
Abstract
Alkaline soda lakes are unique habitats found in specific geographic regions, usually with dry climate. The Carpathian Basin is one of those regions very important for habitat and biodiversity conservation in Europe, with natural soda lakes found in Austria, Hungary and Serbia. In comparison to other two countries from Central Europe, algal biodiversity studies of saline soda lakes in Serbia are scarce. Lake Velika Rusanda has the highest measured salinity of all saline lakes in the Carpathian Basin and there were no reports of its diatom species richness and diversity till now. We conducted 2-year investigation programme to study biodiversity and seasonal dynamics of diatoms in this lake. A total of 27 diatom taxa were found, almost all of them attached to reed and much less in benthos and plankton. Five new diatom species for Serbia were recorded, Craticula halopannonica, Navicymbula pusilla, Hantzschia weyprechtii, Nitzschia thermaloides and Navicula staffordiae. The last mentioned is new for Europe as well. Lake Velika Rusanda is inhabited mostly by alkaliphilous and halophilic diatoms. Since diatoms are used as bioindicators in soda lakes, our results will improve their further application in ecological status assessment of these fragile habitats in the Carpathian Basin.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Alkaline soda lakes, a subgroup of continental salt or athalassohaline waters, are unique habitats due to their specific physical and chemical features (Lengyel et al. 2016). At first sight, these extreme environments seem as inhospitable places for most life forms, but they are inhabited by unusual extremophiles organisms (Grant 2006; Boros et al. 2014a). Soda lakes are very vulnerable due to their shallowness and different adverse anthropogenic impacts. A building of dikes and channels for water regulation and irrigation has led to a dramatic decrease in their number in Hungary and Serbia. Moreover, it is likely that the climate change (e.g., more frequent and more intensive droughts) will pose a serious threat to the structure and function of these aquatic ecosystems in the future (Williams 2002). Therefore, it is not surprising that alkaline soda lakes and ponds are listed in several nature protection legislations issued by international and national institutions. At the EU level these halophytic habitats are included in “Pannonic salt steppes and salt marshes” habitat type with high conservation priority according to the EU Habitats Directive (92/43/EC) and they are part of the Natura 2000 Network. In Serbia, the largest saline lakes are declared as protected areas (Nature park “Rusanda”, Special nature reserve “Slano Kopovo” and Special nature reserve “Okanj Bara”), while most of other caustic lakes are part of the national ecological network.
Since they are very shallow, soda dominated waters are subjected to extreme seasonal and daily variations in most of the environmental parameters (Grant 2006; Lengyel et al. 2016). For example, water temperature in summer can be very high and exceeds 30 °C while in winter it is not unusual to have soda lakes completely frozen and covered by ice. Moreover, the salinity of these water bodies can change considerably, intra- and interannually, over the hypo-, meso- and even hypersaline limits (Boros et al. 2014a). According to Boros et al. (2014a) salinity of soda lakes or pans in the Carpathian Basin ranged from 0.5 to 60.8 g L−1, which was the maximum value measured in Lake Rusanda. On the other hand, the water pH is likely to remain almost unchanged, usually at around 9.5 in soda pans of our region (Lengyel et al. 2016). Finally, high temperature, high light intensity and CO2 in abundance are responsible for ‘‘making soda lakes one of the most productive aquatic environments in the world’’ with the primary production rate almost 15 times higher than freshwater streams and lakes (Grant 2006).
Among primary producers the photoautotrophic picoplankton (PPP), usually consisted of picoeukaryotic algae and phycocyanin-rich picocyanobacteria, is dominant in alkaline soda lakes (Felföldi et al. 2009). Diatoms play relatively minor role in those ecosystems in terms of primary production. However, recent findings have shown that benthic diatoms in shallow alkaline ponds, due to their ecophysiological plasticity (e.g., good osmoregulation) and secondary photoprotective pigments, can survive not only the inhospitable conditions, but also to successfully compete with other algal organisms (Lengyel et al. 2016). They can be found attached to the stems of submerged plants (epiphyton), in a water column (plankton), and depending on light penetration, in benthos on a surface of sediment and stones (epipelon and epilithon, respectively).
There was a significant discontinuity in ecological research on alkaline soda lakes and ponds in Serbia in the last 80 years. The first and only comprehensive study focused on the physico-chemical aspects of shallow astatic water bodies was conducted at the middle of last century in the Vojvodina province (Petrovic 1980). Recently, there were some ecological studies related to chemical composition and invertebrate fauna of three large soda lakes in Serbia (Boros et al. 2014a, b; Boros and Kolpakova 2018). Finally, a review of the algological studies of soda lakes in Serbia was given by Gavrilović et al. (2018). Only Boros et al. (2014a) reported the presence of photosynthetic organisms, e.g., Nodularia crassa, from Lake Rusanda.
Two main goals of this study were: (1) to analyze the diversity of diatom communities in different habitats (epiphyton, plankton and benthos) of Lake Velika Rusanda during 2 years in the late spring and the early summer period, and (2) to analyze ionic composition (Na+, K+, Mg2+, Ca2+, Cl−, SO42−, CO32−, HCO3−) which determine the salinity of soda lake water. The results of the study will contribute to better understanding of diatom communities in soda lakes and their use for conservation and restoration measures in the future.
Materials and methods
Study site and sampling plan
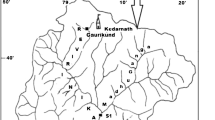
Lake Velika Rusanda is one of the largest alkaline salt lakes in Serbia. It is located in the middle Banat (the Vojvodina Province), at the left bank of the Tisza River, 16.5 km northwest of the City of Zrenjanin, between Novi Bečej and Melenci. The lake is a remnant of the old Tisza River meander and has slightly elongated and curved shape (Fig. 1). It is a shallow, semistatic water body and its maximum depth does not exceed 1.5 m (Stanković 2005). Drawing and calculation of the Lake Velika Rusanda surface (1.48 km2) were done using QGIS version 3.0.1 and the satellite image taken on 8th June 2017. The pronounced characteristic of this lake is the presence of muddy springs on its bottom. Unlike most freshwater lakes, this saline lake is a closed system (without outflow) fed by surface inflow (runoff), direct precipitation and different ground water sources including the upflow of abyssal waters (Institute for Nature Conservation of the Vojvodina Province 2011). With regard to the ionic composition, Lake Rusanda belongs to Na–Cl–SO4–HCO3 chemical type (Petrovic 1980). However, other cations and anions, such as K+, Mg2+, Ca2+, Al3+, Ba2+, Si4+, Sr2+, F−, Br−, I−, HPO42−, HAsO42− and HBO2 are important as well (Stanković 2005).

Location of the sampling sites in the Lake Velika Rusanda. Arrow indicates location of planktonic diatoms sampling; dot shows site were other diatom groups were taken
Lake Velika Rusanda is situated in area with the temperate climate which has a pronounced continental characteristics (Institute for Nature Conservation of the Vojvodina Province 2011). The coldest month is January (0.5 °C), whilst the hottest is July (22.4 °C). Based on data obtained from the weather station in Zrenjanin, and carried out over the period of 20 years (1997–2017), an average annual precipitation is 596 mm (Republic Hydrometeorological Service of Serbia 2018). Mean annual precipitation varies from 278 to 910 mm, with the usual interannual rainfall fluctuations. No overall significant trend could be observed in the local precipitation mean value for the above-mentioned period. Precipitation distribution pattern during a year shows that rainfalls reach a peak during May and June while they decrease by the end of summer. July and August are the two driest months of the year. Rainfalls are also frequent during the autumn period. Since 2011 Lake Velika Rusanda has been protected and proclaimed to be nature park (Institute for Nature Conservation of the Vojvodina Province 2011).
The samples were collected during 2-year period (2017–2018). Plankton samples were taken along the transect (Fig. 1). Diatoms living on mud (epipelic) and stone (epilithic), as well as on submerged reed stems (epiphyton) were sampled at one sampling site in the middle of the lake near the shore. Physical and chemical parameters were measured from water samples taken at the same site. In 2017 the sampling was conducted in March, while in 2018 the samples were taken in April, July and September. During the summer period in 2017 the lake completely dried up.
Physical and physico-chemical parameters
The following parameters were measured in situ: water temperature (T), pH and conductivity (COND) using Water Multimeter 18.52.01 (Eijkelkamp Agrisearch Equipment, Giesbeek, the Netherlands) and transparency (TRANS) with a Secchi disc. To identify chemical type of the soda lake four major cations and four anions were analyzed. The metal concentration (Na, K, Ca and Mg) were determined by inductively coupled plasma optical emission spectrometry (ICP-OES) using Thermo Scientific iCAP 6500 Duo ICP (Thermo Fisher Scientific, Cambridge, the UK). Multi-Element Plasma Standard Solution 4, Specpure®, 1000 μg/ml (Alfa Aesar GmbH & Co KG, Germany) was used for preparation of calibration solution for ICP-OES measurements. The quantification of the metals was done in the following emission wavelengths: Na I 818.326 nm, K I 404.721 nm, Ca II 315.887 nm, Mg I 285.213 nm. For each sample, the ICP-OES measurement was carried out in triplicate (n = 3). The reliability of measurement was approved by relative standard deviation lower than 0.5%. The analytical process quality control, performed by the use of EPA Method 200.7 LPC Solution certified reference material (CRM) for 30 analyte(s) at various concentrations (ULTRA Scientific, USA), indicated that the resulting concentrations were within 98–101%. The carbonates (CO32−) and bicarbonates (HCO3−) were calculated using alkalinity determined by the titration method according to APHA et al. (1995a). Determination of chlorides (Cl−) was done by the argentometric method according to APHA et al. (1995b). The sulphates (SO42−) were analyzed using the gravimetric method with ignition of residue following APHA et al. (1995c).
Sampling and diatom communities analysis
The plankton samples were taken directly from surface water with a 2-L plastic beaker, transferred to a plastic bucket and the final 11-L composite sample was filtered using a plankton net (Hydrobios, mesh size 25 μm) and preserved in 70% ethanol. Epipelic diatoms were collected using a glass pipette with a “cut off” bottom end, while epilithic organisms were removed from the stone surface using toothbrush and placed in a sample bottles containing ethanol (50% final concentration by volume). Epiphytic diatoms were sampled from the five submerged reed stems and fixed with ethanol (final concentration of 20% by volume). Diatom analyses were performed at the Institute of Botany and Botanical Garden ‘‘Jevremovac’’, University of Belgrade, Faculty of Biology. Their preparation for microscopic examination was done according to Taylor et al. (2007) using a hot concentrated hydrochloric acid (HCl) and supersaturated solution of potassium permanganate (KMnO4). Such method of diatom cell cleaning was slightly modified by adding a few drops of conc. HCl before all other steps. Finally, cleaned valves were mounted in Naphrax®. Micrographs were snapped using the Zeiss Axioimager.M1 microscope with DIC optics and AxioVision 4.9 software at 1500× magnification. For quantitative analysis of diatom community 400 valves per sample were counted using the Leica DM750 microscope (Leica Microsystems) with objective HI PLAN 100×/1.25 OIL (at 1000× magnification).
Results
Physical and physico-chemical parameters
Lake Velika Rusanda is one of a few soda lakes in Serbia (maybe the only one) that very rarely dries up. However, this extreme event occured in the summer 2017, when the whole lake bed was completely exposed to the air. During the study, a water temperature varied from 14.2 °C in the spring 2017 to 31.5 °C in July 2018 (Table 1). The measured pH values were always above 9.6. The conductivity was very high and revealed an increasing trend towards the end of the summer 2018.
The chemical analyses of cations and anions in this alkaline saline water indicated the dominance of sodium as well as bicarbonates, carbonates, sulphate and chlorides. Almost all measured ions showed a substantial increase in concentration from the early spring to the late summer, except calcium and magnesium (Table 1). Among all ions, sodium was the most dominant with the maximum concentration of 21,930 mg L−1 in September 2018. The data for anions revealed the opposite trend in dominance between bicarbonates and carbonates in 2017 compared to 2018. In 2017 the concentration of HCO3− was almost twice as much as the amount of CO32−.
Species richness and diversity of diatom communities
Generally, low number of diatom taxa (27 in total), irrespective of the type of microhabitat, was observed in Lake Velika Rusanda (Table 2, Fig. 3). The highest species richness (25 taxa) was recorded in the community attached to the reed (epiphyton). The number of taxa identified from plankton samples was a third of those recorded on the surface of submerged reed stems. Finally, diatom communities inhabited on mud and stones had higher species richness than the plankton community. The highest number of species identified in Lake Velika Rusanda belongs to the genus Nitzschia (8) and one species, Nitzschia supralitorea, was present in all samples. The second most diverse genus was found to be Navicula (3), and similarly to the previously mentioned here, Navicula staffordiae was a unavoidable part of all communities where diatoms were recorded.
Among 27 diatom taxa, 5 are new for the diatom flora of Serbia. Morphological and ecological characteristics of new species for Serbia are given below.
Craticula halopannonica Lange-Bertalot (Fig. 3, Figs. 1, 2)
Reference. Lange-Bertalot 2001 (p. 85, Figs. 1–6: 113)
Description. Valve lanceolate with protracted, rostrate to subcapitate ends. Valve length 56.3–64.3 µm, breadth 9.1–12.3 µm. Raphe filiform, axial area narrow, linear, central area absent. Striae almost parallel, slightly convergent at the ends, 17–19/10 µm.
Distribution in Serbia. C. halopannonica was recorded in Lake Velika Rusanda in epilithic and epipelic communities (sampling site—Fig. 1). The relative abundance in epilithic community was 0.49%, while in epipelic was 27.01%.
Navicula staffordiae Bahls (Fig. 3, Figs. 21–24)
Reference. Bahls 2012 (Figs. 9–15, 26–28)
Description. Valve narrow, lanceolate with unprotracted and acutely rounded ends. Valve length 14.3–35.8 µm, breadth 4.7–6.4 µm. Raphe filiform with proximal ends deflected slightly to the primary side. Distal raphe fissures are hooked to the secondary side. Axial area very narrow. The central area small, elliptic, asymmetric and bordered by 1–4 shortened striae. Striae weakly radiate, becoming convergent toward the ends, 14–16/10 µm. Areolae usually visible in LM, c. 33/10 µm.
Distribution in Serbia. N. staffordiae was recorded in Lake Velika Rusanda in plankton, epiphytic, epilithic and epipelic communities (transect and sampling site—Fig. 1). The relative abundance in plankton was 1.25%, epiphyton between 13.75 and 26.67%, epilithon 1.92–30.54%, while in epipelon it was even 42.24%.
Navicymbula pusilla (Grunow) Krammer (Fig. 3, Figs. 17, 18)
Basionym. Cymbella pusilla Grunow
Reference. Krammer 2003 (p. 136, Figs. 21–30; p. 138, Figs. 1–24; p. 139, Figs. 1–6; p. 140, Figs. 1–6; p. 141, Figs. 1–10)
Description. Valve moderately dorsiventral and semilanceolate. Dorsal margin moderately arched, ventral margin slightly convex or straight. Ends not protracted and variable rounded. Valve length 26.87–27.59 µm, breadth 5.67–5.73 µm. Axial area narrow, central area absent. Raphe filiform and terminal fissures hooked to the dorsal side. Striae radiate, becoming convergent at the ends, in the middle 15–17/10 µm, up to 17–18/10 µm towards the ends.
Distribution in Serbia. N. pusilla was recorded in Lake Velika Rusanda in epiphytic and epilithic communities (sampling site; Fig. 1). The relative abundance in epiphyton was 2.75%, while in epilithon it was 0.49%.
Hantzschia weyprechtii Grunow (Fig. 3, Figs. 5, 6)
Reference. Krammer and Lange-Bertalot 1988 (p. 92; Figs. 1–4: 129)
Description. Ventral margins straight, dorsal convex. Valve length 47.7–53.2 µm, breadth 5.14–9 µm. Fibulae irregularly distant, 5–9/10 µm. Striae visible in LM, 28–29/10 µm.
Distribution in Serbia. H. weyprechtii was recorded in Lake Velika Rusanda in epiphytic, epilithic and epipelic communities (sampling site—Fig. 1).
Nitzschia thermaloides Hustedt (Fig. 3, Figs. 12, 13).
Reference. Lange-Bertalot et al. 2017 (p. 108; Figs. 10–12: 462).
Description. Valve linear to linear-lanceolatae, concave retracted in the centre with obtusely wedge-shaped ends. Valve length 17.07–32.94 µm, breadth 3.64–5.54 µm. Fibulae sometimes with central gap, 14–19/10 µm.
Distribution in Serbia: N. thermaloides was recorded in Lake Velika Rusanda in epiphytic, epilithic and epipelic communities (sampling site—Fig. 1). The relative abundance in epiphytic community was 3.33–5%, in epilithic 0.99–16.75%, while in epipelon it was 0.57%.
It should be mentioned that new diatom species in the flora of Serbia (C. halopannonica, N. staffordiae, N. pusilla, H. weyprechtii, N. thermaloides) were found in Lake Velika Rusanda at low elevation (82 m a.s.l.). The lake was characterized by an alkaline water (pH above 9.6), high conductivity (14.57–37.80 mS cm−1), temperatures which can reach 31.5 °C during the summer period and high concentrations of sodium, chlorides, sulphate and (bi)carbonates.
The values of Shannon–Wiener index (H calculated with the natural logarithm) indicated different seasonal pattern between communities. Firstly, it is interesting that the highest H values for plankton and epiphyton were recorded in spring 2017 (1.47 and 1.45, respectively), while in the spring next year (after the lake had dried up in the summer 2017) the diversity of these communities was very poor. Secondly, the H-maxima for diatom communities on mud and stone were recorded in the summer 2018 (1.53 and 1.32, respectively).
Seasonal dynamics of diatom communities
An interesting feature of diatom communities in this soda lake is unpredictability in terms of their general presence and absence. For example, diatoms were not recorded in plankton in the summer, but only in the spring period. On the other hand, diatom assemblage on mud existed mainly in the summer.
Considering the diatom community structure, plankton showed different patterns of dominant species in comparison with other two communities (Fig. 2). In the spring 2017, Craticula halophila was the most abundant species in plankton (36.3%), but this species was rarely found among epiphytes (0.5%). Similarly, Anomoeoneis sphaerophora was among dominant diatom species only in plankton (27.3%). Among species attached to the surface of submerged reed stems Surirella brebissonii var. kuetzingii was most frequently found in the spring 2017 (46.3%). It is worth mentioning that some individuals of this species had modified valval shape (Fig. 3, Fig. 26). In the spring next year, a completely different diatom community structure was observed, both in plankton and epiphyton. On the other hand, the epiphytic community was exclusively comprised of two Nitzschia species, N. supralitorea (95.0%) and N. thermaloides (5.0%). Additionally, these two species were dominant diatoms on the surface of submerged stones in the spring 2018, 67.0% and 16.7%, respectively, but other species were recorded as well (S. brebissonii var. kuetzingii, Halamphora veneta and N. staffordiae).

Composition of diatom community in Lake Velika Rusanda in the spring period: a plankton (spring 2017); b epipelon (summer 2018) c epiphyton (spring 2017); d epiphyton (summer 2018); e epilithon (spring 2018) and f epilithon (summer 2018)

Light microscope (LM) micrographs. Figures 1, 2 Craticula halopannonica; Figs. 3, 4 C. halophila; Figs. 5, 6 Hantzschia weyprechtii; Figs. 7–11 Nitzschia inconspicua; Figs. 12, 13 N. thermaloides; Figs. 14–16 N. supralitorea; Figs. 17, 18 Navicymbula pusilla; Fig. 19 Anomoeoneis sphaerophora; Fig. 20 A. sphaerophora f. costata; Figs. 21–24 Navicula staffordiae; Figs. 25, 26 Surirella brebissonii var. kuetzingii. Scale bar = 10 µm
In the summer period of 2018, there were only a slight difference in community structure among diatoms attached to the reed, stones and mud surface. In epiphyton and epilithon, Nitzschia inconspicua prevailed (55.2% and 43.8%, respectively), followed by Navicula staffordiae and N. supralitorea. In epipelon two dominant species were N. staffordiae and C. halopannonica (42.2% and 27.0%, respectively).
Discussion
In the summer period of 2017 Lake Velika Rusanda completely dried up. This lake belongs to the group of semistatic water bodies which means that it rarely dries up. As for every closed-basin lake the ratio between precipitation and evaporation is crucial for its water level fluctuations. According to the following climate change scenario (Vuković et al. 2018) it can be expected a rise in mean temperature by 2.5 °C, a decrease of summer precipitation and an increase in the incidence of high precipitation events in Serbia by the end of the century. These factors would have a dramatic effect on hydrological regime of Lake Velika Rusanda and, as a consequence, on its ecosystem processes.
The increase of salinity toward the end of the summer 2018 was the result of negative balance between precipitation and evaporation in the warmer part of the year. If we take into account an average salinity, according to salinity categorization given by Hammer (1986), Lake Velika Rusanda can be considered as a hyposaline lake. However, we recorded much higher salt content in this lake (calculated as the sum of 8 major ions) toward the end of the summer. Boros et al. (2014a) measured the similar maximum salinity in this water body (60.8 g L−1). Apart from seasonal increase, an interannual rise in salinity can be expected, particularly in the warmer part of a year, because climate change scenarios forecast more frequent summer droughts in this area.
Bearing in mind that the salinity depends on the season, the ion content was based on the percentage of equivalent ions (> 25%) in total cations and anions using the early summer sample (2017). The analysis of ionic composition confirmed previous findings (Petrovic 1980; Boros et al. 2014b) that water of Lake Velika Rusanda belongs to Na–HCO3–SO4–Cl chemical type. The only difference was in the sequence of the ions dominance (e.g., Na–Cl–SO4–HCO3 in Petrovic 1980).
In Lake Velika Rusanda in all water samples where temperature was above 25 °C, CO32− dominated over HCO3−. In the summer period, biologically driven process of sodium bicarbonate breaking down (phytoplankton assimilation of CO2) leads to the increase in carbonate concentration in water (Boros et al. 2014a). One part of produced Na2CO3 precipitates from the water (for instance above 25 °C usually thermonatrite Na2CO3 × H2O is formed), while at the same time, some soda molecules react with calcium bicarbonates forming Ca2CO3 or carbonate calcareous mud sediment (Boros et al. 2014a).
When it comes to the structure of diatom communities in this alkaline soda lake, a low species richness (from 9 taxa in plankton up to 25 in epiphyton) as well as low diversity (H usually < 1.5) were recorded. The study of benthic diatoms from 39 soda lakes in Central Europe revealed similar results indicating lower species number in soda waterbodies compared to freshwater lakes (Stenger-Kovács et al. 2016). The authors of the above-mentioned study explained that by higher stress and reduced habitat heterogeneity of alkaline soda lakes and pointed to the importance of maintaining the specific diatom community as an indicator of their preserved natural limnological and hydrological characteristics.
More taxa were observed within epiphytic than planktonic and benthic diatom communities. Diatoms that were found on submerged reed stems, but were lacking on mud and stone (such as Craticula halophila, Tryblionella hungarica, etc.) are mostly benthic species recorded in lakes with similar environmental conditions (Neusiedlersee, the limnology of a shallow lake in Central Europe 1979; Subakov–Simic et al. 2004; Krizmanić et al. 2008). Hammer (1986) stated in his comprehensive publication regarding salt lakes that such waterbodies are sometimes too turbid, thus the light limitation prevents benthic and planktonic diatoms to grow. Since we sampled mud and stone near the shore within euphotic zone, the occurrence of these taxa was not hindered by light. It is more likely that these taxa would be found in benthos if we had sampled more frequently. Similarly, the diatoms observed in the water column are not true planktonic species. For instance, above-mentioned C. halophila, earlier known as Navicula halophila, was dominant species in plankton in spring, but it is mostly benthic littoral species (Veres et al. 1995; El Hamouti and Gibert Beotas 2012). Also the other authors found C. halophila in plankton and benthos of shallow alkaline lakes (Subakov–Simic et al. 2004; Żelazna-Wieczorek et al. 2015).
The diatom communities in Lake Velika Rusanda showed high fluctuations in diversity (H) related to the season as well as the habitat type. Planktonic diatoms were the most diverse group in the spring period. Our results are in accordance with the findings of Krizmanić et al. (2008) who have observed the increase in diversity of planktonic diatoms in salt marshes that experienced a drop in salinity. On the other hand, the highest diversity of benthic diatoms (epipelon and epilithon) was in the summer. Lengyel et al. (2016) in their study of benthic diatom composition in three soda pans noticed that H values increased from winter to spring and peaked in summer. In turbid soda lakes the growth of benthic algae is limited by light, but in summer when the lake water level is lower, the light can easily reach the lake bottom.
When it comes to the ecological preferences of diatoms, soda lakes in the Pannonian Basin are inhabited mostly by alkaliphilous, brackish–fresh, and fresh–brackish species (Van Dam et al. 1994; Lange-Bertalot et al. 2017, Gavrilović et al. 2018). Most of the described taxa in Lake Velika Rusanda are found in brackish waters and inland saline waters with medium to higher electrolyte content (Krammer and Lange-Bertalot 1988; Van Dam et al. 1994; Lange-Bertalot 2001; Lange-Bertalot et al. 2017). Autecology information for Nitzschia palea is still insufficiently known due to their vague LM characteristics (Lange-Bertalot et al. 2017). However, in our samples we recorded taxa which are widely distributed, in different type of eutrophic to polytrophic habitat with medium to moderately high electrolyte contents, such as Nitzschia supralitorea, Surirella brebissonii, S. brebissonii var. kuetzingii (Lange-Bertalot et al. 2017). In two epiphytic spring samples (2017 and 2018), S. brebissonii var. kuetzingii and N. supralitorea, both fresh–brackish species dominated in Lake Velika Rusanda, while in the summer period N. inconspicua, brackish-fresh species, prevailed. Additionally, two brackish species of the same genus (C. halophila and C. halopannonica) showed different microhabitat preference in this lake. The first was the most frequent diatom species in the plankton (spring sample), together with alkalibiontic species A. sphaerophora, whilst the second (C. halopannonica) dominated in the epipelon (summer sample). When it comes to A. sphaerophora Blinn (1993) reported a significant correlation between this species and HCO3− concentration. This could be one possible explanation for A. sphaerophora dominance in the spring in Lake Velika Rusanda, because HCO3− anions were much higher in the spring compared to other seasons. C. halopannonica, a typical species from saline area, is probably more widespread and it has similar morphological characteristics with C. halophila. According to Lange-Bertalot (2001) C. halopannonica is clearly distinguished from C. halophila by protracted to rostrate ends and larger valves. Micrographs from our samples confirmed this. Among diatoms recorded for the first time in Serbia, N. staffordiae showed the widest microhabitat preference in Lake Velika Rusanda (Table 2). Till now, this species has been known only from a type locality (Sand Creek, Garfield County, Montana, the USA) with alkaline water (pH 8.85), conductivity of 1.756 mS cm−1and temperature of 21.8 °C (Bahls 2012). Compared to our results, N. staffordiae was recorded at the site with much higher conductivity (14.57–20.50 mS cm−1), but in September 2018 when conductivity was 37.80 mS cm−1 this species was not recorded. According to the literature data, Navicymbula pusilla was recorded in waters with high Ca- and Cl-salinity, and with moderate to high electrolyte content (Krammer 2003; Potapova 2011; Lange-Bertalot et al. 2017). Our results showed the dominance of sodium ions as well as bicarbonates, carbonates, sulphates, and chlorides. The concentration of Ca2+ is significantly lower compared to other ions in the water. Distribution of H. weyprechtii is not well-known and recent literature data note its appearance in Nakdong River estuary, Korea (Joh 2014) and in the western Mediterranean basin of Turkey (Toudjani et al. 2017). The water bodies in the western Mediterranean basin of Turkey are characterized by alkaline water with pH ranging between 8.1 and 8.7, temperature between 14.2 and 23.8 °C and conductivity values between 0.179 and 0.776 mS cm−1. In comparison with our results this taxon was found in a habitat with higher pH as well as much higher conductivity values. N. thermaloides is characteristic for marine coasts, brackish river estuaries and salinized running waters (Lange-Bertalot et al. 2017). According to Ros et al. (2009), N. thermaloides was found in epilithic and epipelon communities in alkaline, saline habitat at 17.86 °C, pH 7.75 and conductivity values of 11.91 mS cm−1. Compared to our results, N. thermaloides was recorded in water of higher temperature (25.3–31.5 °C), higher alkalinity and pH (9.64–9.70) and elevated conductivity (14.57–17.38 mS cm−1).
Conclusions
Alkaline soda lakes are poorly investigated in this part of the Carpathian Basin, but represent important centers of biodiversity and deserve to be considered in conservation proposals. In Lake Velika Rusanda we recorded 27 diatom taxa, among which five are new taxa for diatom flora of Serbia. Additionally, this is the first study on the diatoms of Lake Velika Rusanda. Future ultimate goal should focus on conservation measures of these unique and fragile habitats in the Carpathian Basin and diatoms can be used as good indicators of ecological conditions. Our study provides an overview of species richness and diatom diversity of Lake Velika Rusanda which is the first step. It is necessary to conduct more research to get more precise data on diatom richness in other alkaline soda lakes in this part of the Carpathian Basin.
References
APHA, AWWA, WPCF (1995a) Method 2320 B. In: Eaton AD, Clesceri LS, Greenberg AE (eds) Standard methods for the examination of water and wastewater. American Public Health Association, Washington, pp 2–26
APHA, AWWA, WPCF (1995b) Method 4500-Cl− B. In: Eaton AD, Clesceri LS, Greenberg AE (eds) Standard methods for the examination of water and wastewater. American Public Health Association, Washington, pp 4–49
APHA, AWWA, WPCF (1995c) Method 4500-SO4 2− C. In: Eaton AD, Clesceri LS, Greenberg AE (eds) Standard methods for the examination of water and wastewater. American Public Health Association, Washington, pp 4–135
Bahls LL (2012) Seven new species in Navicula sensu stricto from the Northern Great Plains and Northern Rocky Mountains. Nova Hedwig Beih 141:19–37
Blinn DW (1993) Diatom community structure along physicochemical gradients in saline lakes. Ecology 74:1246–1263
Boros E, Kolpakova M (2018) A review of the defining chemical properties of soda lakes and pans: an assessment on a large geographic scale of Eurasian inland saline surface waters. PLoS One 13(8):e0202205. https://doi.org/10.1371/journal.pone.0202205
Boros E, Ecsedi Z, Olàh J (2014a) Ecology and management of soda pans in the Carpathian Basin. Hortobágy Environmental Association, Balmazújváros
Boros E, Zs Horváth, Wolfram G, Vörös L (2014b) Salinity and ionic composition of the shallow astatic soda pans in the Carpathian Basin. Ann Limnol-Int J Lim 50:59–69. https://doi.org/10.1051/limn/2013068
El Hamouti N, Gibert Beotas L (2012) Distribution of diatoms in the Baza Lacustrine system, SE, Spain. Quaternaire 23(3):253–260
Felföldi T, Somogyi B, Márialigeti K, Vörös L (2009) Characterization of photoautotrophic picoplankton assemblages in alkaline lakes of the Carpathian Basin (Central Europe). J Limnol 68(2):385–395. https://doi.org/10.4081/jlimnol.2009.385
Gavrilović B, Ćirić M, Vesić A, Vidaković D, Novaković B, Živanović M (2018) Biodiversity overview of soda pans in the Vojvodina region (Serbia). J Geogr Inst Jovan Cvijic SASA 68(2):195–214
Grant WD (2006) Alkaline environments and biodiversity. In: Gerday C, Glansdorff N (eds) Extremophilies, (Life under extreme environmental condition), Developed under the Auspices of the UNESCO. EOLSS Publishers, Oxford, UK, http://www.eolss.net
Hammer UT (1986) Saline lake ecosystems of the world. Dr W. Junk Publishers, Dordrecht
Institute for Nature Conservation of Vojvodina Province (2011) Studija zaštite: Park Prirode “Rusanda”—Predlog za stavljanje pod zaštitu kao zaštićenog područja II kategorije. Novi Sad, Srbija, pp. 188. (in Serbian)
Joh G (2014) The diverse species of the genus Hantzschia (Bacillariophyta) in sand flats of the Nakdong River estuary in Korea. J Ecol Environ 37(4):245–255
Krammer K (2003) Cymbopleura, Delicata, Navicymbula, Gomphocymbellopsis, Afrocymbella. In: Lange-Bertalot H (ed) Diatoms of Europe, Diatoms of the European Inland waters and comparable habitats, Vol. 4. A.R.G. Gantner Verlag K.G., Rugell, pp 1–529
Krammer K, Lange-Bertalot H (1988) Bacillariophyceae. 2. Teil: Bacillariaceae, Epithemiaceae, Surirellaceae. In: Ettl H, Gerloff J, Heynig H, Mollenhauer D (eds) Süßwasserflora von Mitteleuropa, 2/2. G. Fischer, Stuttgart, pp 1–596
Krizmanić J, Subakov–Simić G, Cvijan M, Karadžić V (2008) Diatoms of the three salt marshes in Vojvodina (Serbia). Abstract book. 20th International Diatom Symposium, Dubrovnik, Croatia
Lange-Bertalot H (2001) Navicula sensu stricto. 10 genera separated from Navicula sensu lato. Frustulia. In: Lange-Bertalot H (ed) Diatoms of Europe: diatoms of the European inland waters and comparable habitats, Vol. 2. A.R.G. Gantner Verlag. K.G., Ruggell, pp 1–526
Lange-Bertalot H, Hofmann G, Werum M, Cantonati M (2017) Freshwater benthic diatoms of Central Europe: over 800 common species used in ecological assessments. Koeltz Botanical Books, Schmitten-Oberreifenberg (English edition with updated taxonomy and added species)
Lengyel E, Padisák J, Hajnal É, Szabó B, Pellinger A, Stenger-Kovács C (2016) Application of benthic diatoms to assess efficiency of conservation management: a case study on the example of three reconstructed soda pans, Hungary. Hydrobiologia 777(1):95–110
Petrović G (1980) On the chemistry of some salt lakes and ponds in Yugoslavia. Hydrobiologia 81:195–200
Potapova M (2011) Navicymbula pusilla. Diatoms of the United States. http://westerndiatoms.colorado.edu/taxa/species/navicymbula_pusilla. Accessed 21 Mar 2018
Ros MD, Marín-Murcia JP, Aboal M (2009) Biodiversity of diatom assemblages in a Mediterranean semiarid stream: implications for conservation. Mar Freshw Res 60(1):14–24. https://doi.org/10.1071/MF07231
Stanković S (2005) Lakes of Serbia. Zavod za udžbenike i nastavna sredstva, Belgrade (in Serbian)
Stenger-Kovács C, Hajnal É, Lengyel E, Buczkó K, Padisák J (2016) A test of traditional diversity measures and taxonomic distinctness indices on benthic diatoms of soda pans in the Carpathian basin. Ecol Indic 64:1–8. https://doi.org/10.1016/j.ecolind.2015.12.018
Subakov–Simić G, Plemić N, Karadžić V, Cvijan M, Krizmanić J (2004) Qualitative and quantitative composition of the Slatina near Opovo. 33th Annual conference of the Yugoslav Water Pollution Control Society “Water 2004”, Conference Proceedings. Borsko jezero, Serbia, pp. 327–330 (in Serbian)
Taylor JC, Harding WR, Archibald CGM (2007) WRC Report TT 281/07: A methods manual for the collection, preparation and analysis of diatom samples version 1.0. Water Research Commission, The Republic of South Africa
Toudjani AA, Celekli A, Gümüş EY, Kayhan S, Lekesiz HÖ, Çetin T (2017) A new diatom index to assess ecological quality of running waters: a case study of water bodies in western Anatolia. Ann Limnol-Int J Lim 53:333–343. https://doi.org/10.1051/limn/2017012
Van Dam H, Mertens A, Sinkeldam J (1994) A coded checklist and ecological indicator values of freshwater diatoms from the Netherlands. Neth J Aquat Ecol 28(1):117–133. https://doi.org/10.1007/BF02334251
Veres AJ, Pienitz R, Smol JP (1995) Lake water salinity and periphytic diatom succession in three subarctic lakes, Yukon Territory, Canada. Arctic 48(1):63–70
Vuković A, Vujadinović M, Rendulić S, Đurđević V, Ruml M, Babić V, Popović D (2018) Global warming impact on climate change in Serbia for the period 1961–2100. Therm Sci. https://doi.org/10.2298/TSCI180411168V
Williams WD (2002) Environmental threats to salt lakes and the likely status of inland saline ecosystems in 2025. Environ Conserv 29:154–167. https://doi.org/10.1017/S0376892902000103
Żelazna-Wieczorek J, Olszyński RM, Nowicka-Krawczyk P (2015) Half a century of research on diatoms in athalassic habitats in central Poland. Oceanol Hydrobiol Stud 44(1):51–67
Acknowledgements
This study was financially supported by the Serbian Ministry of Education, Science, and Technological Development (Grant No. OI172001). Finally, we would like to thank to Ranko Perić and dr Aleksandra Vesić for their assistance in the field and dr Zlatko Levkov on identification help.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by A. Oren.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Vidaković, D., Krizmanić, J., Dojčinović, B.P. et al. Alkaline soda Lake Velika Rusanda (Serbia): the first insight into diatom diversity of this extreme saline lake. Extremophiles 23, 347–357 (2019). https://doi.org/10.1007/s00792-019-01088-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00792-019-01088-6