
Abstract
Studies of long-term soil and ecosystem development on static geomorphic surfaces show that old soils become depleted in most rock-derived nutrients. As they are depleted, however, static surfaces also are dissected by fluvial erosion. This fluvial erosion leads to colluvial soil transport on the resulting slopes, which in turn can rejuvenate the supply of weathering-derived nutrients to plants. We evaluated the influence of erosion and consequent landscape evolution on nutrient availability along a slope on the Island of Kaua’i, near the oldest, most nutrient-depleted site on a substrate age gradient across the Hawaiian Islands. Noncrystalline minerals characteristic of younger Hawaiian soils increased from 3% of the soil on the static constructional surface at the top of the slope to 13% on the lower slope, and the fraction of soil phosphorus (P) that was occluded (and hence unavailable) decreased from 80% to 56% at midslope. Foliar nitrogen and P concentrations in Metrosideros polymorpha increased from 0.82% and 0.062% to 1.13% and 0.083% on the constructional surface and lower slope, respectively. The increase in foliar P over a horizontal difference of less than 250 m represents nearly half of the total variation in foliar P observed over 4.1 million years of soil and ecosystem development in Hawai’i. The fraction of foliar strontium (Sr) derived from weathering of Hawaiian basalt was determined using 87Sr:86Sr; it increased from less than 6% on the constructional surface to 13% and 31% on lower slope and alluvial positions. Erosional processes increase both nutrient supply on this slope and the fine-scale biogeochemical diversity of this old tropical landscape; it could contribute to the relatively high level of species diversity observed on Kaua’i.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
As soils in humid environments age, their stock of minerals susceptible to weathering becomes depleted. Plant productivity on old soils is supported proximately by efficient recycling of elements within ecosystems, and ultimately by inputs of elements from the atmosphere. Unless those inputs are substantial, soil fertility declines, and phosphorus (P) in particular is likely to become limiting to plant growth (Walker and Syers 1976). This pattern of development underlies the widespread occurrence of nutrient-depleted, infertile soils on old substrates from the tropics to the warm temperate zone (Sanchez 1976; Richter and Markewitz 2001).
These dynamics of long-term soil development and their implications for ecosystem functioning have been documented along a 4-million-year gradient in substrate age across the Hawaiian Islands (Riley and Vitousek 1995; Crews and others 1995; Vitousek and others 1997; Chadwick and others 1999). The islands represent a linear array of volcanoes that increase in age from southeast to northwest, and the current climate, parent material, topography, and vegetation of sites across the islands can be held nearly constant. Moreover, the origin of the islands in Earth’s mantle means that Hawaiian basalt differs systematically from the surrounding ocean and from upwind continents in its strontium isotope ratios (Capo and others 1998; Kennedy and others 1998; Stewart and others 2001), and in other ways that can be used as mineralogical, elemental, and isotopic tracers of the sources of elements to soils and ecosystems (Chadwick and others 1999; Kurtz and others 2000, 2001).
Across the Hawaiian age gradient, most primary minerals have been depleted and most rock-derived cations lost via leaching by about 20,000 years (Vitousek and others 1997; Chadwick and others 1999). Thereafter, precipitation is the major source of strontium (Sr) and presumably of other divalent cations as well (Kennedy and others 1998). P is less mobile than cations, and rock-derived P remains important for over a million years. However, most rock-derived P is lost by 4 million years; P supply limits plant productivity (Herbert and Fownes 1995), and the majority of P inputs to the oldest site are derived via long-distance transport of dust from Asia (Chadwick and others 1999).
These progressive changes in soil fertility and ecosystem functioning apply only to the geomorphologically static, constructional topography on which they were documented. Another progressive change across the Hawaiian Islands is that, as landscapes age, fluvial erosion dissects the constructional surfaces, creating a diverse landscape of ridges, valleys, and hillslopes. The landscape progresses from being composed almost entirely of constructional surfaces; to a matrix of constructional, eroded, and depositional surfaces in which the proportion of constructional surfaces diminishes over time; to landscapes in which virtually the entire surface is either eroded or depositional (Figure 1). In the Hawaiian Islands, less than 2% of the surface of 4-million-year-old Kaua’i is made up of constructional surfaces (McDonald and others 1983).
In this article, we evaluate whether the colluvial transport of soils that dominate slopes can rejuvenate the supply of rock-derived nutrients on old substrates, by removing nutrient-depleted material and giving biological communities access to rock-derived elements from deep in the soil profile. The fact that erosion can “reset the clock” of soil and ecosystem development has long been recognized (Lepsch and others 1977; Jenny 1980). Numerous studies of landslides [for example, see Zarin and Johnson (1995), Walker and others (1996), and Restrepo and others (2003)] demonstrate that sufficiently large and deep slides push ecosystems to very early stages of soil development and primary succession, and studies of topographic gradients (catenas or toposequences) yield predictable changes in soil depth, development, and nutrient availability as a function of slope position [for example, see Schimel and others (1985), Brubaker and others (1993), and Enoki and others (1997)]. On older soils, landslides and other pathways of mass loss can play dual roles as short-term vectors of nutrient loss and longer-term enhancers of nutrient inputs via weathering (Scatena and Lugo 1995), and a number of studies in the tropics and subtropics have reported greater availability of rock-derived nutrients on slopes and depositional areas in comparison to stable geomorphic surfaces (Tanner 1977; Silver and others 1994; Scatena and Lugo 1995; Chen and others 1997; Restrepo and others 2003).
The Hawaiian Islands provide an unusually valuable context and set of tools for understanding the consequences of erosion and landscape evolution for biogeochemistry, including: (a) the same dominant tree species in all slope positions, (b) a consistent difference in Sr isotopes between Hawaiian basalt and atmospheric inputs that enables the source of Sr in ecosystems to be identified, and (c) well-documented patterns of nutrient supply and forest response during ecosystem development on constructional surfaces. We determined soil properties, nutrient availability, and plant tissue chemistry across a topographic gradient developed in 4.1-million-year-old substrate on the Island of Kaua’i, near the oldest site on a substrate age gradient across the Islands. Soils on the constructional surface there are depleted in rock-derived nutrients, most soil P is in occluded forms, and forest growth is limited by the supply of P (Crews and others 1995; Herbert and Fownes 1995; Chadwick and others 1999).
Methods
Study Sites
Our focal topographic gradient was located at 22°9′N, 159°37′W, near the Alakai Swamp Trail in the Na Pali-Kona Forest Reserve (Figure 2). The topography of the area consists of flat-topped ridges that lead west and southwest from the east Kaua’i volcano (Holcomb and others 1997); these represent the remaining constructional surfaces on the Island of Kaua’i. These remnant surfaces are dissected by steep-sided stream valleys up to 100 m deep, and the region as a whole is bounded by much deeper cliffs and valleys (Figure 2). Rainfall at the focal site is approximately 3000 mm/year (Giambelluca and others 1986) and declines substantially from east to west across the area.
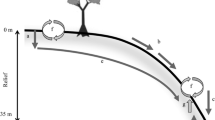
We chose an accessible slope that supports closed-canopy forest; many slopes nearby are steeper and fern dominated. We surveyed the slope by using a Total station, beginning on the constructional surface at an elevation of 1125 m. Our main transect stretched 400 m horizontally and incorporated 85 m of relief (Figure 3). We divided this slope into the static shield surface, a gentle shoulder slope, a steep back slope (averaging > 20°), a flatter toe slope, and a small streamside alluvial area. In addition to this focal slope, we collected foliage of Metrosideros polymorpha from three constructional surface–slope pairs arrayed along the regional rainfall gradient (Figure 2). These include Kilohana (∼5000 mm/year precipitation), on the constructional surface and the steep slope of Wainiha Valley; the long age-gradient site (2500 mm/year) and a steep adjacent slope; and Kumuwela (1700 mm/year), on an Acacia koa age-gradient site (Pearson and Vitousek 2002) and the adjacent slope. Metrosideros dominates the three wetter sites; Acacia dominates the driest one, with a few large Metrosideros in the overstory.
Sampling and Analyses
Soils.
Soil profiles were described and sampled in March 2000 for determination of mineralogy and soil P fractions. Ten soil pits were arrayed along the focal transect—two on the shield surface, one shoulder slope, one upper slope, two midslope, two lower slope, and two toe slope. Each pit was dug to greater than 110-cm depth or to a horizon that was difficult to penetrate; in the shoulder, upper, and midslope positions, we encountered relatively unweathered rock at less than 100 cm (Table 1). Soils on the shield surface developed in relatively deep residual material, whereas slope soils developed in colluvial material that increased in depth from the upper to the lower slope (Table 1). Based on landscape position, soil morphology, and our observation of thick layers of continentally derived dust near the surface, the toe slope represents the surface of a large, ancient translational slump that has been stable for at least tens of thousands of years.
Soil profiles were identified and described in the field, and sampled via channel sampling using standard USDA–Natural Resource Conservation Service (NRCS) procedures (Schoeneberger and others 1998). We noted the deepest soil horizon containing common roots (Table 1); generally, most roots were found in the top 10–30 cm of soil, but they reached down to relatively unweathered rock in the midslope (Table 1). Bulk density was measured using −33-kPa equilibration of saran-coated natural clods (Soil Survey Laboratory Staff 1992). Subsamples of the less than 2-mm fraction were treated to remove organic matter using hydrogen peroxide and dodium acetat. buffered at pH 5, and the noncrystalline mineral portion was measured by weight loss after extraction using acid ammonium oxalate in the dark [modified from Jackson and others (1986) and McKeague and Day (1966)]. The distribution of P fractions in the less than 2-mm soil fraction was characterized using a modified Hedley–Stewart fractionation (Hedley and others 1982; Tiessen and Mohr 1993).
In addition, five samples from surface mineral horizons (to 10 cm) were collected at six points along the transect (shield surface, shoulder slope, midslope, lower and toe slopes, and alluvium) in June 2000 for determination of nitrogen (N) mineralization and nitrification. Rocks and roots were removed from each sample, which was then divided into three subsamples. One subsample was weighed, dried at 105°C, and reweighed to determine water content; the second was extracted in 2 M potassium chloride (KCl), and the third was incubated for 30 days in the lab at 22°C, and then extracted in KCl. Finally, 10 anion and cation exchange resin bags (Binkley and Matson 1983) were placed 10 cm deep in the soil at each of the six slope positions in June 2000 and retrieved 30 days later. Resin bags were then extracted in 0.5 M HCl and analyzed for ammonium, nitrate, and phosphate.
Ammonium and nitrate concentrations in soil extracts were determined on an Alpkem AutoAnalyzer at Stanford University; net N mineralization was calculated as final (postincubation) ammonium-N plus nitrate-N minus initial ammonium-N plus nitrate-N, whereas net nitrification was calculated as final minus initial nitrate-N. Ammonium, nitrate, and phosphate in the resin extracts also were determined using the Alpkem AutoAnalyzer, following neutralization of the HCl; results are reported as microgram sof ammonium-N, nitrate-N, or phosphate-P absorbed per bag per day.
Foliage.
Foliar nutrient concentrations provide an index of nutrient availability to plants (Van den Driessche 1974), one that correlates well with soil measurements and with nutrient limitation to plant growth in Hawaiian Metrosideros forests (Vitousek and others 1995, 1997). Leaves of Metrosideros polymorpha were collected across the main transect four times in 1999–2001. Canopy twigs were shot down with a slingshot, and the youngest fully expanded sun leaves (behind a formed bud or developing flush) were harvested. Leaves were kept at 4°C until leaf area was determined on a Delta-T leaf area meter, generally within 48 h. In all, five trees were sampled on the constructional surface, two on the shoulder slope, five upper slope, eight midslope, four lower slope, two toe slope, and two alluvial. In addition, leaves from two or more Metrosideros trees were collected on the shield and on steep slopes at the other sites along the moisture gradient.
Leaves were oven dried at 70°C and ground. A subsample from each tree was acid digested using a block digestor, and analyzed for N and P on the AutoAnalyzer. A second subsample was ashed in a muffle furnace at 550°C; the residue was then dissolved in nitric acid (HNO3), and analyzed for cation concentrations using atomic absorption spectrophotometry. A third subsample was analyzed for Sr isotope ratios at Cornell University or Stanford University, using one of two initial procedures. At Cornell, samples were ashed at 450°C, and the residue was digested in ultrapure chloric acid, HClO4 and HNO3; Sr was separated using Eichrom Sr-specific resin. At Stanford, samples were ashed at 550°C and dissolved in 2.5 M HCl, and Sr was separated using AG-50 resin. Metrosideros tissues analyzed in both labs yielded identical results (within analytical error); repeated analyses of the standard NBS-987 averaged 0.710246 at Cornell and 0.71024 at Stanford, with 2σ variations of 0.00001 and 0.00005, respectively.
Statistics.
One-way analysis of variance was performed on log-transformed data to test for significant variation in foliar and soil characteristics among slope positions, using Systat8 (SPSS 1988). Where significant (P < 0.05) F values were obtained, slope positions were compared pairwise using Tukey’s honestly significant difference.
Results
The soils of the shield surface and the toe slope classified as Humaquepts, although their properties are close to those of the Plinthic Kandiudox found in comparable slope positions at the slightly drier age-gradient site. In contrast, the upper-, mid-, and lower-slope soils are Hapludands and Hydrudands (Table 1). The abundance of noncrystalline soil minerals (allophane, imogolite, and ferrihydrite) varied from a depth- and density-weighted mean of approximately 3% on the constructional surface to 8% and 13% on the midslope and lower slope, respectively, before dropping to 3% on the toe slope (Figure 4). These noncrystalline minerals dominate the Andisols of younger sites on the age gradient across the Hawaiian Islands (Torn and others 1997; Vitousek and others 1997), and their increased abundance on slopes here demonstrates that slope soils are functionally younger than those on the shield surface.
Soil P fractions also varied along the slope transect. Nearly 80% of the P on the shield was in the residual form—a fraction that roughly corresponds to the occluded (insoluble and/or physically protected) form in Walker and Syers’ (1976) scheme—whereas residual P on the slope ranged from 45% to 68% (Table 2). In contrast, organic P increased from 6% on the shield to 20%–40% on the slopes. HCl-extractable P (corresponding to P in primary minerals) was less than 1.5% in all slope positions, and even that may reflect primary minerals in Asian dust (Kurtz and others 2001).
Available P as indexed by field-incubated ion exchange resins was significantly greater in the alluvial soil than in all other slope positions. Although the midslope and lower slope had about twice as much resin-available P as the shield surface and toe slope (Figure 5C), this difference was not statistically significant. Resin-extractable NO3-N was significantly elevated in the upper slope, midslope and lower slope positions, in comparison to shield, shoulder, toe, or alluvial sites (Figure 5B). Resin extractable NH4-N followed no clear pattern (Figure 5A) across the slope, although lower-slope concentrations were significantly greater than those on the toe slope. Both N mineralization and nitrification were significantly greater on the slope and (marginally) alluvium than in the relatively static shield and toe-slope landscape positions (Figure 6).
P concentrations in Metrosideros leaves increased monotonically from 0.062% on the shield surface to 0.083% on the lower slope (Figure 7B), before dropping to 0.064% on the toe slope. N concentrations followed a similar pattern (Figure 7A). Foliar N on the toe slope in particular was strikingly low, significantly below foliar N concentrations on the midslope and lower slope.
Calcium (Ca) and magnesium concentrations in Metrosideros foliage decreased from shield to slope; Ca then increased on toe slope and alluvium. The low foliar Ca on slopes is comparable to that in intermediate-aged sites on the substrate age gradient across the islands (Vitousek and others 1995). In contrast, K concentrations were lowest on shield and toe slope and higher elsewhere (Table 3).
Foliar Sr on the shield was derived almost entirely from atmospheric deposition; its 87Sr:86Sr ratio was 0.7089, versus 0.7092 for marine aerosol. Less than 6% of foliar Sr was derived from basalt weathering, which has a 87Sr:86Sr ratio of 0.7036 (Kennedy and others 1998). The fraction of Sr derived from basalt increased to 12%–14% on the slope, and up to more than 30% in the alluvial area (Figure 8); the increase from shield to alluvial was statistically significant. These results suggest that the supply of rock-derived elements is rejuvenated on slopes, although not to the point where it dominates cation supply.
Measurements of foliar N, P, and Sr isotopes in three additional shield–slope pairs arrayed across the regional rainfall gradient (Figure 2) yielded similar results. Foliar P in Metrosideros was greater in all four slope sites (including the focal transect) in comparison to adjacent shield surfaces, across a precipitation range from 1700 to 5000 mm/year (Table 4). The difference between shield and slope positions decreased with decreasing precipitation. Foliar N generally followed the same pattern. The 87Sr:86Sr ratio showed a stronger contribution of basalt weathering on all four slopes in comparison to the adjacent shield sites (Table 4). 87Sr:86Sr ratios on two of the shield surfaces were above those in marine aerosol, suggesting that long-distance transport of Asian dust contributed more Sr in those sites than did basalt weathering (Kennedy and others 1998; Kurtz and others 2001).
Discussion
The biological availability of N and P increased substantially on slopes in comparison to constructional landscape positions, as documented by both soil and plant properties (Figures 5, 6, 7 and Table 4). The overall pattern of increased nutrient availability downslope is familiar from many studies on toposequences (Schimel and others 1985; Silver and others 1994; Chen and others 1997; Enoki and others 1997; Hirobe and others 1998). In Puerto Rico, Scatena and Lugo (1995) interpreted this increase as reflecting nutrient depletion on more stable ridge positions, coupled with movement of water and of nutrient-rich particles to downslope positions. They demonstrated that both geologic and ecological disturbances were more frequent on slopes; landslides did not occur on ridge tops, but had an approximate 1000-year return interval on slopes. Moreover, both tree-fall gaps and background mortality led to faster turnover of forest biomass on slopes than on ridges. Consequently, the slope and riparian areas were systematically younger in both soil and forest age than the more stable ridge areas (Scatena and Lugo 1995).
On the main Kauai slope discussed here, several results suggest that the slope soils are effectively younger than those on the shield surface. Soils on the geomorphologically static shield surface are dominated by crystalline clay minerals and enriched in the immobile elements zirconium, niobium, and tantalum, demonstrating intense weathering and the loss of most of the soil mass by leaching (Vitousek and others 1997; Kurtz and others 2000). Noncrystalline minerals characteristic of earlier stages of soil development are more abundant in slope than shield soils (Figure 4), and occluded P decreases while organic P increases downslope in a pattern consistent with that observed in younger, more fertile soils (Crews and others 1995). Moreover, the fraction of plant Sr derived from basalt weathering versus marine aerosol increases downslope (Figure 8). Although this increase is relatively small, Sr (like other alkaline earth elements) is highly mobile in soils—and the weathering contribution of less mobile elements like P on slopes likely is greater than that of the major cations.
The foliar chemistry of Metrosideros polymorpha can be compared directly between this and other gradients in Hawai’i, as the same species and canopy position were sampled in every case. On our main topographic gradient, foliar N and P increased from 0.82% and 0.062% on the shield surface to 1.13% and 0.083% on the lower slope. In contrast, foliar N peaked at 1.42% in the 20,000-year-old site on the substrate age gradient across Hawai’i, and P peaked at 0.113% in the 150,000-year-old site (Vitousek and others 1995). Variation in foliar chemistry along a horizontal distance of less than 250 m was 54% and 45% of that over 4 million years of soil and ecosystem development, for N and P, respectively.
The transport of nutrients in solution could contribute to increased nutrient availability on slopes (Giblin and others 1991; Silver and others 1999), and the substantial increase in resin-extractable nitrate on midslope and lower slope (Figure 5B) appears consistent with transport. However, rates of N mineralization and nitrification also are substantially greater on slope than shield surface (Figure 6), suggesting that much or all of the nitrate on the slope represents in situ production in a relatively N-rich site—again like younger, more fertile soils on the age gradient.
The variation in nutrient availability and plant chemistry along topographic gradients that we observed here has several implications. First, blanket statements that “soils on old substrates are highly weathered and infertile” may apply to static surfaces, but they are misleading when applied to entire landscapes (Figure 1). Only when the parent material itself is highly weathered from previous erosional cycles, or when the depth of weathering is greater than that of erosion, will entire landscapes support more or less infertile soils. Second, not all soils affected by erosion are relatively fertile—the source areas of landslides generally are infertile because they represent a very early, nutrient-poor stage of soil development (Walker and others 1996). Also, long-stabilized features like the toe slope we sampled on the Kauai slope (Figure 3) can be very low in available nutrients—and could have been so since they formed.
Third, we suggest that the interaction between soil development and the evolution of landscapes—between nutrient depletion on stable surfaces and erosion that dissects those surfaces and creates complex landscapes—drives systematic changes in the biogeochemical diversity of landscapes. Very young landscapes in Hawai’i are comprised of a fine-scale matrix of lava flows of different ages and different nutrient availability (Vitousek and others 1992; Vitousek and Benning 1995), although at this young age all of the substrates are relatively infertile. Older areas (tens to hundreds of thousands of years) generally are more uniform; accumulated volcanic ash deposition has blurred the boundaries between lava flows, soil development has proceeded to relatively fertile soils, and the surface is as yet little dissected. In still older areas like Kaua’i, erosion has caused the landscape to become a fine-scale matrix once again, now with nutrient-depleted remnant shield surfaces, rejuvenated slopes, alluvial areas, and other landforms in close proximity (Figure 1). This fine-scale variation in nutrient availability overlays coarser-scale variation associated with climatic gradients at all stages of ecosystem development (Harrington and others 1995; Austin and Vitousek 1998; Chadwick and Chorover 2001; Schuur and Matson 2001).
Finally, the biogeochemical diversity of old landscapes could contribute to patterns of species diversity. In Hawai’i, plant diversity increases with island age, reaching a peak on Oahu and Kaua’i, the oldest islands that still support a wide range of climate and topography. This systematic increase in diversity is widely believed to be due to a long-term accumulation of the products of evolution and speciation—a reasonable explanation. However, the greater biogeochemical diversity of these old landscapes also could contribute to the formation and maintenance of diversity on the older islands, by providing a broader range of conditions in which species could evolve and persist.

Changes in topography as a function of volcano age. Fluvial erosion progressively dissects the constructional surfaces of shield volcanoes, to the point where few stable surfaces remain on the oldest islands. (a) the eastern portion of the active Kilauea volcano. Most of the surface is <1000 yrs old; the visible topography is derived from volcanic (cinder cones and craters) and tectonic activity (block faults on the south flank). (b) Kohala Volcano, which developed ~ 500,000 years bp with substantial post-shield erruptive activity 150,000–200,000 years bp. Both volcanic and fluvial erosional features are visible; fluvial erosion is more important on the wetter eastern portion of the volcano. (c) West Maui Volcano formed ~1.2–1.4 million years bp. Most of the surface has been influenced by fluvial erosion, although the outline of the original shield volcano still is apparent. (d) The Island of Kaua’i, which formed 4–5 million years bp. Almost the entire surface has been influenced by fluvial erosion, including canyons > 1000 m deep. All of the volcanoes are mapped to scale; all reach a maximum elevation of 1200–1600 m.

Locations of the main slope transect (triangle) and the satelite constructional surface-slope pairs (circles) on the island of Kaua’i, Hawai’i. The area of gentle slopes south and east of the sites is the Alakai Swamp, which includes the last remaining constructional surfaces on Kaua’i. The sites are arrayed on a moisture gradient, with Kilohana to the northeast receiving ~5000 mm/yr and Kumuwela to the southwest ~ 1700 mm/yr (Giambelluca and others 1986).

A profile diagram of slope positions and elevations along the main transect, on the Alakai Swamp Trail, Kaua’i. The vertical scale is exaggerated four-fold.

The depth-integrated abundance of non-crystalline minerals in soils along the slope transect, Kaua’i.

Ammonium-n (A), nitrate-N (B), and phosphate-P (C) absorbed by resin bags placed along the topographic gradient, in μg per bag per day. Values are means (N = 10), with standard errors; bars that do not share a common letter differ significantly at p < .05.

Net N mineralization (A) and net nitrification (B) measured in laboratory incubations of A-horizon soils collected along the topographic gradient on Kaua’i. Values are means (N = 5), with standard errors; bars that do not share a common letter differ significantly.

(A) Foliar N (%) in sun leaves of Metrosideros polymorpha along the topographic gradient on Kaua’i. (B) Foliar P (%) in Metrosideros on the topographic gradient. Values are means, with standard errors; bars that do not share a coomon letter differ significantly.

Bars represent the ratio of 87Sr: 86Sr in Metrosideros leaves collected along the topographic gradient on Kaua’i. Values are means, with standard errors; bars that do not share a common letter differ significantly. The line shows the percentage of foliar Sr derived from basalt weathering, assuming a simple two-component mixing model of basalt ( 87Sr/86Sr = 0.7036) and atmospheric deposition of marine aerosol ( 87Sr/86Sr = 0.7092).
References
AT Austin PM Vitousek (1998) ArticleTitleNutrient dynamics on a precipitation gradient in Hawai’i Oecologia (Berl) 113 519–29
D Binkley PA Matson (1983) ArticleTitleIon exchange resin bag method for assessing forest soil nitrogen availability Soil Sci Soc Am J 47 1050–2
SC Brubaker AJ Jones DT Lewis K Frank (1993) ArticleTitleSoil properties associated with landscape position Soil Sci Soc Am J 57 235–9
RC Capo BW Stewart OA Chadwick (1998) ArticleTitleStrontium isotopes as tracers of earth surface processes: theory and methods Geoderma 82 197–225
OA Chadwick J Chorover (2001) ArticleTitleThe chemistry of pedogenic thresholds Geoderma 100 321–53
OA Chadwick L Derry PM Vitousek BJ Huebert LO Hedin (1999) ArticleTitleChanging sources of nutrients during four million years of ecosystem development Nature 397 491–7
Z-S Chen C-F Hsieh F-Y Jiang T-S Hsieh I-F Sun (1997) ArticleTitleRelations of soil properties to topography and vegetation in a subtropical rain forest in southern Taiwan Plant Ecol 132 229–41
TE Crews K Kitayama J Fownes D Herbert D Mueller-Dombois RH Riley PM Vitousek (1995) ArticleTitleChanges in soil phosphorus and ecosystem dynamics across a long soil chronosequence in Hawai’i Ecology 76 1407–24
T Enoki H Kawaguchi G Iwatsubo (1997) ArticleTitleNutrient-uptake and nutrient-use efficiency of Pinus thunbergii Parl. along a topographical gradient of soil nutrient availability Ecol Res 12 191–9
JS Gartlan D Newbury C Mc DW Thomas PG Waterman (1986) ArticleTitleThe influence of topography and soil phosphorus on the vegetation of Korup Forest Reserve, Cameroun Vegetatio 65 131–48
TW Giambelluca MA Nullet TA Schroeder (1986) Rainfall atlas of Hawaii Department of Land and Natural Resources Honolulu
AE Giblin KJ Nadelhoffer GR Shaver JA Laundre AJ McKerrow (1991) ArticleTitleBiogeochemical diversity along a riverside toposequence in arctic Alaska Ecol Monogr 61 415–35
RA Harrington JH Fownes FC Meinzer PG Scowcroft (1995) ArticleTitleForest growth along a rainfall gradient in Hawaii: Acacia koa stand structure productivity, foliar nutrients, and water- and nutrient-use efficiencies Oecologia (Berl) 102 277–84
MJ Hedley JWB Stewart BS Chauhan (1982) ArticleTitleChanges in inorganic and organic soil phosphorus fractions induced by cultivation practices and laboratory incubations Soil Sci Soc Am J 46 970–6
DA Herbert JH Fownes (1995) ArticleTitlePhosphorus limitation of forest leaf area and net primary productivity on a weathered tropical soil Biogeochemistry 29 223–35
M Hirobe N Tokuchi G Iwatsubo (1998) ArticleTitleSpatial variability of soil nitrogen transformations along a forest slope in a Cryptomeria japonica D. Don plantation. Eur J Soil Biol 34 123–31
RT Holcomb PW Reiners BK Nelson N-LE Sawyer (1997) ArticleTitleEvidence for two shield volcanoes exposed on the island of Kauai, Hawaii Geology 25 811–4
ML Jackson . Lim LW Zelzny (1986) ArticleTitleOxides, hydroxides, and aluminosilicates. In Methods of soil analysis, Part 1, Physical and mineralogical properties. 2nd edition. Agronomy 9 102–49
H Jenny (1980) Soil genesis with ecological perspectives Springer-Verlag New York
MJ Kennedy OA Chadwick PM Vitousek LA Derry D Hendricks (1998) ArticleTitleReplacement of weathering with atmospheric sources of base cations during ecosystem development, Hawaiian Islands Geology 26 1015–8
AC Kurtz LA Derry OA Chadwick (2001) ArticleTitleAccretion of Asian dust to Hawaiian soils: isotopic, elemental, and mineral mass balances Geochim Cosmochim Acta 65 1971–83
AC Kurtz LA Derry OA Chadwick MJ Alfano (2000) ArticleTitleRefractory element mobility in volcanic soils Geology 28 683–6
IF Lepsch SW Buol RB Daniels (1977) ArticleTitleSoil–landscape relationships in the Occidental Plateau of Sao Paulo State, Brazil. II. Soil morphology, genesis, and classification. Soil Sci Soc Am J 41 109–15
GA McDonald AT Abbot FL Peterson (1983) Volcanoes in the sea: the geology of Hawaii University of Hawaii Press Honolulu 517
JA McKeague JH Day (1966) ArticleTitleDithionite- and oxalate-extractable Fe and Al as aids in differentiating various classes of soils Can J Soil Sci 46 13–22
AJ Miller EAG Schuur OA Chadwick (2001) ArticleTitleRedox control of phosphorus pools in Hawaiian montane forest soils. Geoderma 102 219–37
HL Pearson PM Vitousek (2002) ArticleTitleNitrogen and phosphorus dynamics and symbiotic nitrogen fixation across a substrate age gradient in Hawai’i Ecosystems 5 587–96
Restrepo C, Vitousek P, Neville P. 2003. Landslides significantly alter land cover and the distribution of biomass: an example from the Ninole ridges of Hawai’i. Plant Ecol 166: 131–143
DD Richter D Markewitz (2001) Understanding soil change: soil sustainability over millennia, centuries, and decades. Cambridge University Press Cambridge
RH Riley PM Vitousek (1995) ArticleTitleNutrient dynamics and trace gas flux during ecosystem development in Hawaiian montane rainforest Ecology 76 292–304
PA Sanchez (1976) Properties and management of soils in the tropics John Wiley and Sons New York
FN Scatena AE Lugo (1995) ArticleTitleGeomorphology, disturbance, and the soil and vegetation of two subtropical wet steepland watersheds of Puerto Rico Geomorphology 13 199–213
D Schimel MA Stillwell RG Woodmansee (1985) ArticleTitleBiogeochemistry of C, N, and P in a soil catena of the shortgrass steppe Ecology 66 276–82
PJ Schoeneberger DA Wysocki EC Benham WD Broderson (1998) Field book for describing and sampling soils Natural Resources Conservation Service, USDA, National Soil Survey Center Lincoln, NE
Schuur, E.A.G., Matson, PA (2001) Net primary productivity and nutrient cycling across a mesic to wet precipitation gradient in Hawaiian montane forcits. Occologia 128: 431–442
WL Silver AE Lugo M Keller (1999) ArticleTitleSoil oxygen availability and biogeochemistry along rainfall and topographic gradients in upland wet tropical forest soils Biogeochemistry 44 301–28
WL Silver FN Scatena AH Johnson TG Siccama MJ Sanchez (1994) ArticleTitleNutrient availability in a montane wet tropical forest: spatial patterns and methodological considerations Plant Soil 164 129–45
Soil Survey Laboratory Staff. 1992. Soil survey laboratory methods manual. Soil Survey Investigations Report no. 42, V.2.0, USDA
InstitutionalAuthorNameSPSS (1998) Systat version 8.0. SPSS Chicago
BW Stewart RC Capo OA Chadwick (2001) ArticleTitleEffects of rainfall on weathering rate, base cation provenance, and Sr isotope composition of Hawaiian soils Geochim Cosmochim Acta 65 1087–99
EVJ Tanner (1977) ArticleTitleFour montane rain forests of Jamaica: a quantitative characterization of the floristics, the soils and the foliar mineral levels, and a discussion of the interrelations J Ecol 65 883–918
H Tiessen JO Moir (1993) Characterization of available P by sequential extraction. MR Carter (Eds) Soil Sampling and methods of analysis. Louis Boca Raton, FL 75–86
Van den Driessche, R. (1974) Prediction of mineral nutrient status of trees by foliar analysis. bot. Rev. 40: 347–394
PM Vitousek G Aplet DR Turner JJ Lockwood (1992) ArticleTitleThe Mauna Loa environmental matrix: foliar and soil nutrients Oecologia (Berl) 89 372–82
PM Vitousek TL Benning (1995) Ecosystem and landscape diversity: islands as model systems. PM Vitousek LL Loope H Adsersen (Eds) Biological diversity and ecosystem function on islands. Springer-Verlag Berlin 73–84
PM Vitousek OA Chadwick T Crews J Fownes DM Hendricks D Herbert (1997) ArticleTitleSoil and ecosystem development across the Hawaiian Islands. GSA Today 7(9) 1–8
PM Vitousek DR Turner K Kitayama (1995) ArticleTitleFoliar nutrients during long-term soil development in Hawaiian montane rain forest Ecology 76 712–20
LR Walker DJ Zarin N Fetcher RW Myster AH Johnson (1996) ArticleTitleEcosystem development and plant succession on landslides in the Caribbean Biotropica 28 566–76
TW Walker JK Syers (1976) ArticleTitleThe fate of phosphorus during pedogenesis Geoderma 15 1–19
DJ Zarin AH Johnson (1995) ArticleTitleNutrient accumulation during primary succession in a montane tropical forest, Puerto Rico. Soil Sci Soc Am J 59 1444–52
Acknowledgements
We thank the Joseph Souza Center and the Hawaii Department of Land and Natural Resources’ State Parks and Forestry and Wildlife Divisions for access to field sites and logistical support; Heraldo Farrington and David Penn for assistance in the field; and Douglas Turner, Adina Payton, and Bettina Wiegand for help with laboratory analyses and Sr isotope determinations. We thank Dan Richter for a very helpful review of an earlier draft of this article. This research was supported by a grant from the Andrew W. Mellon Foundation, and by NSF grant DEB-0108492.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Vitousek, P., Chadwick, O., Matson, P. et al. Erosion and the Rejuvenation of Weathering-derived Nutrient Supply in an Old Tropical Landscape. Ecosystems 6, 762–772 (2003). https://doi.org/10.1007/s10021-003-0199-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10021-003-0199-8