using a “space for time” substitutive design across 90 sites in nine streams (retrospective comparative study). The latter design included streams with a history of stand-replacing wildfire and streams with severe post-fire reorganization of channels due to debris flows and flooding. Results from these three studies indicated that summer maximum water temperatures can remain significantly elevated for at least a decade following wildfire, particularly in streams with severe channel reorganization. In the retrospective comparative study we investigated occurrence of native rainbow trout (Oncorhynchus mykiss) and tailed frog larvae (Ascaphus montanus) in relation to maximum stream temperatures during summer. Both occurred in nearly every site sampled, but tailed frog larvae were found in much warmer water than previously reported in the field (26.6°C maximum summer temperature). Our results show that physical stream habitats can remain altered (for example, increased temperature) for many years following wildfire, but that native aquatic vertebrates can be resilient. In a management context, this suggests wildfire may be less of a threat to native species than human influences that alter the capacity of stream-living vertebrates to persist in the face of natural disturbance.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Wildfire is a major source of natural disturbance in stream ecosystems (McMahon and deCalesta 1990; Minshall and others 1997; Gresswell 1999). Some of the most dramatic disturbances from wildfire involve post-fire flooding and debris flows. These events completely reorganize channels by redistributing massive amounts of sediment and wood that scour and fill different portions of stream channel networks (Benda and others 2003; Meyer and Pierce 2003; Miller and others 2003; Wondzell and King 2003). In the short-term, these events can lead to massive mortality of benthic macroinvertebrates (Minshall 2003), amphibians (Pilliod and others 2003; Bury 2004), and fishes (Rieman and others 1997; Burton 2005). Over time, stream biota can recover and some responses of benthic macroinvertebrates suggest that community structure begins to stabilize within 7–10 years following wildfire (Minshall 2003). The rate and nature of recovery of fishes and amphibians is likely dependent on characteristics of the species and the disturbed habitat (Dunham and others 2003; Pilliod and others 2003). Wildfire is a natural process and perhaps less of a threat in unmanaged ecosystems (for example, wilderness areas), but the vulnerability of many species to fire-related disturbances has increased due to human influences on aquatic vertebrates and altered fire regimes (Minshall 2003).
Although some groups of organisms appear to have the capacity to recover within several years of wildfire and related disturbances, the extent, magnitude, and duration of many important physical changes to stream environments and their specific influences on stream biota are still poorly understood (Gresswell 1999; Spencer and others 2003; Minshall 2003). One of the most obvious physical changes that may occur in streams is altered water temperature (Beschta and others 1987; Moore and others 2005a). During wildfire, intense heating can lead to short-term (<1 day) increases in stream temperatures of several degrees (for example, Hitt 2003). Over longer timeframes and broader spatial scales, changes to terrestrial vegetation and stream channel morphology related to wildfire may lead to substantially altered water temperatures across stream networks, leading to loss and fragmentation of suitable habitats and increased risk of local extinction for coldwater aquatic species (Dunham and others 2003; Pilliod and others 2003; Bury 2004; Burton 2005; Sestrich 2005).
Wildfire and associated channel disturbance may influence a number of processes that contribute to the heat budget of streams, and a large number of thermal responses are possible (Beschta and others 1987; Johnson 2003; Moore and others 2005a). Increased stream temperatures are likely following fire-related loss of shading streamside vegetation and concomitant increases in solar radiation (Brown and Krieger 1970; Dwire and Kauffman 2003). Alternatively, loss of vegetation may also cause summer base flows to increase slightly (Ziemer 1964; Jones and Post 2004), possibly leading to reduced rates of heating. However, any effect from this decrease in heating rate may be offset by an increase in the size of the low flow stream network, resulting in a longer total length of stream exposed to solar radiation. In addition, channels may be wider and more exposed to solar radiation following channel reorganizing events (Swanson 1981), but localized changes in substrate characteristics can moderate temperatures through surface–subsurface heat exchange (Johnson and Jones 2000). Overall, it appears that habitat alterations resulting from wildfire and channel disturbance (that is, loss of shading vegetation, channel widening) will result in a net increase in stream temperatures despite other associated habitat alterations that could reduce heating rates (that is, increased base flow, surface–subsurface heat exchange).
In this study, we examined headwater stream temperatures in relation to wildfire and channel disturbance history in central Idaho. Our primary goal in this work was to better understand spatial and temporal changes in stream temperatures across a fire-prone landscape and to relate those changes to the distribution of sensitive species. To this end, we adopted a spatially extensive sampling design to study patterns of stream temperatures among sites within several streams with contrasting wildfire and physical disturbance histories. Previous work in other systems provided evidence for increased stream temperatures following wildfire (for example, Minshall and others 1997), but the influence of site-specific variation (Dunham and Vinyard 1997) was not controlled among streams. To account for site variation, we compared thermal variability within and among streams in relation to wildfire and physical disturbance histories. This retrospective spatial analysis was complemented with long-term information from a fortuitous collection of samples over 3–13 years that contrasted pre- and post-fire temperatures in burned and unburned streams. Finally, we examined the distribution of a coldwater guild of aquatic vertebrates (salmonid fishes and amphibians) among sites and streams to determine if observed temperature changes associated with wildfire influenced patterns of species occurrence. Collectively, these results provide a broad complement of spatial and temporal perspectives on post-fire patterns of stream heating and potential threats to coldwater aquatic vertebrates.
Methods
Study Area
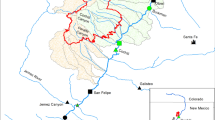
This study was conducted in the Boise River Basin, located in central Idaho (Figure 1). The Boise River Basin is part of the Idaho batholith region, consisting of highly erodible granite rock and steep topography. Precipitation ranges from about 600 to 1,000 mm per year, with greater amounts at higher elevations. Most precipitation falls in the winter as snow, with occasional winter or spring storms that deliver warm rains on snow that can lead to flooding. Localized flooding can also result from convective storms in summer (Miller and others 2003). Climate in the basin is typical of the region with cold, wet winters and freezing temperatures contrasting with generally hot and dry summers and air temperatures commonly exceeding 30°C.

Shaded elevation map of Boise River Basin indicating blue stream lines and sites sampled for stream temperatures and vertebrates in nine streams during the summer of 2003 (closed circles), sites sampled for stream temperatures for summers of 1993–2005 (Cottonwood Creek and Roaring River; open circles), and sites sampled for stream temperatures for the summers of 2002 and 2004 (James and Phifer Creeks; open stars). Wildfires occurring in different years are indicated by outlines around fire perimeters (black outline 1992, white line 1994, green line 2000, purple line 2003). Colors within fire perimeters indicate the severity of wildfires determined by the Boise National Forest, with darkest red corresponding to high severity wildfire, light pink corresponding to low severity wildfire, and an intermediate shade of red corresponding to moderate severity wildfire. Note the spatial resolution of fire severity mapping was greater in 2000 and 2003.
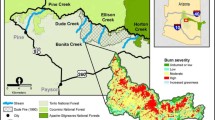
Riparian tree species consisted primarily of willow (Salix, spp.), cottonwood or aspen (Populus, spp.), alder (Alnus, spp.), and birch (Betula, spp.). Hillslope vegetation within study streams ranges from subalpine fir (Abies lasiocarpa) at high elevations to mixed conifer and Ponderosa pine (Pinus ponderosa) forest and shrubsteppe at low elevations, with the latter becoming more prevalent on south-facing slopes. Lower elevation vegetation is characterized by mixed severity fires with return intervals of less than 35 years, whereas expected intervals for higher elevation forests may be an order of magnitude greater (Brown and Smith 2000). Since 1992, several wildfires have burned 95,000 ha within the 217,000 ha upper Middle Fork Boise River Basin upstream of Arrowrock Reservoir, a dramatic shift in fire behavior from a relatively wildfire-free period characterizing most of the twentieth century. Within wildfire perimeters, burn severity was mapped by the Boise National Forest using three categories: (1) high severity—corresponding to stand-replacing fires; (2) moderate severity—corresponding to fires that burn understory vegetation with occasional burning of individual or small groups of trees; (3) low severity—corresponding to fires consuming only understory vegetation or forest litter (Figure 1; Burton 2005).
Stream Temperature Sampling
Pre- and Post-fire Comparisons
We compared two fortuitous samples of stream temperatures before and after wildfires had occurred (Figure 1). In one case, summer (15 July–15 September) stream temperatures recorded in 1993 at two sites in different streams (Cottonwood Creek and Roaring River) were compared to each other following a wildfire in one of the streams (Cottonwood Creek) in 1994. Sampling was continued on an intermittent yearly basis through 2005. In a second case, summer (15 July–15 September) stream temperatures recorded in 2002 at ten sites in two streams (James and Phifer Creeks) were contrasted with temperature samples taken the summer of 2004, following a severe wildfire in one of the streams in 2003 (James Creek; Figure 1). Water temperatures were sampled with Hobo™ and Tidbit™ dataloggers (Onset Computer Corporation, Pocasset, MA, USA) before and after 2002, respectively. Field deployment of temperature dataloggers followed guidelines described in Dunham and others (2005).
Retrospective Study
For a retrospective comparison of stream temperatures in relation to disturbance history, we selected streams in three major disturbance categories (Figure 2): (1) streams without recent stand-replacing wildfire in their watersheds; (2) streams with a recent history of moderate-to-high severity wildfire predominant in the watershed; (3) streams in watersheds with a recent history of moderate-to-high severity wildfire that were followed by massive channel reorganization from a debris flow or severe flood. Such events can occur independently of wildfire, but are much more likely after a wildfire occurs (Wondzell and King 2003). We selected streams within the three categories to contrast the effects of wildfire and fire-related channel disturbance with streams having little recent history of disturbance. Many stream catchments within the Boise River Basin experienced wildfires of lesser extent or lower severity. We chose to focus on streams with relatively high degrees of potential influence to maximize detection of responses. Wildfires in stream catchments selected for the retrospective comparison occurred in 1992 and 1994 (Figure 1), with channel reorganizing events occurring immediately after wildfires in “burned and reorganized” streams followed by subsequent recurrences of varying magnitude 1–2 years post-fire.

Photographs of sites representative of streams sampled during the summer of 2003 for the retrospective comparative study (Figure 1). A Streams without recent stand-replacing wildfire in their watersheds (Trail Creek); B streams with moderate-to-high severity wildfire predominant in the watershed (Cottonwood Creek); C streams with moderate-to-high severity wildfire that resulted in massive channel reorganization (Trapper Creek).
We restricted our study frame to headwater streams draining catchments 1,000–5,300 ha in area, which generally correspond to wetted stream widths of less than 6 m. Catchments larger than this maximum size range are influenced by wildfire, but disturbances tend to be less severe (Benda and others 1998, 2003; Miller and others 2003; C. Luce, unpublished data), and large streams are less likely to experience associated changes in temperature from the loss of canopy near channels (Beschta and others 1987).
Within the range of catchment sizes and stream types (henceforth: burned, unburned, burned and reorganized), we defined stream networks and stream segments using a geographic information system (GIS, ArcGIS software; http://www.esri.com) and 30-m resolution digital elevation maps (DEMs) with the TauDEM software package (Tarboton 1997, 2004). Boundaries to stream segments were identified at tributary junctions, and characteristics of the segments were estimated from DEMs, including stream channel slope (%), contributing area (ha) and segment elevation (m) at the downstream end. These three variables are related to channel forms and landscape processes that influence stream temperatures, and thus provide a useful basis for matching streams to maximize the statistical validity of temperature comparisons among stream types (that is, minimize confounding in statistical comparisons). We used visual inspection of scatter plots based on raw data and principal components to identify streams and segments with overlapping elevation, channel slope, and contributing areas. Within each of the three stream types, we selected three streams for sampling summer temperatures. In each of the resulting nine streams, ten sample sites were randomly located within stream segments nested at progressively smaller catchments to a minimum of 400 ha. This design ensured that longitudinal gradients in streams were represented while maintaining randomness in the selection of specific site locations. We focused on stream segments draining catchments larger than 400 ha because they are more likely to support perennial surface flow and aquatic vertebrates. Summer stream temperatures were measured from 1 July to 15 September 2003.
Sampling of Fish and Amphibians
In the summer of 2003, we used backpack electrofishing with four removals to sample fish and amphibians in block-netted 100-m lengths of stream adjacent each site (n = 90) for the retrospective study of stream temperature. Native vertebrates in headwater streams within the study area include rainbow trout (Oncorhynchus mykiss), Rocky Mountain tailed frog (Ascaphus montanus), and bull trout (Salvelinus confluentus), with localized occurrences of shorthead sculpin (Cottus confusus). Nonnative species include cutthroat trout (O. clarkii), rainbow × cutthroat trout hybrids, and brook trout (S. fontinalis). Focal species in this effort included the two most common native species in the Boise River Basin: tailed frog and rainbow trout. Other species were recorded if found, but we considered their distributions to be too limited (for example, bull trout, nonnative brook trout, and cutthroat trout), or thermal tolerances too uncertain (for example, shorthead sculpin) to include them in this work. A validation study conducted in 2002 (Rosenberger and Dunham 2005; Rosenberger and Dunham, unpublished results) indicated this level of effort was nearly 100% effective for detecting both large (>150 mm) and smaller (<150 mm) salmonid fishes. Detectability using electrofishing is unknown for adult or larval tailed frog, the only common amphibian found in our study streams. All sampling for aquatic vertebrates was conducted during base flow conditions from mid-July to mid-September. Throughout the summer, we rotated our sampling schedule for streams in each category to avoid any potential confounding of stream type and time of sampling (Appendix 1, http://www.springerlink.com).
Data Analysis
Pre–Post Fire Comparisons
For the comparison of two sites (Cottonwood Creek and Roaring River), trends in differences in post-fire summer mean and maximum temperatures between the sites were assessed in relation to time since fire (years 1995–2005) using Spearman rank correlation. Confidence bounds for post-fire temperatures were compared to pre-fire temperature differences to determine the significance of post-fire temperature changes. For the short-term comparison of temperatures in sites sampled within James and Phifer Creeks, the significance of differences in maximum temperatures at sites in the years before and after a fire in James Creek in 2003 was assessed using t tests. Paired t tests were conducted to compare temperatures at sites within each stream (temperatures at sites sampled in 2002 vs. temperatures in the same sites in 2004). An overall t test was performed to compare overall average differences in temperatures before and after fire (all sites collectively) in each stream. We chose maximum temperature for this comparison because it is most likely to be responsive to changes in conditions associated with wildfire (Moore and others 2005a).
Retrospective Study
To consider the probable biological implications of temperature among streams, we modeled the probability of exceeding a maximum temperature of 20°C. The biological influences of temperature vary among species, but previous work indicates occurrence of tailed frog (larvae and adults) is very unlikely when maximum temperatures approach 20°C (for example, Pilliod and others 2003; Welsh and Lind 2002). Rainbow trout often occur at temperatures exceeding 20°C (for example, Zoellick 1999), but temperatures of this magnitude can have adverse sublethal influences (for example, reduced growth; Hokanson and others 1977).
To model the influence of stream type (unburned, burned, burned and channel reorganization) on biologically relevant changes in temperature, we used logistic regression to predict the probability of a site exceeding 20°C. This probability was modeled in relation to stream type and elevation, a covariate to account for longitudinal changes in stream temperature unrelated to stream type. To account for potential lack of independence in temperatures among sites sampled within streams, we used an autoregressive logistic model with an n − 1 spatial lag (Allison 1999). We examined standard measures of model fit, including deviance statistics and Hosmer–Lemeshow goodness of fit to assess the adequacy of a logistic model (Allison 1999).
Fish and Amphibian Distributions
Distributions of tailed frog larvae and small rainbow trout varied little among streams, so quantitative analysis of their associations with temperature was not possible or necessary. To examine the possible influence of different timeframes, we summarized temperatures in relation to the time of sampling. This was done to allow a more precise match between observed temperatures and vertebrate distributions. These summaries included daily mean and maximum daily temperatures prior to sampling (1 July to date of sample), week prior to sampling, and day of sampling (Appendix 1, http://www.springerlink.com).
Results
Pre–Post Fire Comparisons
Post-fire temperature data for comparing Cottonwood Creek and Roaring River were available for 1995–1997, 2001, and 2003–2005. Pre-fire summer mean and maximum temperatures were 1.1 and 1.7°C warmer in Cottonwood Creek, in comparison to Roaring River. Following the fire in Cottonwood Creek in 1994, this temperature difference increased; mean temperatures averaged 3.7°C warmer (3.3–4.0°C, 95% CI) across sampled years following the wildfire, and maximum temperatures averaged 5.2°C (3.2–7.2°C, 95% CI) warmer. Confidence intervals for means of both temperature differences did not overlap pre-fire temperature differences, indicating statistically significant post-fire increases across the timeframe (1995–2005) sampled. The difference in mean temperature between Cottonwood Creek and Roaring River following wildfire showed a declining trend over 1995–2005, but this trend was not statistically significant (r s = −0.66, P = 0.08). The corresponding difference in maximum temperature declined more strongly and significantly, however (r s = −0.91, P = 0.002). Maximum temperature in Cottonwood Creek peaked at an unusually high level in the summer of 1996 (Figure 3), but inspection of temperature records did not suggest anomalies, and this observation did not influence the direction or significance of the correlation between temperature differences and time since fire.

The difference between mean (unfilled symbols) and maximum (filled symbols) summer stream temperatures of two streams recorded across 13 years in the Boise River Basin (Figure 1) with differing burn histories. Timing of the wildfire in Cottonwood Creek is indicated by the vertical dotted line (1994; see also Figure 1). Temperature differences were calculated by subtracting temperatures recorded in Cottonwood Creek (burned) from those recorded in Roaring River (unburned).
In contrast to the comparison of pre- and post-fire temperatures at a single site over 13 years in Cottonwood Creek and Roaring River, pre- and post-fire temperature changes at multiple sites sampled in James and Phifer Creeks over 3 years were less obvious. Of the original ten sites sampled within each stream, data from two sites in each stream were lost due to failure to relocate dataloggers in 1 year (2002 or 2004). Analyses of the eight remaining sites suggested that overall average differences in maximum temperatures before and after fire were only marginally different between the two streams (t = 2.06, P = 0.06, df = 14). Within streams, paired tests revealed significant post-fire increases in temperature in both the burned James Creek (t = −4.46, P = 0.003, df = 7) and unburned Phifer Creek (t = −3.38, P = 0.02, df = 7). Within sites in James Creek, the average pre-post fire increase in temperature was 0.9°C (0.4, 1.4; 95% CI), whereas in Phifer Creek temperatures at sites increased an average of 0.4°C (0.1, 0.7; 95% CI).
Retrospective Study
Streams selected for the retrospective study (Figure 1) included three catchments with little history of recent stand-replacing fire (Beaver, Lost Man, and Trail Creeks). Within these catchments, stand-replacing fires were largely absent (Beaver and Lost Man Creeks) or limited in area compared to wildfire that burned with lower severity (Trail Creek, Figure 1). Three streams selected for study with recent stand-replacing fire occurring within a major portion of their catchments included Cottonwood, Hungarian, and Lost Creeks. The three selected streams with stand-replacing wildfire and recent channel reorganization (since 1992) included South Fork Sheep, Trapper, and Wren Creeks. Debris flows in Trapper and Wren Creeks occurred initially 1 year following wildfire (1995), with subsequent minor recurrences observed through 1996, whereas major channel reorganization in South Fork Sheep Creek occurred in 1993 and 1997 (J. Thornton, Boise National Forest, personal communication; C. Luce and B. Rieman, personal observations; Burton 2005). From interpretation of aerial photographs provided by the Boise National Forest, we estimated the percentage of channel reorganization (stream length) within South Fork Sheep, Trapper, and Wren Creeks to be 51, 85, and 97%, respectively. Within the constraints of catchment features (for example, area, elevation, and channel slope), spatial proximity, and common fire histories that were present, these nine catchments represented the best combination of streams we could choose for comparison in a retrospective study design.
For streams sampled as part of the retrospective study, summer maximum daily temperatures were strongly associated with stream type (unburned, burned, burned and reorganized; Appendix 1, http://www.springerlink.com). Maximum daily temperatures in sites from unburned streams averaged 15.6°C, and ranged from 11.7 to 20.4°C. Maximum daily temperatures in sites from burned streams averaged 19.0°C, and ranged from 13.8 to 24.6°C. Maximum daily temperatures in sites from burned and reorganized streams were warmest, averaging 23.5°C, and ranging from 19.2 to 26.6°C. Within each stream type, the greatest range in temperatures among sites sampled within streams was associated with the largest catchments sampled (for example, Lost Man, Cottonwood, and South Fork Sheep Creeks; Figure 1, Appendix 1). Overall, 41 of the 90 sites sampled exceeded 20°C on at least one occasion during the summer of 2003.
Logistic regression indicated the probability of stream temperature exceeding 20°C was associated with both stream type (X 2 = 28.9, P < 0.0001, df = 2) and site elevation (X 2 = 3.9, P = 0.05, df = 1). Classification rates using a leave-one-out cross validation indicated the logistic model correctly predicted 81.1% of the observations with a classification cutoff of 0.50 for events. A plot of predicted probability of exceeding 20°C against site elevation (Figure 4) indicated strong differences among stream types, with sites from unburned streams showing less than 0.25 probability of exceeding 20°C, sites from burned streams ranging from low (<0.25) to high (>0.75) probability of exceeding 20°C, and sites from burned and reorganized streams showing high (>0.75) probability of exceeding 20°C.

Plot of predicted probability of exceeding 20°C as a function of stream elevation for sites in nine streams in the Boise River Basin (Figure 1) with differing wildfire and channel disturbance history (closed circles unburned streams, open circles burned streams, gray triangles burned and reorganized streams).
Fish and Amphibian Distribution
Tailed frog larvae were detected in 85 of the 90 sites sampled. Tailed frog larvae were not detected from two sites in Beaver Creek (unburned), one site in Lost Creek (burned), and one site in Wren and South Fork Sheep Creeks (both burned and reorganized). Small rainbow trout occurred at every site sampled. Maximum daily stream temperatures at sites where tailed frog larvae and small rainbow trout were detected spanned the full range of observed summer temperatures (11.7–26.6°C). Examination of maximum stream temperatures prior to the dates of fish and amphibian sampling (Appendix 1) revealed no evidence for bias associated with matching temperatures that occurred after sampling. For example, Wren Creek was sampled after summer stream temperatures had peaked, with tailed frog larvae and small rainbow trout found in sites with maximum daily temperatures ranging from 22.5 to 26.6°C. This, combined with the fact that both small rainbow trout and tailed frog larvae were widespread within every stream sampled regardless of the time sampled, suggested they were present in sites when summer daily maximum temperatures occurred. Finally, a plot of the catch of tailed frog larvae in relation to maximum temperatures suggested that they were common (>10 individuals per site) much of the time within all sites, regardless of temperature within the observed range (Figure 5).

Plot of the total catch of tailed frog for each site sampled (n = 90) against maximum summer temperature of the site in streams in the Boise River Basin with differing burn histories (closed circles unburned streams, open circles burned streams, gray triangles burned and reorganized streams).
Discussion
Results of this work provide perspectives on the influences of wildfire and associated disturbances on stream temperature on both short (<5 years) and intermediate (5–11 years) timescales, including patterns from “before-after” comparisons and retrospective comparisons. In retrospective comparisons, stream temperatures were strongly related to past disturbance, especially in relation to channel reorganization following wildfire. Short- and long-term comparisons of pre- and post-fire stream temperatures did not show such dramatic differences, probably because they were not subject to channel reorganizing events. To our knowledge, there are no comparable studies of the effects of wildfire on stream temperatures at multiple sites involving these time scales. An understanding of the effects of wildfire and stream temperature at broader spatial and temporal scales is important for considering the population dynamics and local persistence of relatively mobile and longer-lived species that live in fire-prone areas, such as the stream-living vertebrates considered herein (Dunham and others 2003; Pilliod and others 2003; Burton 2005).
Within portions of the Boise River Basin, including streams sampled in the retrospective study and the pair of streams within the longer-term comparative study of stream temperatures (Figure 3), a consistent pattern was evident. Significantly elevated stream temperatures were apparent for at least a decade following stand-replacing wildfire. Presumably incident solar radiation was still increased over pre-fire levels in these streams (C. Luce, unpublished data), in spite of obvious re-growth of riparian vegetation, especially deciduous species re-sprouting from roots (Dwire and Kauffman 2003). In studies of the effects of forest harvest, low-stature riparian vegetation has been found to provide some degree of shade to streams (Moore and others 2005a), but did not appear to be sufficient to prevent elevated stream heating in our study system. Nevertheless, the fact that temperatures in burned streams in this study were not as warm as those in burned and reorganized streams (with little riparian vegetation) indicates that recovered streamside vegetation in burned streams does provide important shading. Longer-term increases in stream temperatures (>20 years) are more likely in systems where debris flows or severe floods completely eliminate streamside vegetation (including perennial shrubs and trees), which slows the recovery process (C. Luce, unpublished data).
Of all components of the stream heat budget, the influence of riparian shading is the most likely mechanism for the increases in stream temperatures that we observed. However, a host of other mechanisms could also play a role (Beschta and others 1987; Johnson and Jones 2000; Moore and others 2005a). Changes in stream morphology (for example, stream widening, loss of hyporheic exchange) following channel disturbance after wildfire could be factors that also account for the particularly high stream temperatures in these systems. In addition, pre- and post-fire differences in stream temperatures were less pronounced in the comparison of James (severely burned in 2003) and Phifer (unburned) Creeks, which lie in higher-elevation portions of the Boise River Basin outside of other streams studied herein. Loss of riparian vegetation following wildfire in James Creek undoubtedly lead to increased solar insolation, but other thermal processes affecting stream temperature may have been more important, accounting for the muted response to wildfire that we observed. Among these processes, groundwater flux and hyporheic exchange may be a moderating influence (Johnson and Jones 2000; Mellina and others 2002; Moore and others 2005b; Rothwell 2005).
Differences among streams and river catchments in the prevalence of different hydrologic influences on the heat budget implies that the magnitude and duration of stream warming that we observed in the Boise River Basin in burned streams may not be the case in all systems affected by wildfire. The importance of different components of the heat budget and how they are affected by wildfire in different river catchments merits further study. Conceivably, it is possible through an understanding of the relative importance of different thermal processes to identify streams that may be most vulnerable to increased temperatures as a result of wildfire and channel disturbance and associated influences on stream ecosystem processes. However, the effort involved to quantify different components of the heat budgets of streams (Webb and Zhang 1997) may limit the applicability of this concept at the broad scale needed for most management applications. Accordingly, a major challenge for research is to provide a better understanding of local processes that can be efficiently transferred to understanding thermal regimes at broad scales that are most relevant for species persistence and ecosystem function (Poole and others 2004).
Biological responses we observed for rainbow trout and tailed frog larvae in relation to stream temperatures only partially met our expectations. Rainbow trout can persist within the range of temperatures observed in this study, although temperatures on the higher end of what we observed can have important sublethal effects (Li and others 1994; Zoellick 1999; Gamperl and others 2002). Accordingly, the widespread presence of rainbow trout was not limited by temperature. Rapid (1-year) recolonization by rainbow trout following apparent complete extirpation from channel reorganization in streams studied here (Trapper and Wren Creeks) also supports the strong resilience of rainbow trout in this system (Rieman and others 1997; Burton 2005). Studies of the responses of salmonids to comparable disturbances in other areas have shown populations to be resilient as well (Lamberti and others 1991; Roghair and others 2002; Bisson and others 2005; Sestrich 2005), unless the habitats they occupy are highly fragmented (Brown and others 2001; Dunham and others 2003). In this study, only one stream was isolated by an impassible fish movement barrier (a perched road culvert at the mouth of Lost Creek), yet the population of rainbow trout upstream persisted in the face of wildfire. However, this stream did not experience massive channel reorganization like those causing apparent extirpations of populations nearby.
In contrast to our expectations, tailed frog larvae were also distributed widely across our study streams, and occurred in nearly every site sampled, regardless of temperatures ranging up to a maximum of 26.6°C. Most work on thermal relations of tailed frogs in the field has involved the coastal tailed frog (A. truei), which is generally restricted to streams with temperatures below 16°C (for example, Hawkins and others 1988; Pilliod and others 2003; Welsh and Lind 2002). The Rocky Mountain tailed frog, recently recognized as a distinct species, has evolved separately from the coastal tailed frog, likely representing isolation for more than ten million years (Nielsen and others 2001). Published observations for this species have recorded larvae in warmer temperatures (daily maximums up to 21°C) than would be typical for the coastal tailed frog (Adams and Frissell 2001). Adams and Frissell (2001) suggested tailed frog larvae might use coldwater seeps to avoid exposure to warm temperatures. In our study streams, we found little evidence of coldwater thermal refugia occurring at a site (100 m) scale, but undetected refugia could exist at much smaller scales. Alternatively, the short duration of exposure to peak daily temperatures (for example, <2 h) may not be sufficient to restrict the distribution of Rocky Mountain tailed frog larvae. Maximum temperatures observed in our study were less than the range of critical thermal maximums observed for coastal tailed frog larvae in the laboratory (28.9–30.1°C; Pilliod and others 2003). Finally, it is possible that greater availability of food in warmer habitats could extend the distribution of tailed frogs. Both temperature and light are greater in streams with reduced riparian shade. Availability of food for tailed frog larvae, which graze on periphyton, may be greater in streams that receive more light (Kiffney and others 2004), potentially compensating for the physiological costs of living in warmer environments.
In conclusion, results of this work suggest several important considerations for understanding the effects of wildfire and associated influences on stream temperatures and headwater vertebrates. First, it is clear that observable changes to stream temperatures can persist for at least a decade, and likely much longer following wildfire, especially in streams experiencing channel reorganization. Second, the specific magnitude and duration of temperature changes can be difficult to predict due to uncertainties regarding the spatial and temporal variability of influences from different mechanisms affecting the heat budget of streams. This was likely reflected in the variable responses of temperature to wildfire and channel reorganization both within and among streams we studied in the Boise River Basin. Third, in spite of the sometimes-dramatic physical changes to streams related to wildfire, the two vertebrate species considered herein were remarkably resilient in terms of their distribution across the landscape. Given that such disturbances have been a part of the history of streams over long periods (hundreds to thousands of years) in this region (Kirchner and others 2001; Pierce and others 2004) it is likely that these species have survived through similar, if not much larger events than studied here. Finally, in a management context, our results contribute needed empirical support for a growing consensus that management of human influences that modify species responses to wildfire (for example, habitat degradation, fragmentation, nonnative species) may be more effective than focusing on wildfire itself as a threat to fishes and amphibians in streams (Bisson and others 2003; Bury 2004; Dunham and others 2003; Minshall 2003; Pilliod and others 2003; Rieman and Clayton 1997; Rieman and others 2003).
References
Adams SB, Frissell CA. 2001. Thermal habitat use and evidence of seasonal migration by Rocky Mountain Tailed Frogs, Ascaphus montanus, in Montana. Can Field Nat 115:251–6
Allison PD. 1999. Logistic regression using the SAS system: theory and application. Cary, NC: SAS Institute
Benda L, Miller D, Dunne T, Agee J, Reeves G. 1998. Dynamic landscape systems. In: Naiman R, Bilby R, Eds. River ecology and management: lessons from the pacific coastal ecoregion. New York: Springer. Pp 261–88
Benda L, Miller D, Bigelow P, Andras K. 2003. Effects of post-wildfire erosion on channel environments, Boise River, Idaho. Forest Ecol Manage 178:105–49
Beschta, RL, Bilby RE, Brown GW, Holtby LB, Hofstra TD. 1987. Stream temperature and aquatic habitat: fisheries and forestry interactions. In: Salo EO, Cundy TW, Eds. Streamside management: forestry and fishery interactions. Contribution No. 57. Washington: Institute of Forest Resources, University of Washington, Seattle. pp 191–232
Bisson PA, Rieman BE, Luce C, Hessburg PF, Lee DC, Kershner JL, Reeves GH, Gresswell RE. 2003. Fire and aquatic ecosystems of the western USA: current knowledge and key questions. Forest Ecol Manage 178:213–29
Bisson PA, Crisafulli CM, Fransen BR, Lucas RE, Hawkins CP. 2005. Responses of fish to the 1980 eruption of Mount St. Helens. In: Dale VH, Swanson FR, Crisafulli CM, Eds. Ecological responses to the 1980 eruption of Mount St. Helens. New York: Springer. Pp 163–82
Brown DK, Echelle AA, Propst DL, Brooks JE, Fisher WL. 2001 Catastrophic wildfire and number of populations as factors influencing risk of extinction for Gila trout (Oncorhynchus gilae). West North Am Nat 61:139–48
Brown GW, Krieger JT. 1970. Effects of clearcutting on stream temperature. Water Resour Res 4:1133–9
Brown JK, Smith JK. 2000. Wildland fire and ecosystems: effects of fire on flora. General Technical Report RMRS-GTR-412-vol 2, USDA Forest Service, Ogden, UT, 257p
Burton TA. 2005. Fish and stream habitat risks from uncharacteristic wildfire: observations from 17 years of fire-related disturbances on the Boise National Forest, Idaho. Forest Ecol Manage 211:140–9
Bury RB. 2004. Wildfire, fuel reduction, and herpetofaunas across diverse landscape mosaics in northwestern forests. Conserv Biol 18:968–75
Dunham JB, Young MK, Gresswell RE, Rieman BE. 2003a. Effects of fire on fish populations: landscape perspectives on persistence of native fishes and nonnative fish invasions. Forest Ecol Manage 178:183–96
Dunham JB, Chandler G, Rieman BE, Martin D. 2005. Measuring stream temperature with digital data loggers: A user’s guide. General Technical Report RMRS-GTR-150-WWW. Fort Collins, CO: US Department of Agriculture, Forest Service, Rocky Mountain Research Station. 15p
Dunham JB, Vinyard GL. 1997. Incorporating stream level variability into analyses of fish-habitat relationships: some cautionary examples. Trans Am Fish Soc 126:323–9
Dwire KA, Kauffman JB. 2003. Fire and riparian ecosystems in landscapes of the western USA. Forest Ecol Manage 178:61–74
Gamperl KA, Rodnick KJ, Faust HA, Venn EC, Bennett MT, Crawshaw LI, Keeley ER, Powell MS, Li HW. 2002. Metabolism, swimming performance, and tissue biochemistry of high desert redband trout (Onchorynchus mykiss ssp.): evidence for phenotypic differences in physiological function. Physiol Biochem Zool 75:413–31
Gresswell RE. 1999. Fire and aquatic ecosystems in forested biomes of North America. Trans Am Fish Soc 128:193–221
Hawkins CP, Gottschalk LJ, Brown SS. 1988. Densities and habitat of tailed frog tadpoles in small streams near Mt. St. Helens following the 1980 eruption. J North Am Benthol Soc 7:246–52
Hitt NP. 2003. Immediate effects of wildfire on stream temperature. J Freshw Ecol 18:171–3
Hokanson KEF, Kleiner CF, Thorslund TW. 1977. Effects of constant temperatures and diel temperature fluctuations on specific growth and mortality rates and yield of juvenile rainbow trout, Salmo gairdneri. J Fish Res Board Can 34:639–48
Johnson SL. 2003. Stream temperature: scaling of observations and issues for modelling. Hydrol Process 17:497–9
Johnson SL, Jones JA. 2000. Stream temperature responses to forest harvest and debris flows in western Cascades, Oregon. Can J Fish Aquat Sciences 57:30–9
Jones JA, Post DA. 2004. Seasonal and successional streamflow response to forest cutting and regrowth in the northwest and eastern United States. Water Resour Res 40:W05203. doi:10.1029/2003WR002952
Kiffney PM, Richardson JS, Bull JP. 2004a. Establishing light as a causal mechanism structuring stream communities in response to experimental manipulation of riparian buffer width. J North Am Benthol Society 23:542–55
Kirchner JW, Finkel RC, Riebe CS, Granger DE, Clayton JL, King JG, Megahan WF. 2001. Mountain erosion over 10 yr, 10 k.y., and 10 m.y. time scales. Geology 29:591–4
Lamberti GA, Gregory SV, Ashkenas LR, Wildman RC, Moore KMA. 1991. Stream ecosystem recovery following a catastrophic debris flow. Can J Fish Aquat Sci 48:196–208
Li HW, Lamberti GA, Pearsons TN, Tait CK, Li JL, Buckhouse JC. 1994. Cumulative effects of riparian disturbances along high desert trout streams of the John Day Basin, Oregon. Trans Am Fish Soc 123:627–40
McMahon TE, deCalesta DS. 1990. Effects of fire on fish and wildlife. In: Walstad J, Ed. Natural and prescribed fire in Pacific Northwest forests. Corvallis, OR: Oregon State University Press. Pp 233–50
Mellina E, Moore RD, Hinch SG, Macdonald JS, Pearson G. 2002. Stream temperature responses to clearcut logging in British Columbia: the moderating influences of groundwater and headwater lakes. Can J Fish Aquat Sci 59:1886–900
Meyer GA, Pierce JL. 2003. Climatic controls on fire-induced sediment pulses in Yellowstone National Park and central Idaho: a long-term perspective. Forest Ecol Manage 178:89–104
Miller D, Luce C, Benda L. 2003 Time, space, and episodicity of physical disturbance in streams. Forest Ecol Manage 178:121–40
Minshall GW. 2003. Responses of stream benthic macroinvertebrates to fire. Forest Ecol Manage 178:155–62
Minshall GW, Robinson CT, Lawrence DE. 1997. Postfire responses of lotic ecosystems in Yellowstone National Park, U.S.A. Can J Fish Aquat Sci 54:2509–25
Moore RD, Spittlehouse D, Story A. 2005a. Riparian microclimate and stream temperature response to forest harvesting: a review. J Am Water Resour Assoc 41:813–34
Moore RD, Sutherland P, Gomi T, Dhakal A. 2005b. Thermal regime of a headwater stream within a clearcut, coastal British Columbia, Canada. Hydrol Process 19:2591–608
Nielson M, Lohman K, Sullivan J. 2001. Phylogeography of the tailed frog (Ascaphus truei): implications for the biogeography of the pacific northwest. Evolution 55:147–60
Pierce JL, Meyer GA, Jull AJT. 2004. Fire-induced erosion and millennial-scale climate change in northern ponderosa pine forests. Nature 432:87–90
Pilliod DS, Bury RB, Hyde EJ, Pearl CA, Corn PS. 2003. Fire and amphibians in North America. Forest Ecol Manage 178:163–82
Poole GC, Dunham JB, Keenan DM, Sauter ST, McCullough DA, Mebane C, Lockwood JC, Essig DA, Hicks MP, Sturdevant DJ, Materna EJ, Spalding SA, Risley J, Deppman M. 2004. The case for regime-based water quality standards. Bioscience 54:154–61
Rieman BE, Clayton J. 1997. Wildfire and native fish: issues of forest health and conservation of native fishes. Fisheries 22:6–15
Rieman BE, Lee DC, Chandler G, Myers D. 1997. Does wildfire threaten extinction for salmonids: responses of redband trout and bull trout following recent large fires on the Boise National Forest. In: Greenlee J, Ed. Proceedings of the Conference on Wildfire and Threatened and Endangered Species and Habitats, 13–15 November 1995, Coeur d’Alene Idaho. Washington: International Association of Wildland Fire, Fairfield. Pp 47–57
Roghair CN, Dolloff CA, Underwood MK. 2002. Response of a brook trout population and instream habitat to a catastrophic flood and debris flow. Trans Am Fish Soc 131:718–30
Rosenberger AE, Dunham JB. 2005. Validation of abundance estimates from mark–recapture and removal techniques for rainbow trout captured by electrofishing in small streams. North Am J Fish Manage 25:1395–410
Rothwell EL. 2005. The influence of hyporheic flow on the temperature of a riffle-step-pool stream. Thesis (M.S.), Boise State University, 2005. 88p
Sestrich CM. 2005. Changes in native and nonnative fish assemblages and habitat following wildfire in the Bitterroot River Basin, Montana. Thesis (M.S.), Montana State University. 93p
Spencer CN, Gabel KO, Hauer FR. 2003. Wildfire effects on stream food webs and nutrient dynamics in Glacier National Park, USA. Forest Ecol Manage 178:141–53
Swanson FJ. 1981. Fire and geomorphic processes. Mooney HA, Bonnicksen TM, Christensen NL, Lotan JE, Reiners WA, Eds. Fire regimes and ecosystem properties: USDA Forest Service General Technical Report WO-26, Washington DC
Tarboton DG. 1997. A new method for the determination of flow directions and contributing areas in grid digital elevation models. Water Resour Res 33:309–19
Tarboton DG. 2004. Terrain analysis using digital elevation models: a graphical user interface software package for the analysis of digital elevation data and mapping of channel networks and watersheds. Logan, UT: Utah Sate University
Webb BW, Zhang Y. 1997. Spatial and seasonal variability in the components of the river heat budget. Hydrol Process 11:79–101
Welsh HH, Lind AJ. 2002. Multiscale habitat relationships of stream amphibians in the Klamath-Siskiyou region of California and Oregon. J Wildlife Manage 66:581–602
Wondzell SM, King JG. 2003. Postfire erosional processes in the Pacific Northwest and Rocky Mountain regions. Forest Ecol Manage 178:75–88
Ziemer RR. 1964. Summer evapotranspiration trends as related to time after logging of forests in Sierra Nevada. J Geophys Res 69:615–20
Zoellick BW. 1999. Stream temperatures and the elevational distribution of redband trout in southwestern Idaho. Great Basin Nat 59:136–43
ACKNOWLEDGEMENTS
Funding for this work was provided by the National Fire Plan, the US Forest Service Rocky Mountain Research Station, the US Forest Service Regions 1 and 4, US Geological Survey, Forest and Rangeland Ecosystem Science Center, and local assistance from the Boise National Forest. We thank, among many, Michael Kellett, Dona Horan, Gwynne Chandler, Joseph Benjamin, Tracie Kirkham, Matt Aylsworth, Patrick Mckann, Brooke Nelson, Dan Bolin, Ciara Cusack, Tobias Jongeward, Tim Arrington, Kristin Bott, Aaron Golart, Kristen Bednarczyk, Mary Beatty, and Garet Tessmer for assistance in the field. Thanks to David Nagel, Sharon Parkes, and Joe Benjamin for support with spatial data analysis and preparation. Thanks to D. Pilliod and B. Bury for sharing their insights on tailed frogs.
Author information
Authors and Affiliations
Corresponding author
Electronic Supplementary Material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Dunham, J.B., Rosenberger, A.E., Luce, C.H. et al. Influences of Wildfire and Channel Reorganization on Spatial and Temporal Variation in Stream Temperature and the Distribution of Fish and Amphibians. Ecosystems 10, 335–346 (2007). https://doi.org/10.1007/s10021-007-9029-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10021-007-9029-8