
Abstract
Land-use change in the tropics is creating secondary forest at an unprecedented rate. In the tropical Americas, mature dry tropical forest is rapidly being converted to secondary forest during the fallow period of shifting cultivation. We investigated litter phosphorus (P) and nitrogen (N) dynamics in forests recovering from shifting cultivation of maize (corn) in three regions of the Southern Yucatan Peninsula, Mexico. Our goal was to understand how nutrient and water availability affect forest recovery following conversion of mature forest to agricultural land. To investigate such changes at a regional scale, newly fallen litter was collected monthly along a seasonal, a successional, and a precipitation gradient. Reflecting possible P limitation, litter P concentration declined with forest age, while litter N concentration did not differ between age classes. Average litter P concentration from the southern, wettest region was 0.87 mg/g, almost twice the litter P concentration in the drier central and northern regions (0.44 and 0.45 mg/g, respectively). Average N concentrations of litter from the three regions ranged from 1.1% to 1.2%, with no regional differences. However, minima in both P and N concentration from all regions were pronouncedly timed with peak litterfall, suggesting nutrient retranslocation during periods of water stress. Additionally, successional differences in litter P were clearest during wetter months. P nutrient-use efficiency was lowest in the southern region and highest in the central and northern study regions. N nutrient-use efficiency was up to 40 times lower than P nutrient-use efficiency and showed no regional differences. Overall, our results suggest that litter nutrient dynamics in secondary dry tropical forests of the Southern Yucatan are strongly influenced by water and nutrient availability, especially P, as well as land-use history.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Concern over the global implications of tropical deforestation has focused attention on the sparse literature on ecosystem dynamics in both secondary tropical forests and dry tropical forests (Arnason and Lambert 1982; Murphy and Lugo 1986a; Brown and Lugo 1990; Campo and others 2001). In many areas of the tropics, mature forest is being converted to secondary forest during the fallow period of shifting cultivation and following pasture abandonment. Secondary forests comprise 40% of the total tropical forest area and are being formed at a rate of approximately 9 million ha/year (Brown and Lugo 1990). These forests play an important role in global carbon dynamics, due in part to their high rates of growth and nutrient turnover compared to mature forests (Uhl and Jordan 1984; Murphy and Lugo 1986b; Brown and Lugo 1990; Hughes and others 1999).
Despite their rapid rate of formation and areal extent, ecosystem dynamics in secondary tropical forests remain largely undercharacterized. Because litter production and litter nutrients are important drivers of soil chemistry, their dynamics have a direct impact on the recovery of soil fertility during the fallow period of shifting cultivation (Nye and Greenland 1960; Ewel 1976; Lambert and others 1980; Lugo 1992). Intensity of prior disturbance, such as repeated cultivation and fallow periods, may also affect the rate and dynamics of nutrient cycling in secondary tropical forests (Uhl and Jordan 1984; Buschbacher and others 1988; Hughes and others 1999; Lawrence and Schlesinger 2001). Therefore, ecosystem-level studies of nutrient cycling in secondary tropical forests will aid in estimation of how long it takes for cultivated soils to return to nutrient levels capable of sustaining agriculture and continued increases in human population.
The role of litter nutrients in secondary forest recovery may be especially critical in dry tropical forests, where seasonal pulses of nutrients in litterfall constitute one of the most important aspects of the entire nutrient cycle (Campo and others 2001). Dry forests, which comprise 42% of the total forested area in the tropics (Brown and Lugo 1982), receive less scientific and media attention compared to the wet tropics, yet are important components of changing global biogeochemical cycles because of their areal extent and suitability for human habitation (Murphy and Lugo 1986a). Due to the predominant role of water limitation in driving ecosystem dynamics of seasonally dry tropical forests, litter nutrient dynamics in these forests have been poorly studied (Murphy and Lugo 1986a, 1986b; Campo and others 2001). In this study, we investigated litter phosphorus (P) and nitrogen (N) dynamics in successional and mature dry tropical forests of the Southern Yucatan Peninsular Region (SYPR). Our overall goal was to understand the effects of seasonal, successional, and regional precipitation gradients on litter quality during recovery of secondary forests following shifting cultivation of maize. Our specific objectives were (a) to investigate how litter nutrient quality varies during the year, and whether these seasonal dynamics depend on forest age or annual precipitation, and (b) to explore how litter nutrient concentrations, P and N inputs to the forest floor, and nutrient-use efficiency change during forest recovery, and how these dynamics are affected by a regional precipitation gradient.
Previous work in the dry tropics suggests that forest structure and function are primarily driven by water limitation and seasonal drought (Lugo and Murphy 1986). Annual litter production and the litter nutrient cycle in dry tropical forests are very dynamic, especially in areas with highly variable precipitation (Lambert and others 1980; Swift and others 1981; Lugo and Murphy 1986; Martinez-Yrizar and Sarukhan 1990; Whigham and others 1990). Precipitation patterns in the dry tropics are especially important because they are often tied to biological patterns in growth, litterfall, and reproduction (Murphy and Lugo 1986a). Nutrient limitation in the dry tropics is related to water limitation because dry conditions prevent plant uptake of available nutrients from soil, and affect the release and mineralization of nutrients by slowing decomposition. It is widely believed that many dry tropical forests may be especially limited by P because of its adsorption to clay and calcium in carbonate-rich soils such as those of the SYPR (Vitousek 1984; Lugo and Murphy 1986; Silver 1994; Campo and others 2001).
Despite their areal and ecological importance, tropical secondary and dry tropical forests are understudied compared to mature wet tropical forests (Murphy and Lugo 1986a; Brown and Lugo 1990; Martinez-Yrizar and Sarukhan 1990; Castellanos and others 1991; Kauffman and others 1993; Campo and others 2001). In this study, we quantified litter nutrient dynamics in secondary dry tropical forest of the Yucatan to assess the role of water and nutrient availability on forest recovery following human disturbance. This work enables a better understanding of the role of dry tropical forests in global biogeochemical cycles, and how these dynamics are impacted by human activity.
Methods
Study Area
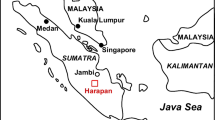
Litter collection sites were located within three study regions in the SYPR (Figure 1 and Table 1): northernmost El Refugio (18°49′N, 89°23′W), centrally located Nicolas Bravo (18°27′N, 88°56′W), and southernmost Arroyo Negro (17°53′N, 89°17′W). The study regions are located 60–120 km apart and situated along a precipitation gradient of approximately 900 mm ppt)/year (El Refugio) to 1400 mm ppt/year. Within each of the three study regions, 10–13 500-m2 circular plots were established in a successional sequence of secondary forest ranging from 2 to 25 years since abandonment from maize agriculture, plus several mature forests of unknown age (Table 1). Prior to recovery, the secondary sites had been used for maize cultivation with no chemical inputs, and most were probably subject to periodic logging before agricultural use. Therefore, mature stands in our study represent forests in a precultivation state, which may have been subject to logging but have not been used for agriculture since Mayan times. The sites were chosen according to their ecological similarity and cultivation history following extensive interviews with villagers (Klepeis 2000). Sites cultivated by the same family but of different successional age were chosen to minimize management-induced and edaphic variation.
The climate of the Yucatan Peninsula is characterized by highly variable precipitation over seasonal and interannual time scales and a mean annual temperature of 25°C (Harmon and others 1995). In our study region, peak rainfall occurs in September/October, and the rainy season extends from July to December (Turner and others 2001). The dry season (defined as < 50 mm/month) typically lasts 3 months in Arroyo Negro, 3–4 months in Nicolas Bravo, and 6 months in El Refugio, during which time 35–40 mm/month falls in all three regions [data courtesy of INEGI (El Instituto Nacional de Estadística, Geografía e Informática), México]. During wet-season months, precipitation is heaviest in the south (136 mm/month in Arroyo Negro), decreasing northward (118 mm/month in Nicolas Bravo and 95 mm/month in El Refugio), further compounding differences in annual precipitation caused by differences in the length of the wet season. Peak litterfall occurs in March at all study sites, although seasonal changes in litter production were least pronounced in Arroyo Negro, the southern, wettest region (Lawrence and Foster 2002). Generally, soils in the Yucatan are shallow, calcareous, and highly permeable due to high organic matter content and underlying limestone bedrock (Whigham and others 1990). Preliminary soil analyses indicate that both sand and organic matter content increase along gradients of forest age and precipitation (Lawrence and Foster 2002). Organic matter may increase with sand content if a clay-rich layer prevents rapid drainage. It may also bind smaller particles together, resulting in higher numbers of sand-sized particles (0.05–2.0 mm). Natural disturbances in the region include fires and hurricanes, such as Hurricane Janet in 1955, Gilbert in 1988, and Isidore in 2002 (Boose and others 1994; Harmon and others 1995).
Although human disturbance, including logging, chicle (latex gum) production, and farming, has contributed to land-cover change in the Southern Yucatan since the early 1900s, the land was also used by the Mayans for intensive agriculture until approximately 1100 years ago (Turner 1974; Klepeis 2000). Government-sponsored development initiatives, coupled with the opening of a highway through the region in 1967, led to immigration of colonists from other regions of Mexico. In the 1970s, rapid conversion of mature forest to agricultural land was spurred by growth of the ejido system, a collective farming initiative promoted by the government. In the future, human population and land-use change is expected to increase because of efforts to shift the current agricultural system from subsistence to market-oriented farming (Klepeis 2000; Turner and others 2001).
Sampling Regime and Nutrient Analyses
Newly fallen fine litter, consisting of leaves, small reproductive parts, twigs, and small branches less than 2 cm in diameter, was collected monthly from litter traps and analyzed for N and P. Four 1-m2 litter traps, made of 1 × 1-mm nylon mesh, were arrayed orthogonally 8 m from the center in each of the thirty-six 500-m2 plots. The traps were positioned 10–20 cm off the ground to prevent soil moisture from affecting litter chemistry between collections, and trap positions remained the same during the entire sampling year. Composites of the four samples per plot were analyzed for the year between January and December 1999. During this sampling year, there were 11 missing litter composites (that is, all traps at a plot were stolen) out of 432. For these missing data points, we used an average nutrient concentration computed from the preceding and subsequent values at the respective plot. In the case of several, but not all, traps being stolen at a plot, the composite was made with the litter collected from the remaining traps at that plot. Because decomposition is relatively slow in dry tropical forests (Arnason and Lambert 1982; Lugo and Murphy 1986; Harmon and others 1995), we believe that our monthly collections did not result in substantial changes in litter chemistry. Nutrient loss due to leaching may have affected litter nutrient content, especially during the wet season, with N being more susceptible to losses than P (see Cuevas and Medina 1986).
The litter was dried in the field at less than 70°C. It was ground through a Wiley Mill (Thomas Scientific, Swedesboro, NJ, USA) to pass through a no. 20-mesh screen. Litter P and N concentrations were determined using a modified Kjeldahl digestion on a Tecator 2000 Digestion System (Perstorp Analytical, Sweden). This method uses a concentrated salicylic acid–sulfuric acid catalyst and 30% hydrogen peroxide to hydrolyze organic P and N to inorganic forms. The digestion matrix was analyzed on an Alpkem Flow Solution IV Autoanalyzer (OI Analytical, College Station, Texas, USA) in accordance with the US EPA methods for total P and Kjeldahl N determination. An estimate of litter C concentration was determined by combustion of 59 random samples out of a total of 432. Litter C concentration was not measured for all samples because of the negligible variability in C content of the litter. Ground samples were dried at 50°C for 48 h prior to being weighed for combustion in a Fison 1108 Elemental Autoanalyzer (CE Elantech, Lakewood, NJ, USA). Measurements of litter C content were used to calculate mass-based C:N and C:P ratios for secondary and mature sites in each region as another estimate of litter quality.
Total nutrient inputs (kg ha−1 year−1) were calculated by multiplying monthly litter production values for each sampling site (Lawrence 2004) by nutrient concentration for the same site and month, and adding them over the entire year. These annual nutrient input values were then used to calculate within-stand nutrient-use efficiency (NUE), defined as kilograms of dry litter mass per kilograms of litter nutrient content (sensu Vitousek 1982, 1984). This index was developed as an improvement over using the simple inverse of plant-tissue nutrient concentration as originally suggested by Chapin (1980; Vitousek 1982). We chose this index of NUE to assess stand nutrient cycling due to the nature of our data set, the relative ease of studying litter nutrients compared to other components of the cycle, and the assumption that an understanding of the litter nutrient cycle may serve as a model for the whole stand nutrient cycle (Vitousek 1982, 1984; Silver 1994). Litterfall is the predominant pathway through which N and P travel from plants to soil (Vitousek 1982). We also investigated regional differences in resorption proficiency, defined as the minimum nutrient concentrations in senesced leaves (Killingbeck 1996), in mature forest sites.
We investigated percent reduction in litter nutrient concentrations among regions and age classes in order to compare the degree to which litter P and N concentrations changed during the annual cycle. The percent reduction from maximum to minimum nutrient concentration and the percent reduction from maximum nutrient concentration to that occurring during peak litterfall were calculated for each site. We did not attempt to quantify true nutrient resorption from live to senesced leaves because our sampling regime did not permit this type of analysis. However, our assessment of the relative changes in litter nutrient concentrations during the annual cycle provides a similarly comprehensive perspective on nutrient cycling, because it accounts for relative changes in the nutrient content of newly fallen litter in forests that receive markedly different annual precipitation.
Statistical Analyses
We explored seasonal patterns in nutrient concentrations as a function of age and precipitation by using repeated-measures analysis of variance (ANOVA) with each of the 12 sampling periods as the repeated measure. Regional patterns in mean annual litter nutrient concentrations and NUE were analyzed using simple ANOVA. To investigate successional changes in litter nutrient concentrations over all regions combined, we used two-way ANOVA to allow for attribution of variance to regional differences in litter nutrients over the precipitation gradient. Linear regressions were used to model age-related changes in litter nutrient concentration and NUE in secondary forests of each region. Regional comparisons in percent reduction from maximum to minimum or peak litterfall levels were carried out using ANOVA.
For analytical purposes, the 36 study sites were grouped across all three regions into age classes that best represent their ecological similarity and take into account any uncertainty in exact forest age. These age classes (2–5 years, 6–10 years, 12–25 years, and mature forest) refer to stand age in January 1999. All statistical analyses were carried out using SAS V8 (SAS Institute 1989).
Results
Seasonal Patterns in Litter Nutrient Concentration
Across all regions and ages, litter P and N concentration changed significantly through time (time effect, P < 0.0001). Litter nutrient concentrations reached their minima during peak litterfall in March and were at maximum levels during the wet season, July–December (Figures 2 and 3). Seasonal patterns in litter P concentrations varied as a function of both age and region in separate analyses (time*region effect, P < 0.01; and time*age-class effect, P = 0.03). Arroyo Negro showed a more marked decline during the dry season and did not vary as much during the wet season (Figure 2a). Phosphorus concentration peaked in mature forests earlier than in younger secondary forests (Figure 3a). In contrast, seasonal patterns in litter N concentrations varied as a function of region (time*region effect, P < 0.01) but not as a function of age (time*age-class effect, P = 0.13). The significant regional effect seems to be caused by erratic variation in El Refugio.
During peak litterfall in March, litter P and N concentrations were reduced an average of 47% and 33%, respectively, from their maximum levels, which always occurred during the wet season. The greatest proportional reduction in litter P concentration (from maximum observed to peak litterfall value) generally occurred in the driest region, despite a greater absolute change in the wettest region [Arroyo Negro (Figure 2a)]. Older forests also tended to experience greater proportional change than younger forests (Figure 3a). When comparing litter P maximum versus absolute minimum, regardless of when that minimum occurred during the annual cycle, the reduction was greatest in the driest region as well (P = 0.07) (Figure 4a). In contrast, there were no strong regional or successional patterns of relative reduction in litter N concentration, whether comparing maximum to peak litterfall levels (Figures 2b and 3b) or maximum to absolute minimum (Figure 4b). Not only the greatest relative reduction in, but also the lowest minimum, litter P concentration in mature forests (0.22 mg/g) occurred at a site in the driest region, El Refugio, whereas the highest minimum value occurred in an Arroyo Negro site (0.37 mg/g). In contrast, minima in litter N concentrations showed no regional trend, ranging from 0.62% to 0.96% with high intraregional variability.
Regional and Successional Trends in Litter Nutrient Concentration
Litter P concentration in the southern, wettest site, Arroyo Negro, was significantly higher than that of Nicolas Bravo and El Refugio over the entire course of the annual cycle (regional effect, P < 0.0001) (Figure 2a). Mean litter P concentration was 0.87 mg/g in Arroyo Negro versus 0.44 and 0.45 mg/g in Nicolas Bravo and El Refugio, respectively. Litter N concentrations were not significantly different among regions (regional effect, P = 0.75). Litter C:N ratios in the SYPR ranged from 40 to 44, whereas C:P ratios ranged more widely, from 470 to 1220 (Table 2).
Litter N concentration did not vary significantly, though it tended to increase, with forest age (Figure 5b). Mean litter P concentration, averaged over all three regions, declined with forest age from 0.65 mg/g in the youngest age class to 0.50 mg/g in the mature sites (age-class effect, P = 0.06) (Figure 5a). Although the means show an effect of age class on litter P concentration, the repeated-measures analysis over the entire annual cycle does not (age-class effect, P = 0.41). The age-class effect is obscured because during the dry season litter P converges on the same low level regardless of age class (Figure 3a).
We further examined the association between P dynamics and precipitation indicated by a decline in litter P concentration during dry months and in dry regions (Figures 2a and 3a). The effect of age on litter P was much stronger during the wet season, June–December, than it was over the complete annual cycle (age-class effect, P = 0.01 for the wet season only and P = 0.06 for the entire year). Furthermore, the decrease in mean litter P concentration with forest age over all sites is likely driven by the wettest region, Arroyo Negro, where P concentration declined most with age. Regression analyses relating average P concentration to age in secondary forests indicate a negative relationship in Arroyo Negro (R 2 = 0.47 and P = 0.06). However, in secondary forests of centrally located Nicolas Bravo and the driest site El Refugio, litter P concentrations were not significantly related to age.
Total Litter Nutrient Inputs and Nutrient-use Efficiency
Total annual P input ranged from 3.01 kg P ha−1 year−1 in a 2-year-old forest in El Refugio to 14.45 kg P ha−1 year−1 in a mature forest in Arroyo Negro (Table 3). Across all age classes, total P inputs from litter were significantly higher in Arroyo Negro than in Nicolas Bravo and El Refugio (P < 0.0001). Mean P inputs in mature forests were 9.6, 4.9, and 5.8 kg P ha−1 year−1, respectively. Nitrogen inputs ranged from 59.8 kg N ha−1 year−1 to 204.3 kg N ha−1 year−1, with substantial variability within regions and age classes (Table 3). However, there were no significant regional or age-related differences in litter N inputs to the forest floor.
Overall, P NUE increased with decreasing precipitation (Figure 6a and Table 3). Phosphorus NUE over the SYPR ranged fourfold, from 913 to 3964 kg litter/kg P, while N NUE spanned a much narrower range, 68 to 129 kg litter/kg N. Regional differences in P NUE were large, with average P NUE in Arroyo Negro significantly lower than in the drier regions of Nicolas Bravo or El Refugio (1279 vs 2488 and 2509 kg litter/kg P, respectively) (P < 0.0001) (Table 3). There were no significant differences in N NUE among regions (Figure 6b and Table 3). Regression analyses indicate that, in Arroyo Negro, P NUE increased significantly with increasing forest age (R 2 = 0.62 and P = 0.02). However, there were no significant relationships between forest age and P NUE for El Refugio and Nicolas Bravo, nor were there any significant relationships between N NUE and forest age in any of the study regions.
Discussion
Regional and successional litter nutrient dynamics
In the SYPR, there is a strong association between litter P concentration and precipitation on a regional scale. Litter P concentration was higher in the wet southern region, compared to the drier central and northern sites (Figure 2a). Although the distance between study areas is at most 120 km, the 900- to 1400-mm/year gradient in total annual precipitation appears to be an important driver of forest nutrient status. Regional variability in precipitation may result in a threshold (perhaps around 1200 mm/year), below which P limitation is continuous, and above which P limitation is seasonal. The lack of a simple gradient effect of rainfall on litter P concentration and P NUE is evidenced by the fact that Arroyo Negro sites drive regional and successional patterns in these metrics. Additionally, litter P concentration and P NUE in Nicolas Bravo are not intermediate between El Refugio and Arroyo Negro, but are similar to El Refugio (Figures 2a and 6a).
Phosphorus return from litter in our mature study sites is higher than that reported for dry forests in Chamela, Mexico (679 mm ppt/year and 3.88 kg ha−1 year−1), but lower than that reported for a seasonal forest in Belize (1720 mm ppt/year and 9 kg ha−1 year−1) (Lambert and others 1980; Campo and others 2001). Nitrogen return from litter in our mature sites is comparable to those reported for Guatemala (2000 mm ppt/year and 169 kg ha−1 year−1) and Belize (156 kg ha−1 year−1) (Ewel 1976; Lambert and others 1980).
Preliminary analysis of species composition data (Table 4) indicates a high degree of similarity in forest composition and species abundance in both secondary and mature sites among the three study areas (D. Perez Salicrup and D. Lawrence unpublished data). However, differences in community structure cannot be completely discounted as a potential factor contributing to differences in litter nutrient status at the regional scale. Forests in Nicolas Bravo seem to share fewer species than Arroyo Negro and El Refugio, the end points of our precipitation gradient. This compositional difference may explain why the precipitation effect is not gradual, as expected, but more of a threshold.
During succession, litter P concentration declines, while litter N concentration does not change significantly (Figures 3 and 5), suggesting that P is limiting compared to N and is cycled more efficiently as the forests age (Vitousek 1982). In his study of successional forest in Guatemala, Ewel (1976) also reports that litter N concentration did not decline with stand age. Some of this change may be a result of species turnover as a forest ages (Table 4), although the specifics of such change are difficult to define given our use of composite litter samples. However, changes in nutrient-use efficiency during succession may also reflect physiological change within the species themselves, as many species persist over the entire successional sequence sampled. Furthermore, the relative influence of forest age on litter nutrient dynamics is affected by both seasonal and regional variability in precipitation. During the rainy season (July–December), successional differences in litter P concentration were more apparent than they were during the dry season (Figure 3a). Additionally, differences in litter P concentration due to forest age were most pronounced in the wettest site, Arroyo Negro, whereas this trend was not as strong in the other study regions. In contrast, the variability in litter N concentrations was not attributable to differences in age or region, regardless of seasonal precipitation patterns (Figures 2b and 3b). Overall, these results suggest that forest function in the SYPR is primarily limited by water availability with secondary limitation by P.
Seasonal Trends in Litter Nutrient Concentrations
Seasonal trends in litter nutrient dynamics also indicate a strong relationship between precipitation and litter P concentration. During March–April, the period of peak litterfall, litter N and P levels were at their lowest across all regions and age classes (Figures 2 and 3). These dynamics may reflect retranslocation of essential nutrients during times of drought. This retranslocation may be an adaptive process that is more energy efficient than taking up nutrients from the soil (Potter and others 1987). It may also be a nutrient-conservation mechanism for plants in low-nutrient environments (Vitousek 1982; Killingbeck 1996), even if the nutrient stress is caused simply by lack of soil moisture.
When litter nutrient dynamics are considered in the context of concurrent changes in litter mass (Figure 7a) (D. Lawrence unpublished manuscript), nutrient supply appears to be more uniform in younger than in older forests (Figure 7b). During peak litterfall, when nutrient concentrations are lowest, variation in nutrient input is driven by differences in total litter production. With more litterfall at that time, mature forests deposit roughly 40% more P than the youngest forests. In contrast, during the wet months (July–December), litter production does not vary with forest age. Differences in total inputs are driven by concentration, and the younger forests deposit 20%–90% more P than mature forests. These higher nutrients come when soil moisture is high, speeding decomposition and promoting rapid release into the soil solution. Thus, enhanced nutrient availability during the wet season may encourage rapid growth in young forests, enhancing recovery after human or natural disturbance.
The greatest proportional reductions in litter P concentration during peak litterfall were generally in the driest region, El Refugio (Figure 4a), suggesting that water limitation contributes to greater nutrient-use proficiency in El Refugio than in the other study regions. In mature study sites, minima in litter P also declined with decreasing precipitation, which further suggests that removal of essential nutrients such as P prior to senescence may be driven by water limitation. Additionally, relative reduction in litter nutrients during peak litterfall tended to be higher in older forests, consistent with the theory that the cycling of limiting nutrients becomes more conservative as forests age. Our results suggest the interaction of both seasonal and regional water limitation and successional change in the ability of trees to retranslocate nutrients.
Nutrient-use Efficiency
Phosphorus NUE was lowest in Arroyo Negro and highest in the driest site, El Refugio (Figure 6a), suggesting that P is less limiting in this system where water is more available. Phosphorus NUE increased significantly with forest age only in Arroyo Negro. In the drier sites, P NUE was high regardless of forest age. However, the lack of a simple gradient between water availability and P NUE is illustrated by the fact that P NUE in Nicolas Bravo is not clearly intermediate between El Refugio and Arroyo Negro (Table 3). This may be driven by a combination of differences in species composition (Table 4) and litterfall mass and seasonality (Figure 7).
In contrast, N NUE displayed no regional or successional trends. Nitrogen NUE was 20 times lower, on average, than P NUE (Figure 6b and Table 3), suggesting the relative degree of N and P availability to plants. High P NUE and a lack of regional and successional effects on N NUE suggest that P is the limiting nutrient in this system, as indicated for other tropical forests (Vitousek 1984; Lugo 1992; Silver 1994). Phosphorus cycling in the tropics is believed to be largely moderated by weathering processes rather than through atmospheric and biological processes, although recent work in secondary tropical forests suggests an important role for biota (McGrath and others 2000; Lawrence and Schlesinger 2001). In highly calcareous soils such as those in the SYPR, P may be chemically bound to calcium and clay constituents (Vitousek 1984; Silver 1994). Phosphorus availability in dry tropical soils may be further reduced because xeric conditions prevent dissolution and plant uptake of P bound to the soil (Lugo and Murphy 1986). This condition would be exaggerated in our driest sites, producing the regional and seasonal patterns in litter P concentration and P NUE we found. In contrast, the N cycle is sustained to a large extent by atmospheric and biological processes, the latter of which may be augmented by abundant microbial N fixation in warm tropical soils (Vitousek 1984).
High P NUE compared to N NUE has been described for numerous tropical forests, ranging from very dry to wet montane sites [see Vitousek (1984), Lugo and Murphy (1986), and Singh (1992)]. Our results suggest P limitation and relatively abundant N, within the constraints of highly variable regional and seasonal precipitation. Additional variability in P and N NUE may be driven in part by patterns in litter production at each site (D. Lawrence unpublished manuscript). For example, the low input of 4.65 kg P ha−1 year−1 in one mature site in Arroyo Negro is due to notably small biomass and litter production at that site (Read and Lawrence 2004). It is also possible that regional and successional differences in NUE are due in part to the interplay of two factors whose relative contributions are difficult to quantify: the shift from less-efficient to more-efficient species along those gradients, and increasing efficiency within species as they respond to reduced nutrient availability or other stresses (Boerner 1984).
Conclusion
We investigated litter nutrient dynamics in the SYPR because litter is one of the most labile components of the nutrient cycle and has a direct effect on soil fertility, and thus plays a critical role in the recovery of forests following cultivation. In so doing, we have provided an understanding of ecosystem-level response to natural and human-induced environmental gradients in the dry tropics. Litter N and P concentrations have markedly different dynamics over seasonal, successional, and regional scales. Our study suggests that water limitation and P limitation may serve as important controls on ecosystem nutrient cycling in and recovery of these forests. When forests in the SYPR are released from water stress—that is, during the rainy season or in a wetter region—litter nutrients are sustained at levels higher than those detected during the dry season or in regions with lower annual precipitation. Phosphorus-use efficiency tends to diminish with increasing precipitation, as indicated by comparing stand-level NUE, percent reduction in litter P, and resorption proficiency. This work highlights the importance of regional environmental gradients, such as precipitation, which are seemingly small in comparison to global gradients, but that may exert control over ecosystem properties on a regional scale. During this period of rapid land-use change in the tropics, and in anticipation of projected global environmental change, it is critical that we understand nutrient dynamics in both mature dry tropical systems and in the secondary forests that are becoming an increasingly important part of the tropical landscape.

Map of study areas in the Southern Yucatan Peninsular region (SYPR), Mexico. As point of reference, shaded areas indicate location of Calakmul Biosphere Reserve.

Seasonal trends in litter phosphurus (a) and nitrogen (b) concentration by region. Monthly means ± 1 S.E. for 10 sites in Arroyo Negro (AN), 13 sites in Nicholas Bravo (NB), and 13 sites in El Refugio (ER). The magnitude of the seasonal trend in P in Arroyo Negro is significantly greater than that of Nicholas Bravo and El Refugio (repeated measures ANOVA, regional effect, P<0.0001). Seasonal N dynamics were not significantly different among regions. Minima in litter P during March correspond to peak litterfall. Note different scales on the y-axes.

Seasonal trends in litter phosphorus (a) and nitrogen (b) concentration by age class. Points are monthly means ± 1 S.E. for 10 2–5 year-old sites, 8 6–10 year old sites,, 10 12–25 year-old sites, and 8 mature sites, combined across all regions.There were no significant differences between age classes for P or N (repeated measures ANOVA, age class effect, P; p=0.41, N: p=0.75)

Percent reduction in litter phosphorus (a) and nitrogen (b) concentrations by region, across all age classes. For each site, the percent reduction in nutrient concentration from maximum to minimum levels during the annual cycle was calculated, and regional means were compared using ANOVA. Bars are means ± 1 S.E. Percent reduction in litter P increases with decreasing precipitation from Arroyo Negro to El Refugio (ANOVA, p=0.07). Percent reduction in N does not vary significantly by region (ANOVA, p=0.94).

Average litter phosphorus (a) and nitrogen (b) concentrations across all regions, as a function of forest age. Bars indicate means ± 1 S.E. over the entire annual cycle. Although data presented here illustrate successional changes only, we used Two-way ANOVA to allow for attribution of variance to the regional gradient. The data indicate a decline in litter P (model, P<0.0001; age class effect, p=0.06) but no significant change in litter N (model, p=0.96) with increasing forest age.

Nutrient use efficiency as a function of forest age, for phosphorus (a) and nitrogen (b). Annual NUE was calculated as dry litter mass/litter nutrient content (kg/kg). Phosphorus NUE was significantly lower in Arroyo Negro (AN) then in El Refugio (ER) and Nicholas Bravo (NB) (ANOVA, p<0.0001). In Arroyo Negro, P NUE increased with forest age in secondary sites ( P Nue=41.48*age + 754.90, R2=0.62, p=0.02). There were no regional or successional differences in N NUE.

Seasonal trends in litterfall mass (a) and P return from litter (b) as a function of forest age. Mean over all three regions is presented. In (b), horizontial lines indicate mean monthly p input for each age class and vertical lines, to scale, indicate the standard deviation in monthly P input for the year.
References
JT Arnason JDH Lambert (1982) ArticleTitleNitrogen cycling in the seasonally dry forest zone of Belize, Central America. Plant Soil 67 333–42
REJ Boerner (1984) ArticleTitleFoliar nutrient dynamics and nutrient use efficiency of four deciduous tree species in relation to site fertility. J Appl Ecol 21 1029–40
ER Boose DR Foster M Fluet (1994) ArticleTitleHurricane impacts to tropical and temperate forest landscapes. Ecol Monogr 64 369–400
S Brown AE Lugo (1982) ArticleTitleThe storage and production of organic matter in tropical forests and their role in the global carbon cycle. Biotropica 14 161–87
S Brown AE Lugo (1984) ArticleTitleBiomass of tropical forests: a new estimate based on forest volumes. Science 223 1290–3
S Brown AE Lugo (1990) ArticleTitleTropical secondary forests. J Trop Ecol 6 1–32
R Buschbacher C Uhl EAS Serrao (1988) ArticleTitleAbandoned pastures in eastern Amazonia. II. Nutrient stocks in the soil and vegetation. J Ecol 76 682–99
MA Cairns PK Haggerty R Alvarez BHJ De Jong I Olmsted (2000) ArticleTitleTropical Mexico’s recent land-use change: a region’s contribution to the global carbon cycle. Ecol Appl 10 1426–41
J Campo M Maass VJ Jaramillo A Martinez-Yrizar J Sarukhan (2001) ArticleTitlePhosphorus cycling in a Mexican tropical dry forest ecosystem. Biogeochemistry 53 161–79
J Castellanos M Maass J Kummerow (1991) ArticleTitleRoot biomass of a dry deciduous tropical forest in Mexico. Plant and Soil 131 225–228
FS Chapin (1980) ArticleTitleThe mineral nutrition of wild plants. Annu Rev Ecol Syst 11 233–60
E Cuevas E Medina (1986) ArticleTitleNutrient dynamics within Amazonian forest ecosystems. I. Nutrient flux in fine litter fall and efficiency of nutrient utilization. Oecologia (Berl) 68 466–72
JJ Ewel (1976) ArticleTitleLitter fall and leaf decomposition in a tropical forest succession in eastern Guatemala. J Ecol 64 293–307
ME Harmon DF Whigham J Sexton I Olmstead (1995) ArticleTitleDecomposition and mass of woody detritus in the dry tropical forests of the northeastern Yucatan Peninsula, Mexico. Biotropica 27 305–16
RF Hughes JB Kauffman VJ Jaramillo (1999) ArticleTitleBiomass, carbon, and nutrient dynamics of secondary forests in a humid tropical region of Mexico. Ecology 80 1892–907
RF Hughes JB Kauffman VJ Jaramillo (2000) ArticleTitleEcosystem-scale impacts of deforestation and land use in a humid tropical region of Mexico. Ecol Appl 10 515–27
JB Kauffman RL Sanford Jr DL Cummings IH Salcedo EVSB Sampaio (1993) ArticleTitleBiomass and nutrient dynamics associated with slash fires in neotropical dry forests. Ecology 74 140–51
KT Killingbeck (1996) ArticleTitleNutrients in senesced leaves: keys to the search for potential resorption and resorption proficiency. Ecology 77 1716–29
Klepeis P. 2000. Deforesting the once deforested: land transformation in southeastern Mexico [dissertation]. Worchester (MA): Clark University.
JDH Lambert JT Arnason JL Gale (1980) ArticleTitleLeaf litter and changing nutrient levels in a seasonally dry tropical hardwood forest, Belize, C.A. Plant Soil 55 429–43
D Lawrence (2004) ArticleTitleRegional and local determinants of seasonal litterfall patterns in the dry tropical forests of the Southern Yucatan. . . .
D Lawrence DR Foster (2002) ArticleTitleChanges in forest biomass, litter dynamics and soils following shifting cultivation in southern Mexico: an overview. Interciencia 27 400–8
D Lawrence WH Schlesinger (2001) ArticleTitleChanges in soil phosphorus during 200 years of shifting cultivation in Indonesia. Ecology 82 2769–80
AE Lugo (1992) ArticleTitleComparison of tropical tree plantations with secondary forests of similar age. Ecol Monogr 62 1–41
AE Lugo PG Murphy (1986) ArticleTitleNutrient dynamics of a Puerto Rican subtropical dry forest. J Trop Ecol 2 55–72
A Martinez-Yrizar J Sarukhan (1990) ArticleTitleLitterfall patterns in a tropical deciduous forest in Mexico over a five-year period. J Trop Ecol 6 433–44
DA McGrath NB Comerford ML Duryea (2000) ArticleTitleLitter dynamics and monthly fluctuations in soil phosphorus availability in an Amazonian agroforest. For Ecol Manag 131 167–81
PG Murphy AE Lugo (1986a) ArticleTitleEcology of tropical dry forest. Annu Rev Ecol Syst 17 67–88
PG Murphy AE Lugo (1986b) ArticleTitleStructure and biomass of a subtropical forest in Puerto Rico. Biotropica 18 89–96
Nye PH, Greenland DJ. 1960. The soil under shifting cultivation. Technical Comment No. 51. Harpenden (England): Commonwealth Bureau of Soils.
CS Potter HL Ragsdale CW Berish (1987) ArticleTitleResorption of foliar nutrients in a regenerating southern Appalachian forest. Oecologia (Berl) 73 268–71
L Read D Lawrence (2004 (forthcoming)) ArticleTitleRecovery of biomass following shifting cultivation in dry tropical forests of the Yucatan. Ecol Appl . .
InstitutionalAuthorNameSAS Institute (1989) SAS/STAT User’s guide, version 6, volumes 1 and 2. 4th ed SAS Institute Cary (NC) 1789
WL Silver (1994) ArticleTitleIs nutrient availability related to plant nutrient use in humid tropical forests? Oecologia (Berl) 98 336–43
L Singh (1992) ArticleTitleDry matter and nutrient inputs through litter fall in a dry tropical forest of India. Vegatatio 98 129–40
MJ Swift A Russell-Smith TJ Perfect (1981) ArticleTitleDecomposition and mineral–nutrient dynamics of plant litter in a regenerating bush–fallow in sub-humid tropical Nigeria. J Ecol 69 981–95
BL Turner II (1974) ArticleTitlePrehistoric intensive agriculture in the Mayan lowlands. Science 185 118–24
II BL Turner SC Villar D Foster J Geoghegan E Keys P Klepeis D Lawrence PM Mendoza S Manson Y and others Ogneva-Himmelberger (2001) ArticleTitleDeforestation in the Southern Yucatan Peninsular Region: an integrative approach. For Ecol Manag 154 353–70
C Uhl CF Jordan (1984) ArticleTitleSuccession and nutrient dynamics following forest cutting and burning in Amazonia. Ecology 65 1476–90
P Vitousek (1982) ArticleTitleNutrient cycling and nutrient use efficiency. Am Nat 119 553–72
P Vitousek (1984) ArticleTitleLitterfall, nutrient cycling, and nutrient limitation in tropical forests. Ecology 65 285–98
DF Whigham P Zugasty 41Towle E Cabrera Cano J O’Neill E Ley (1990) ArticleTitleThe effect of annual variation in precipitation on growth and litter production in a tropical dry forest in the Yucatan of Mexico. Trop Ecol 31 23–34
Acknowledgements
This work was undertaken through the auspices of the Southern Yucatan Peninsular Region project with core sponsorship from NASA’s LCLUC (Land-cover and Land-use Change) program (NAG 56406) and the Center for Integrated Studies on Global Change, Carnegie Mellon University (CIS-CMU; NSF-SBR 95-21914). Additional funding from NSF, the A. W. Mellon Foundation, and the University of Virginia also supported the specific research in this article. SYPR is a collaborative project of El Colegio de la Frontera Sur (ECOSUR), Harvard University, the George Perkins Marsh Institute of Clark University, the University of Virginia, and Carnegie Mellon University. We are especially grateful to Diego Perez Salicrup, Fausto Bolom-Ton, Juan Mendez Diaz, Jessica Sisco, Heidi Wasson, and Tana Wood for assistance in the field, to Jakara Hubbard for assistance in the lab, and to the farmers of the SYPR who made this work possible.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Read, L., Lawrence, D. Litter Nutrient Dynamics During Succession in Dry Tropical Forests of the Yucatan: Regional and Seasonal Effects . Ecosystems 6, 747–761 (2003). https://doi.org/10.1007/s10021-003-0177-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10021-003-0177-1