
Abstract
The Boundary Waters Canoe Area (BWCA) Wilderness of northern Minnesota, USA, ememplifies how fire management and natural disturbance determine forest composition and landscape structure at a broad scale. Historically, the BWCA (>400,000 ha) was subject to crown fires with a mean rotation period of 50–100 y. Fires often overlapped, creating a mosaic of differently aged stands with many stands burning frequently or, alternatively, escaping fire for several centuries. The BWCA may never have reached a steady-state (defined as a stable landscape age-class structure). In the early 1900s, a diminished fire regime began creating a more demographically diverse forest, characterized by increasingly uneven-aged stands. Shade-tolerant species typical of the region began replacing the shade-intolerant species that composed the fire-generated even-aged stands. Red pine (Pinus resinosa) stands are relatively uncommon in the BWCA today and are of special concern. The replacement of early-to-midsuccessional species is occurring at the scale of individual gaps, producing mixed-species multiaged forests. We used LANDIS, a spatially explicit forest landscape model, to investigate the long-term consequences of fire reintroduction or continuing fire absence on forest composition and landscape structure. Fire reintroduction was evaluated at three potential mean fire rotation periods (FRP): 50,100, and 300 y. Our model scenarios predict that if fire reintroduction mimics the natural fire regime (bracketed by FRP = 50 and 100 y), it will be most successful at preserving the original species composition and landscape structure, although jack pine (Pinus banksiana) may require special management. With limited fire reintroduction, all of the extant species are retained although species dominance and landscape structure will be substantially altered.
If fire remains absent, many fire-dependent species will be lost as local dominants, including red pine. The landscape appears to be in a state of rapid change and a shift in management to promote fire may need to be implemented soon to prevent further deviation from historic, presettlement conditions.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
INTRODUCTION
The Boundary Waters Canoe Area (BWCA) Wilderness of northern Minnesota, USA, lies within the Superior National Forest. The BWCA exemplifies how human policies and natural disturbance operate together at a regional scale to determine long-term forest dynamics. Prior to European settlement, the BWCA (438,000 ha) was subject to crown fires with a mean fire rotation period (FRP, sensu Frelich 2002) of between 50 and 100 y (Heinselman 1973), although this fire regime varied temporally and spatially among land forms. Frequent ground fires also occurred in stands of red pine (Pinus resinosa) and white pine (P. strobus) (Heinselman 1996). Natural fires continued unabated in the BWCA until the early part of the 20th century. Natural fires often overlapped and created a mosaic of relatively young, even-aged stands and older stands that by chance may have escaped fire for centuries. Given this variation, stand ages ranged from less than 10 y to greater than 500 y. Topographic variation and the complex of lakes and wetlands also created refugia that were significantly less likely to burn. The BWCA may have never reached a steady-state (defined as a stable landscape structure) and was never dominated by large contiguous areas of old-growth forest (Heinselman 1973, 1981; Baker 1989). Due to the high fire frequency, the BWCA forests were dominated by even-aged stands of two species that are fire-adapted through opportunistic reproduction: jack pine (P. banksiana), which is serotinous, and aspen (Populus tremuloides). After the reoccurrence of fire, these forest stands were often replaced with the same species (Heinselman 1973; Frelich and Reich 1995).
Since 1910, fire has largely been absent (rotation period >1000 y) as a result of active fire suppression and the absence of large fires that historically burned into the BWCA (Baker 1992; Frelich and Reich 1995). Climatic patterns may have also contributed to the cessation of large, natural fires (Bergeron and others 2001; Johnson and others 2001). The lack of fires has significantly changed tree species demographics, landscape structure, and fuel loading by removing the principal agent of change and renewal. The combined effects of demographics, landscape structure, and fuel loading will determine future forest trajectories and influence forest management options.
Today, much of the forest is transitioning toward a shade-tolerant mixed-age composition. The shade-intolerant, early-to-midsuccessional species that compose the remaining even-aged stands are being replaced by shade-tolerant species. These species include white spruce (Picea glauca), black spruce (Picea mariana), balsam fir (Abies balsamea), and white cedar (Thuja occidentalis). Although elements of the previous even-aged forest can survive in the overstory for long periods of time (Heinselman 1973), seed sources for the colonization of newly disturbed areas may not be adequate. A reduction in pine regeneration was noted soon after fire suppression began (Buell and Niering 1957) and has occurred throughout northern Minnesota (Tester and others 1997). In the past, more frequent and lower-intensity fires (Bergeron and Brisson 1990) also prevented the development of spruce and fir understories in red and white pine stands (Frelich and Reich 1995; Heinselman 1996). Of special management concern, red pine is dependent on fire for adequate regeneration (Bergeron and Brisson 1990) and may be lost as a local dominant within the next 100–200 years (Heinselman 1973; Frelich and Reich 1995).
In addition, the absence of fire has altered landscape structure. Landscape structure is defined here as the spatial arrangement of forest communities (defined by species composition) across the landscape and the spatial arrangement of tree age classes. The replacement of early-or midsuccessional species is occurring at a relatively fine scale as small gaps are created (via wind, disease, and/or insects), producing multiaged forest stands (Frelich and Reich 1995). Future gaps in these mixed-species multiaged stands will likely be filled by the same shade-tolerant species (Frelich and Reich 1995). Without fire, the existing even-aged stands will converge to a multiaged mixed-species forest (Heinselman 1973; Baker 1992). Landscape structure at the broadest scales may be characterized by reduced tree species diversity and the absence of young even-aged forests.
A previous, coarse-grained model of fire suppression in the BWCA concluded that indices of landscape structure will change over time, although not all at the same rate (Baker 1992). Baker (1992) used a model of disturbances and age maps that operated on a grid with 0.5-km × 0.5-km resolution. The model was initialized with a random-age map and individual tree species were not modeled. Patch shape (a patch is an aggregation of pixels with equal age), Shannon diversity, and richness of patch age changed soon after fire was removed from the landscape. Average patch age and shape complexity changed after only several decades and patch size and fine-grained texture changed significantly only after several centuries (Baker 1992).
Finally, the significant reduction in fires may have changed potential fire intensity and severity. Fuels, including fine fuels, coarse woody debris, and “ladder” fuels, have increased over time. If fire is reintroduced, these fuels may initially lead to more intense fires than would have occurred more than 100 years ago (Heinselman 1981; Clark 1988). Flammable understories (that is, ladder fuels) may create optimal conditions for crown fires in red or white pine stands (Heinselman 1996).
Current management policies in Minnesota (Tester and others 1997) and elsewhere in North America are beginning to include the restoration of natural processes, with a particular focus on fire (Moore and others 1999; Malakoff 2002). The concept of restoring or mimicking the historic or natural range of variability has been advocated as a management tool that would promote ecosystem health and ecological restoration (Landres 1999). However, reintroduction of the range of natural variability of fire will not necessarily return the system to its historic state. Because tree age classes are older, fuels have increased, and landscape configuration has been altered due to the absence of fire, the reintroduction of fire may create novel landscapes and potentially have unanticipated negative consequences. For example, crown fires may be desirable in senescing even-aged jack pine forests, but they are undesirable in rare old-growth stands of red or white pine.
Our objective is to explore the long-term consequences of continued fire absence or fire reintroduction, including the reintroduction of the historic range of fire variability, on forest composition and landscape structure in the BWCA. Ultimately, our goal is to estimate long-term consequences of management policies and enable better management decisions. Therefore, we simulated four different fire management scenarios and examined whether species composition, age structure, and landscape structure differed among the predicted landscapes. As our focus is on the long-term ecological consequences of fire reintroduction versus continued fire absence, the effects of the July 4, 1999, blowdown were not addressed. The effects of insects and disease, which can be important in these systems, are not consider here .
We used a spatially explcit interactive forest landscape model to simulate both continued fire absence and changes in the current policy that would allow or reestablish varying degrees of the natural fire regime. The purpose of the landscape model is to explain the consequences of a given set of assumptions and fire rotation periods. Our fire model is nonmechanistic and does not include climatic effects and therefore does not predict fire frequency. Rather, a predetermined fire rotation period, representing different management options, is imposed upon the landscape. Because we did not incorporate weather effects (compared with Rothermal 1972; Bessie and Johnson 1995), we assumed that fuel accumulation, cohort age, and species fire susceptibility determined the effects of fire reintroduction.
Our analysis is unique in that we initialized our simulations with actual stand ages and dominant and codominant species drawn from the most detailed possible data for the year 1995. By using actual, versus random, stand age data, we incorporate landscape history and the legacy effects of past disturbances. The use of empirical forest composition data allowed us to simulate the ecological effects of individual species and life histories on landscape dynamics.
METHODS
Study Area
The BWCA is a protected wilderness area within the Superior National Forest, Minnesota, in the north central US. The 438,000-ha tract is no longer modified directly by humans and contains almost all of the presettlement flora and fauna native to the area (Heinselman 1973). BWCA forests are transitional between boreal forests and Great Lakes north temperate forests and are dominated by red pine, jack pine, white pine, black spruce, white spruce, balsam fir, white cedar, quaking aspen, and tamarack (Larix laricina) (Heinselman 1973; Baker 1989). Red maple (Acer rubrum), black ash (Fraxinus nigra), bigtooth aspen (Populus grandidentata), paper birch (Betula papyrifera), and balsam poplar (P. balsamifera) are also present (Heinselman 1973).
The climate is cold temperate continental and the glacial geology is typical of the Laurentian continental shield (Heinselman 1973). Soils are generally thin due to glacial scouring, although some till, outwash, and lacustrine deposits exist. Soil type and depth to bedrock are highly heterogeneous at fine spatial scales (Heinselman 1973).
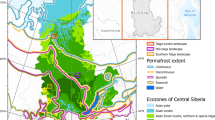
Our research area included a portion (195,000 ha) of the total BWCA (Figure 1). This area is dominated by the Gabbro Lake Bedrock Complex, the largest Land Type Association (LTA) within the BWCA (Cleland and others 1997). Small portions of neighboring LTAs to the east, south, and west are also included. By focusing on only a few large contiguous land types, we minimized variation in the physical environment and decreased the effect of landscape borders.

Study area within the Boundary Waters Canoe Area Wilderness, Minnesota, USA.
With the exception of prescribed burning in response to the July 4, 1999, blowdown, the current fire regime is limited to relatively small natural fires and the current management policy is effectively equivalent to total suppression (> 1000 y rotation period). Clearcut logging occurred up to the late 1970s in our study area. Together approximately 40% of the study area has been disturbed in the past 100 years (Mark White, University of Minnesota-Duluth, personal communication), not including the 1999 blowdown. The 1999 blowdown partially damaged or destroyed approximately 30% of BWCA forests (Mark White personal communication) and approximately 5% of our study area, concentrated in the north.
Landscape Simulation Model
A forest landscape simulation model, LANDIS, was used to assess how fire reintroduction or suppression will affect the species composition and landscape structure of the BWCA. In addition to modeling and testing the effects of fire, LANDIS simulates succession and other spatially interactive processes, including blowdowns and seed dispersal (Mladenoff and others 1996; Mladenoff and He 1999). LANDIS uses a 10-y time-step and was designed to model broad-scale spatially interactive processes over long time periods. LANDIS operates on a raster map that is divided into cells; cell size can have grain sizes from 10 m × 10 m to 1 km × 1 km. Each cell is assigned to an ecoregion or land type that is assumed to have homogeneous soils and climate. All modeled processes occur within a spatial context, including explicit location, extent, and/or spread of processes, as necessary.
Within LANDIS, disturbances occur stochastically but their rotation period can be set to represent any number of historic or potential scenarios. The LANDIS fire module was designed to approximate fire initiation, spread, and severity given a limited number of state variables and input parameters (He and Mladenoff 1999b) that do not include weather effects such as wind speed, humidity, and fuel moisture (Rothermal 1972; Finney 1998). The location of fire ignition events (as would occur from lightening strikes) is random, and the probability of subsequent fire initiation is determined by the designated fire rotation period and stand age (He and Mladenoff 1999b). Simulated fire spread is limited by the fuel accumulation of neighboring cells and a designated maximum fire size (He and Mladenoff 1999b). Fuel accumulation increases logarithmically, not linearly, with time since the last fire and determines a fire severity class that varies from 1 (lower potential mortality) to 5 (complete mortality). More frequent fires (lower FRP) will produce a lower mean fire severity class, averaged across the entire landscape. Less frequent fires will increase the mean fire severity class.
Actual fire severity (the mortality caused by a fire) is a function of the fire severity class, cohort ages, and species susceptibility; young cohorts are most vulnerable to fire mortality (He and Mladenoff 1999b). A fire tolerance class (1–5) is assigned to each species; if the fire severity class exceeds a species tolerance class, all cohorts of that species will be killed. The same fire severity class may have different ecological consequences, dependent upon the forest community and the age of the existing cohorts. For example, a short period of fuel accumulation (low fire severity class) would cause complete mortality in a young jack pine stand and little mortality in a stand dominated by old (> 100 y) red pines. Although our model does not simulate fire intensity per se, any fire that causes complete mortality in a conifer stand is effectively a crown fire.
LANDIS has previously been validated for both internal logic and ecological representation, and sensitivity analyses of key parameters have been performed (He and others 1998; He and Mladenoff 1999a, 1999b; Mladenoff and He 1999). Nevertheless, the constraints of our input data and the stochastic nature of the model restrict analysis and conclusions to the general patterns created at the landscape scale; predictions of actual events and communities at specific locations are not possible and strict validation is not possible when modeling future potential scenarios (Urban and others 1999).
Input Data
LANDIS was initialized with estimates of the composition, stand ages, climate, and soils of the landscape in the year 1995. The data were compiled and analyzed at a resolution of 28.5 m × 28.5 m, the same resolution as the available satellite vegetation classification data (Wolter and White 2002), for a total of approximately 1.8 million pixels (Figure 2). Using the maximum resolution available allowed us to examine the effects of lakes and wetlands and the fate of small forest stands of less common species (for example, red pine). Although lakes and wetlands likely only reduce the probability of fire spread, we assumed that they would completely stop fire spread. Lakes and wetlands are also important as they will limit seed dispersal of upland tree species.

Percent cover estimated from satellite vegetation (1995) classification with a 28.5 × 28.5-m resolution (Wolter and White 2002). Mixed classes were grouped with their respective dominants for map display. CO indicates unclassified conifer.
Initial Forest Composition and Age Input Map
Multiple steps were required to generate the necessary initial site composition and age map. First, a map of stand ages was compiled from Heinselman’s (1973) stand age delineations to which more recent logging data were added (Mark White, University of Minnesota-Duluth, personal communication). Stand age changes due to the 1999 blowdown were not included. This stand age map was combined with a satellite vegetation classification of canopy dominant species (Wolter and White 2002) in Arc/Info (ESRI 2001). The combination resulted in more than 256 unique species–age combinations, exceeding the maximum allowed by LANDIS. Therefore, these classes were reduced to about 200 by combining the classes with the least number of pixels into the nearest age class with the same vegetation classification.
An estimate of codominant tree species (species and age) is also required to initialize LANDIS. Unfortunately, U.S. Forest Service Forest Inventory and Analysis data (Hansen and others 1992; for example, He and others 1998) or other spatially extensive community data are not available for the BWCA. Therefore, we used forest composition data from Ohmann and Ream’s (1971) BWCA stand descriptions. Ohmann and Ream (1971) quantified eight typical community types (defined by canopy dominance) with associated subcanopy or codominant species and respective basal areas. Each of the satellite vegetation classes (a) was assigned to one of the Ohmann community types or (b) was left as a single species community as defined by Wolter and White (2002), dependent on whether a pixel was classified as a mixed class or as a single-species class (Wolter and White 2002). Undoubtedly, many of the single-species pixels from Wolter and White (2002) contained understory species, but it could not be determined whether or which additional species may be present.
Finally, we incorporated the codominants as defined by Ohmann and Ream (1971) with the canopy dominants as determined by Wolter and White (2002). Within LANDIS, species are represented as the presence of 10-year age cohorts in a cell; individual tree or other quantitative data are not estimated. Species dominance in a cell is approximated by species importance value (IV), defined as
where A is the age of the oldest cohort, L is longevity of the species, and RC is the reclass coefficient. The reclass coefficient ranges from zero to one and weights importance values by relative species height and diameter (that is, “stature”). The species with the highest IV is the dominant. To preserve the codominant species composition and structure (species and relative basal areas) measured by Ohmann and Ream (1971), we set the basal area (BA) proportions they measured equal to LANDIS importance value proportions, such that for each community:
where BACODOM and BADOM are basal areas from codominants and dominants, respectively, drawn from Ohmann and Ream (1971), and IVCODOM and IVDOM are LANDIS importance values. Because we know the dominant species and age from our combination of the satellite vegetation data and stand age data, maximum ages for each codominant species were calculated as:
A codominant cohort was then assumed to be present at that age; younger cohorts of the same species were not estimated. If the calculated codominant age (ACODOM) was less than 10 years or greater than the life span of that species (LCODOM), it was not added to the codominant structure for that community type.
If a mixed-species community was relatively young and the codominant basal area (BACODOM) was small relative to the basal area of the dominant (BADOM), the site contained only the dominant species. As forest age increased, codominants began to appear as young cohorts. Codominant age increased as the age of the dominant species increased. A very short-lived codominant in a relatively old forest site may have had an age calculated to be greater than its longevity and was excluded. The original site age was not always preserved. In rare cases, the age of a codominant exceeded the age of the dominant, although this result did not change either the importance values or the dominant species. This method preserved the dominant species, as provided by Wolter and White (2002), the stand ages provided by Heinselman (1973), and provided realistic estimates of codominant species and ages. The method is appropriate when only species age class presence is being tracked over time.
Calculation of Species Establishment Probabilities
LANDIS also requires a species establishment coefficient (SEC), the probability of each species establishment in each ecoregion or land type (He and others 1998). We generated SECs using the LINKAGES model (Pastor and Post 1986) and a modification of a previously developed procedure (He and others 1998). LINKAGES is an ecosystem process model that simulates individual tree establishment, growth, competition, and mort-ality as a function of soil water, nutrient dynamics, and monthly average temperature and precipitation (Pastor and Post 1986). The method for calculating SECs presented in He and others (1998) was modified as follows: For each species, LINKAGES was initiated with a stand of 200 trees of that species, 1 year old and 1 cm in diameter at 1.4 m, thus simulating seedling growth and survival in open-grown conditions. LINKAGES was altered to output a probability of establishment and growth (or SEC) based on the change in biomass between years 0 and 10. For each LINKAGES replication, if the stand exhibited positive biomass growth during the first 10 years, the replication represented a successful establishment and growth to age 10; 100 replications were used for each species. The SEC was calculated as the number of successful establishments (replications) divided by the total number of replications. In this way, a mean probability of establishment was generated for each species and for each ecoregion, that is, LINKAGES predictions were extrapolated to the entire ecoregion by expected value (King 1991).
Required input data to LINKAGES include soil carbon (Mg/ha), soil nitrogen (Mg/ha), soil water field capacity (cm), wilting point (cm), and mon-thly climate data (mean temperature, mean precipitation, standard deviation temperature, and standard deviation precipitation). In addition to mean soil carbon and soil nitrogen, LINKAGES was altered to accept the variability of initial soil carbon (soil C standard deviation) and initial soil nitrogen (soil N standard deviation) within an ecoregion. Variability of soil carbon and nitrogen was added to represent the range of soil conditions across an ecoregion at a coarse spatial resolution. Mean soil carbon, soil carbon standard deviation, field capacity, and wilting point were derived from the STATSGO database (Cosby and others 1984; Saxton and others 1986; Davidson and Lefebvre 1993; STATSGO 1994; Davidson 1995). Mean soil nitrogen and standard deviation were estimated from Host and Pastor (1998). At the beginning of each replication, base soil carbon and nitrogen were estimated from a mean for the ecoregion plus or minus an amount determined by the product of the standard deviation and a random normal number generator. Climate data were derived from ZedX GIS coverages (ZedX 1995).
Species establishment coefficients were calculated for two ecoregions, both derived from STATSGO polygons (Table 1 and Figure 1). A third ecoregion was created from the satellite vegetation classification (Wolter and White 2002) and includes only areas designated as pure black spruce forests. We assumed that only black spruce could establish in these lowlands. However, due to the wide dispersal of this ecoregion across the study area, these black spruce bogs could potentially serve as important seed sources for the establishment of black spruce on adjoining upland areas.
Model Run Scenarios
Model scenarios varied only by mean fire rotation period (FRP), averaged over the entire study area and all years. The following FRPs were used to test how fire absence and increasing FRP will affect the forest composition and structure of the BWCA: no fire, and means of 50 y, 100 y, 300 y. FRPs equal to means of 50 and 100 y bracket the historic fire regime (Heinselman 1973) or “range of natural variability” (Landres and others 1999; Tinker and Knight 2001). A FRP of 300y represents a management policy of moderate fire reintroduction. Each scenario was run for 300 y beginning from the year 1995. Fire size follows a lognormal distribution (He and Mladenoff 1999b) with a mean fire size of 7500 ha (Heinselman 1973). Harvesting was set to zero and wind throw disturbance was maintained at current levels (rotation period ∼ 500 y) for all scenarios.
Each scenario was replicated four times with different random number seeds. Due to the intensive computing time and space required for each model run (>1 Gbyte), more replicates were not possible. However, given the number of individual cells (>1.8 million) and the length of the simulation, we believe more replicates were not necessary to estimate the effect of random events or stochastic variation. These replicates were not meant as a Monte Carlo simulation but rather as a means to assess within-scenario variation; they were not used to test for significant differences.
Landscape Structure
One objective was to determine whether, and to what degree, the BWCA landscape changed in regard to stand age structure (that is, even-aged or uneven-aged) and species structure (that is, mono-dominant or mixed-species). Texture was chosen as the most representative landscape metric for the analysis of landscape structure because it incorporates information from neighboring cells when determining the age or species structure for any given cell. Texture analysis using the eight neighboring cells calculates the texture of a square block containing 9 cells (Musick and Grover 1991; Plotnick and others 1993). To assess scale variability, texture was analyzed at three scales (resolutions): 0.08 ha, 8.1 ha, and 812 ha. The smallest scale represents individual cells. The two larger scales were generated by aggregating individual cells, 10 × 10 and 100 × 100, respectively. Texture was analyzed using APACK, freely available landscape pattern analysis software (Mladenoff and DeZonia 2001).
Stand age structure was calculated using the inverse difference moment, a measure of the mean difference between the age of a cell (the age of the oldest cohort) and the age of its eight neighbors (Musick and Grover 1991). A value of 1 indicates that all adjacent neighbors have the same age as the current site (completely even-aged), and a value of 0 indicates that all neighbors have different ages (an uneven-aged forest).
Species structure was calculated from output maps of the dominant species or community at the end of each simulation; 8 classifications were used: jack pine, aspen and birch, black spruce, red pine, white spruce and fir, white pine, White Cedar, and other. Because maps of dominant species are composed of nominal data, the angular second moment was used to measure texture (Musick and Grover 1991). Similar to the inverse difference moment, the angular second moment ranges from 0 (highly disaggregated) to 1 (a single cover type for all nine cells) (Musick and Grover 1991).
RESULTS
Simulated fires vary spatially (Figure 3). Areas in large contiguous stands burned more frequently than edges, islands, and areas dominated by lakes and wetlands (Figure 3a and b). Notice that when the mean FRP was 50 or 100 y, only isolated forest stands escaped burning. When fire was infrequent (FRP = 100y), 26% of the landscape never burned and less than 5% of the landscape burned more than twice over the length of the simulation (Figure 3c). Spatial variation of fire occurence created a mosaic of mean fire frequencies that ranged from 33–300 y, 42–300 y, and 100–300 y (excluding areas that did not burn), for mean FRP = 50, 100, and 300 y, respectively (Figure 3).

Burn frequency (number of fires per century) for three scenarios: (a) fire rotation period (FRP) = 50 y, b) FRP = 100 y, c) FRP = 300 y. Data are drawn from one scenario out of four total.
Although the location of each potential fire ignition was random, fuel accumulation affected the probability of fire initiation, spread, and fire severity class (Figure 4). Although the fire severity class was positively correlated with mean FRP as expected (Figure 4a, b, and c ), we cannot assume that the fire regime was dominated by low-mortality (for example, surface) fires. Both of the frequent-fire scenarios (FRP 50 and 100) were dominated by fire-intolerant species (Figure 5a and b ). Aspen stands (fire tolerance = 2) would have experienced complete mortality in most fires when FRP was equal to 50y and the fire severity class was higher than 2. Jack pine stands (fire tolerance = 3) would have experienced complete mortality when the fire severity class was higher than 3 (FRP = 100y). When FRP was equal to 300y, all species would have experienced complete mortality, although there were fewer total fires.

Mean annual percentage of landscape burned (bars; N = 4) and mean fire severity (line; N = 4; 1 = low potential mortality and 5 = complete mortality) over 300 simulation years. Three scenarios are presented: (a) mean fire rotation period (FRP) = 50 y; (b) FRP = 100 y; (c) FRP = 300 y.

Dominant vegetation change over 300 simulation years and four model scenarios: (a) fire rotation period (FRP) = 50 y, (b) FRP = 100 y, (c) FRP = 300 y, (d) no fires simulated. Species not depicted in any given graph are summarized with “Other.” Error bars represent ± 1 standard error (N = 4).
Early-successional species, including aspen, birch, and jack pine, dominated scenarios with very frequent fires (FRP = 50y) and frequent fires (FRP = 100y; Figure 5). The greatest landscape evenness occurred when mean FRP equaled 300 y (Figure 5). White spruce, balsam fir, and white pine dominated when fire was excluded. In all scenarios, the areas occupied by black spruce remained fairly constant and did not expand significantly beyond the current black spruce lowlands.
There are multiple potential causes of variability in cell age (Figure 6), including area burned, fire severity, and cohort senescence. When fires were more frequent and less severe (Figure 4a and b ), few cohorts survived and few cells advanced to a greater age (Figure 6a and b ), indicating that these low-severity fires caused high mortality. When fires were infrequent (mean FRP = 300y), more cells survived to beyond 100 years of age (Figure 6c). Without fire, cohort senescence became the only source of variability in cell age and species longevity determined age. The low-fire-probability scenario had a bimodal age distribution at the end of the simulation (Figure 6d). The cells less than 160 years of age correspond to recruitment of late-successional species following the senescence of jack pine and aspen between years 100 and 150 (Figure 4d). Cell ages greater than 250 represent old cohorts of white pine (Figure 7).

Maximum cohort age at final year (300) for four scenarios: (a) fire rotation period (FRP) = 50 y, (b) FRP = 100 y, (c) FRP = 300 y, (d) no fires simulated. The data represent the mean of 4 replications (N = 4).

Demographics for three fire-dependent species: red pine (Pinus resinosa), white pine (Pinus strobus), and jack pine (Pinus banksiana), and four model scenarios: FRP = 50,100, and 300 y and no fires, respectively. The data represent the mean of 4 replications (N = 4).
Change over Time
The temporal change of the landscape proportion dominated landscape dominance: by different species or functional groups depicts the transition from current day conditions to the endpoint conditions (Figure 5). Because species dominance is a function of cohort age (Eq. 1), the temporal analysis of landscape dominance largely reflects the oldest cohorts of a species or functional group. In all of the fire reintroduction scenarios, jack pine dominance initially declined due to the combined effect of both fire severity (Figure 4) and the advanced age of the existing cohorts. When FRP was 50 y, jack pine dominance recovered within the first 100 years as fire severity quickly declined (Figure 4a). When FRP was 100 y, jack pine recovery was much longer due to the periodic occurrence of relatively severe fires (Figure 4b). These fires destroyed the oldest jack pine cohorts although they provided suitable conditions for reproduction. In both FRP equal to 50-y and 100-y scenarios, red pine increased significantly over time, white spruce and fir declined, and other species remained relatively constant. Both red and white pine had better regeneration than jack pine when FRP was 50 y (Figure 7). However, landscape dominance is a function of cohort age relative to longevity (Eq. 1); therefore, red and white pine dominance is lower than jack pine dominance when FRP was equal to 50 years (Figure 5).
With a 300-y fire rotation period Jack pine declined throughout the simulation (Figure 5c). Simultaneously, the landscape became increasingly dominated by red pine, white pine, white spruce, and fir. Red pine and white pine regeneration was reduced compared with FRP of 100 y, although more cohorts survived to greater age (Figure 7h).
When fire was entirely excluded, jack pine, aspen/birch, and red pine were eventually entirely lost as canopy dominants (Figure 5d). White spruce, fir, and white pine increased in landscape dominance. At year 300, white pine was not regenerating and these white pine cohorts were nearing the end of their lifespan (Figure 7h).
Landscape Structure
Landscape age structure exhibited a notable discontinuity among scales (Figure 8a). At the two finest resolutions (0.08 and 8.1 ha), age texture followed a predictable pattern, whereas texture is greatest for FRP of 50 and 100 y and least for fire exclusion, that is, the landscape was more even-aged when disturbance was frequent or very frequent (FRP = 50 and 100 y). At the coarsest resolution tested, 812 ha, age texture was more variable over time, FRP equal to 50 y was the least even-aged, and fire exclusion was the most even-aged at the end of the simulation. There was greater temporal variability at large scales when fires were excluded.

Landscape structure of simulated BWCA forests: (a) inverse difference moment of site ages and (b) angular second moment of dominant species. Change over 300 simulation years and four model scenarios represented: mean fire rotation period (FRP) = 50 y (dotted line), FRP = 100 y (solid line), FRP = 300 y (dashed line), and no fires (dot–dash line). The inverse difference moment was calculated using an eight-neighborhood rule. Error bars represent ± 1 standard error (N = 4).
The angular second moment of dominant species reflects age texture to a degree but lacks the strong discontinuity among scales. At all scales, the historic fire regimes (FRP = 50 and 100 y) had the highest angular second moments (Figure 8b) during the first 250 years of the simulation, indicating that species were arranged in large contiguous patches. After year 150, species aggregation in the fire exclusion scenario increased and became greater than all other scenarios at year 300. This corresponded with the loss of species diversity and concomitant increase in white spruce/fir abundance in the last 100 years of the fire exclusion scenario (Figure 5d). Temporal variation of species texture consistently increased with scale for all scenarios (Figure 8b).
DISCUSSION
The shifts in species composition over time and among scenarios enables us to estimate the potential consequences of continued fire absence or fire reintroduction on both species composition and landscape structure. Our results confirm prior estimations of projected change and provide a range of possible projections. Frelich and Reich (1995) found that a change in successional pathways had already begun as a result of fire absence. Our model indicates that this trend will clearly continue if fire absence continues, with significant changes in species composition occurring in the immediate future and the landscape will increasingly shift toward white spruce and balsam fir. Contrary to Frelich and Reich (1995), we predict a constant black spruce component, reflecting the relatively low species establishment coefficient relative to white spruce. Our species establishment coefficients were calculated assuming full sunlight. This may underestimate the ability of black spruce to establish under moderate shade conditions that have higher soil moisture. Regardless of which spruce or fir species ultimately becomes dominant, a landscape highly vulnerable to insect outbreaks will be created (Heinselman 1973). That such a system would be stable is unlikely (Heinselman 1973; Holling 1988) and will be addressed in ongoing model refinements.
Our three fire reintroduction scenarios indicate that even a modest fire reintroduction regime can conserve all of the species of the current landscape, at least at low numbers. Red pine, a species of particular concern, successfully regenerates under all fire reintroduction scenarios. However, survival to ages beyond 150 years is limited under the very frequent (FRP = 50 y) fire reintroduction scenarios. Today, old-growth stands of red pine (>150 years of age) are most often found on rocky outcrops, on the north-facing slopes of hillsides, and on islands, all of which serve as refugia from crown fires (Heinselman 1996). We did not incorporate topographic variation and these geographic features often exist at scales finer than our model resolution. The complex geography of lakes and wetlands do, however, create some refugia. Nevertheless, we likely underestimate the ability of isolated stands to survive to advanced age.
Under all but the most frequent fire reintroduction scenario (FRP = 50 y), dominance by jack pine, arguably the most fire-dependent species, is substantially reduced in area over time. When a very frequent fire regime is reintroduced (FRP = 50 y), jack pine dominance is generally maintained. The scenario of frequent burning (FRP = 100 y) demonstrates the sensitivity of jack pine to fire severity. Because fire severity is higher, this scenario favors an initial expansion of early-successional aspen and birch, both of which can disperse seeds much greater distances than jack pine. Jack pine does, however, increase in dominance over time. Each fire has unique consequences for jack pine regeneration. If a fire burns twice over an area before jack pine has reached reproductive age, jack pine will be locally extirpated (Frelich 2002). Older cohorts may survive, dependent upon the fire severity class. Nevertheless, we can deduce a larger pattern without reconstructing the history of every simulated fire: Increased fire frequency provides opportunity for regeneration and the reduced fire severity allows more cohorts to reach maturity (Figure 7j). Although jack pine still exists when FRP reachs 300 y, regeneration is low and infrequent, indicating an eventual loss.
Both higher-frequency (FRP = 50 and 100 y) fire reintroduction scenarios, which encompass the historic range of variability, predict a loss of white cedar in the landscape. Although a reduction in white cedar was anticipated if fire is reintroduced, current community data indicate that, like red pine, cedar is able to survive and reproduce in isolated refugia that escape fire, such as islands, low depressions, and along lake shores. Again, our predictions probably underestimate the ability of white cedar to persist on the landscape.
Without fire reintroduction, our prediction of landscape structure follows the successional and demographic transition to an older, more uneven-aged, mixed-species landscape that is already occurring in the BWCA (Frelich and Reich 1995). As noted previously (Baker 1989; Frelich and Reich 1995), these results are highly scale dependent. At the finer scales, the absence of fire creates the most heterogeneous age structure. He and Mladenoff (1999b) also found that when succession dominated age variation, the landscape became disaggregated (increased fine-grain heterogeneity) at the finest scale. At the coarsest scale considered (812 ha), the fire absence scenario emerges as the most even-aged and monodominant landscape. This pattern reflects the synchronicity of cell ages (Figure 6d) and the loss of species diversity (Figure 5d), respectively. At the end of the simulations, large-scale age structure also exhibits a correlation between heterogeneity and fire introduction (FRP = 300 > FRP = 100 > FRP = 50y). These results confirm the patterns elucidated by He and Mladenoff (1999b) where more severe fires created coars-grain heterogeneity.
If we define steady-state as the temporal stability of age structure (Romme 1982; Baker 1989) and (or) the dominant species structure, our results indicate that at the coarsest scale measured, none of the scenarios clearly exhibits steady-state behavior (Figure 8a and b ); all of our scenarios exhibit large variation in landscape structure. As our landscape contained limited environmental variation, with only three broad land types, steady-state conditions are not precluded a priori (Baker 1992). Both Heinselman (1973) and Baker (1989) concluded that the BWCA was never in a stable state, although we cannot know which of our scenarios, if any, best approximates these historic conditions. In this respect, the BWCA is somewhat similar to western North American conifer ecosystems (Romme 1982; Turner and Romme 1994). At all scales, the fire absence scenario exhibits the least stead-state behavior. This relatively high temporal variation indicates that a transition to a “steady-state mosaic” (Bormann and Likens 1979), which was not historically common in the BWCA (Frelich and Reich 1995), would not develop within the time frame we examined.
Scenario Limitations
Our scenarios are limited by both the available input data and the model design goals. As mentioned earlier, the absence of topographic variation in the input data and a lack of high-resolution soils data limit the amount of among-site variability in both fire spread and species establishment coefficients. Likewise, our initial stand ages and forest composition data are limited by the available data. In addition to input data limitations, the model does have some inherent design limitations.
Red pine offers a good example of the input data and model design limitations. Without fire reintroduction, red pine regeneration falls to zero, although in reality we would expect some scattered but poor regeneration in isolated refugia. When the conditions are optimal for red pine regeneration (FRP = 100 y), our results depict the formation of large contiguous stands of red pine. Red pine is a weak seeder (Bergeron and Brisson 1990) and regeneration is a function of postfire soil conditions (Ohmann and Grigal 1957; Heinselman 1981). We were unable to simulate variable soil conditions because of the coarse resolution of STATSGO data and LANDIS does not simulate species-specific differences in seed volume or quantity.
Another model limitation is a lack of variability in fire severity due to species flammability and weather. Historically, areas dominated by jack pine may have burned at a greater frequency than the surrounding landscape (Heinselman 1973; Baker 1993). Model refinements that incorporate species-specific flammability are currently under development. Very extreme drought is often as important, if not more important, than fuel for determining fire severity (Bessie and Johnson 1995, Johnson and others 1998, 2001; Frelich 2002). However, this variability is not currently included in our model and we may be underestimating long-term average fire severity for some scenarios. At the same time, not every forest type will sustain crown fires and our infrequent-fire scenario (FRP = 300 y) may overestimate average fire severity. Despite these limitations, we were able to predict and analyze the broad-scale trends, and we do not believe these limitations detract from our conclusions about these general trends.
Management Implications
Our research provides insights that will be valuable when considering future management options for the BWCA. First, both species composition and age structure have begun to change from single speciesdominated even-aged forests, toward a mixed species late-successional forest. Our results indicate that this change will accelerate in the next 100 years. Although a return to presettlement fire conditions (FRP between 50 and 100 y) would best serve to retain the original character of the BWCA, the reintroduction of a reduced-fire regime will preserve all of the existing elements. Furthermore, fire reintroduction need not be constrained to avoid high-severity fires, including crown fires. High-severity fires would not necessarily lead to shifts in landscape structure beyond what has existed in the past 500 years (Baker 1992) and our results suggest that the risk of species loss is minimal. On the contrary, some large fires are likely necessary to sufficiently reset succession and allow pine regeneration, although we did not explicitly test this hypothesis.
A simplistic reintroduction of the historic range of variability may not provide complete landscape restoration. As Landres (1999) noted, future systems are a function of current states. A direct linear return to prior states may not be possible. Our model indicates that a mean fire regime equivalent to the most frequent historic fire regime (mean FRP = 50 y, range = 33–300 y) is necessary to maintain landscape dominance of jack pine and the historic character of the landscape. Even with fire reintroduction, many jack pine stands will senesce and be replaced by late-successional species before fire creates an opportunity for reproduction. Frelich and Reich (1995) reached similar conclusions based on field studies. Maintaining the original landscape proportion of jack pine may require additional species-specific management practices. Reintroduced fire must be spatially heterogeneous (with spatially variable frequency) so as to preserve the extensive jack pine forests. For red pine and white cedar management, a spatially heterogeneous approach to fire will be effective if isolated refugia are allowed to escape the most intense fires.
References
WL Baker (1989) ArticleTitleLandscape ecology and nature reserve design in the Boundary Waters Canoe Area, Minnesota Ecology 70 23–35
WL Baker (1992) ArticleTitleEffects of settlement and fire suppression on landscape structure Ecology 73 1879–87
WL Baker (1993) ArticleTitleSpatially heterogeneous multi-scale response of landscapes to fire suppression Oikos 66 66–71
Y Bergeron J Brisson (1990) ArticleTitleFire regime in red pine stand at the northern limit of the species’ range Ecology 71 1352–64
Y Bergeron S Gauthier V Kafka P Lefort D Lesieur (2001) ArticleTitleNatural fire frequency for the eastern Canadian boreal forest: consequences for sustainable forestry Can J For Res 31 384–91 Occurrence Handle10.1139/cjfr-31-3-384
WC Bessie EA Johnson (1995) ArticleTitleThe relative importance of fuels and weather on fire behavior in subalpine forests Ecology 76 747–62
MF Buell WA Niering (1957) ArticleTitleFir–spruce–birch forest in northern Minnesota Ecology 38 602–10
JS Clark (1988) ArticleTitleEffect of climate change on fire regimes in northwestern Minnesota Nature 334 233–5 Occurrence Handle10.1038/334233a0
DT Cleland PE Avers WH McNab ME Jensen RG Bailey T King WE Russell (1997) National hierarchical framework of ecological units MS Boyce A Haney (Eds) Ecosystem Management: Applications for sustainable forest and wildlife resources Yale University Press New Haven, CT 181– 200
BJ Cosby GM Hornberger RB Clapp TR Ginn (1984) ArticleTitleA statistical exploration of the relationships of soil moisture characteristics to the physical properties of soils Water Resources Res 20 682–90
EA Davidson (1995) ArticleTitleSpatial covariation of soil organic carbon, clay content, and drainage class at a regional scale Landscape Ecol 10 349–62 Occurrence Handle10.1007/BF00130212
EA Davidson PA Lefebvre (1993) ArticleTitleEstimating regional carbon stocks and spatially covarying edaphic factors using soil maps at three scales Biogeochemistry 22 107–31
InstitutionalAuthorNameEnvironmental Systems Research Institute (ESRI), Inc (2001) ArcDoc Version 8.1.2 ESRI, Inc Redlands, CA
Finney MA. 1998. Farsite: Fire Area Simulator—Model Development and Evaluation. Research Paper RMRS-RP-4 Revised. USDA Forest Service Intermountain Fire Sciences Laboratory. Missoula, MT
LE Frelich (2002) Forest dynamics and disturbance regimes: studies from temperate evergreen-deciduous forests Cambridge University Press Cambridge, UK
LE Frelich PB Reich (1995) ArticleTitleSpatial patterns and succession in a Minnesota southern-boreal forest Ecol Monogr 65 325–46
Hansen MH, Frieswyk T, Glover JF, Kelly JF 1992. The eastwide forest inventory data base: Users manual. GTR NC-151. St. Paul, MN: USDA Forest Service North Central Forest Experiment Station
HS He DJ Mladenoff (1999a) ArticleTitleThe effects of seed dispersal on the simulation of long-term forest landscape change Ecosystems 2 308–19 Occurrence Handle10.1007/s100219900082
HS He DJ Mladenoff (1999b) ArticleTitleSpatially explicit and stochastic simulation of forest landscape fire disturbance and succession Ecology 80 81–99
HS He DJ Mladenoff TR Crow (1998) ArticleTitleLinking an ecosystem model and a landscape model to study forest species response to climate warming Ecol Model 114 213–33 Occurrence Handle10.1016/S0304-3800(98)00147-1
ML Heinselman (1973) ArticleTitleFire in the virgin forests of the boundary waters canoe area Minnesota, Quatern Res 3 329–82
ML Heinselmann (1981) Fire and succession in the conifer forests of northern North America DC West HH Shugart DB Botkin (Eds) Forest succession: concepts and application Springer-Verlag New York 374–405
ML Heinselman (1996) The Boundary Waters Wilderness ecosystem Minneapolis, MN University of Minnesota Press
CS Holling (1988) ArticleTitleTemperate forest insect outbreaks, tropical deforestation and migratory birds Mem Entomol Soc Can 146 21–32
Host G, Pastor J. 1998. Modeling forest succession among ecological land units in northern Minnesota. Conserv Ecol [online] 2
EA Johnson K Miyanishi MHJ Weir (1998) ArticleTitleWildfires in the western Canadian boreal forest: Landscape patterns and ecosystem management J Veget Sci 9 603–10
EA Johnson K Miyanishi SRJ Bridge (2001) ArticleTitleWildfire regime in the boreal forests and the idea of suppression and fuel buildup Conserv Biol 15 1554–7 Occurrence Handle10.1046/j.1523-1739.2001.01005.x
AW King (1991) Translating models across scales in the landscape MG Turner RH Gardner (Eds) Quantitative methods in landscape ecology Springer-Verlag New York 479–517
PB Landres P Morgan FJ Swanson (1999) ArticleTitleOverview of the use of natural variability concepts in managing ecological systems Ecol Appl 9 1179–88
D Malakoff (2002) ArticleTitleArizona ecologist puts stamp on forest restoration debate Science 297 2194–6 Occurrence Handle10.1126/science.297.5590.2194 Occurrence Handle12351767
Mladenoff DJ, Dezonia B. 2001. APACK. User’s Guide and Application, available at: http://Flel.Forest.Wisc.Edu/Projects/APACK/. Madison, WI: University of Wisconsin–Madison
MM Moore WW Covington PZ Fule (1999) ArticleTitleReference conditions and ecological restoration; a southwestern Ponderosa pine perspective Ecol Appl 9 1266–77
HB Musick HD Grover (1991) Image textural measures as indices of landscape pattern MG Turner RH Gardner (Eds) Quantitative Methods in Landscape Ecology Springer-Verlag New York 77–104
LF Ohmann DF Grigal (1981) ArticleTitleContrasting vegetation responses following two forest fires in northeastern Minnesota Am Midland Nat 106 54–64
LF Ohmann RR Ream (1971) Virgin plant communities of the Boundary Waters Canoe Area. USDA Forest Service RP-NC-63 North Central Experiment Station St. Paul, MN
RE Plotnick RH Gardner RV O’Neill (1993) ArticleTitleLacunarity indices as measures of landscape texture Landscape Ecol 8 201–11 Occurrence Handle10.1007/BF00125351
RC Rothermal (1972) A mathematical model for predicting fire spread in wildland fuels. USA Forest Service RP INT-115 Intermountain Forest and Range Experiment Station Ogden, UT
KE Saxton WJ Rawls JS Romberger RI Papendick (1986) ArticleTitleEstimating generalized soil-water characteristics from texture Soil Sci Soc Am 50 1031–6
InstitutionalAuthorNameSTATSGO. (1994) State Soil Geographic (STATSGO) Data Base. Data use information. Report number 1492 U.S. Department of Agriculture National Cartography and GIS Center Fort Worth, TX
JR Tester AM Starfield LE Frelich (1997) ArticleTitleModeling for ecosystem management in Minnesota pine forests Biol Conserv 80 313–24 Occurrence Handle10.1016/S0006-3207(96)00069-9
DB Tinker DH Knight (2001) ArticleTitleTemporal and spatial dynamics of coarse woody debris in harvested and unharvested lodgepole pine forests Ecol Model 141 125–49 Occurrence Handle10.1016/S0304-3800(01)00269-1
DL Urban MF Acevedo SL Garman (1999) Scaling fine-scale processes to large-scale patterns using models derived from models: meta-models DJ Mladenoff WL Baker (Eds) Spatial modeling of forest landscape change Cambridge University Press Cambridge, UK
PT Wolter MA White (2002) ArticleTitleRecent forest cover type transitions and landscape structural changes in northeast Minnesota, USA Landscape Ecol 17 133–55 Occurrence Handle10.1023/A:1016522509857
X Zed (1995) Hi-Rez Data Climatological Series Zed X Boalsburg, PA
Acknowledgments
We wish to thank Mark White (NRRI, University of Minnesota–Duluth) and Peter Wolter (University of Wisconsin–Green Bay) for providing satellite data and stand age coverages. This project was funded by the U.S. Forest Service National Fire Plan through the North Central Research Station.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Scheller, R.M., Mladenoff, D.J., Crow, T.R. et al. Simulating the Effects of Fire Reintroduction Versus Continued Fire Absence on Forest Composition and Landscape Structure in the Boundary Waters Canoe Area, Northern Minnesota, USA. Ecosystems 8, 396–411 (2005). https://doi.org/10.1007/s10021-003-0087-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10021-003-0087-2