
Abstract
Abnormalities of iron homeostasis have been linked to insulin resistance, type 2 diabetes and cardiovascular disease. Carnosine, an over-the-counter food supplement with chelating properties, has been shown to decrease serum iron and improve glucose metabolism in diabetic rodents. We have previously demonstrated that carnosine supplementation prevented worsening of glucose metabolism in healthy overweight and obese middle-aged adults. Yet, the impact of carnosine on markers of iron metabolism in humans has not been investigated. We aimed to determine whether carnosine supplementation has an effect on iron parameters in overweight and obese, otherwise healthy adults. We included 26 participants, who were randomly allocated to receive 1 g carnosine (n = 14) or identical placebo (n = 12) twice daily for 12 weeks. Iron parameters including iron, ferritin, transferrin, soluble transferrin receptor, total iron binding capacity and iron saturation were measured in serum or plasma by standard commercial assays. Carnosine supplementation decreased plasma soluble transferrin receptor compared to placebo (mean change difference ± standard error: − 0.07 ± 0.03 mg/l, p = 0.04). None of the other iron parameters were different between carnosine and placebo groups. At follow-up, soluble transferrin receptor was associated inversely with urinary carnosine concentrations and positively with serum carnosinase-1 activity (both p < 0.02). Our findings suggest that carnosine may modulate iron metabolism in high-risk groups which could ameliorate insulin resistance and prevent type 2 diabetes. Larger human clinical trials are required to confirm our results.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Iron is the most abundant transition metal in the body and a potent pro-oxidant that has been known to modulate glucose and lipid metabolism (Liu et al. 2009). Abnormalities of iron homeostasis have been shown to play a role in the development of several chronic diseases such as type 2 diabetes (T2DM), cardiovascular disease (CVD), neurodegenerative diseases and non-alcoholic fatty liver disease (Liu et al. 2009; Zheng et al. 2008; Rajpathak et al. 2009a; Oshiro et al. 2011). We and others have shown that iron overload is associated with insulin resistance, cardiometabolic syndrome and T2DM via inducing oxidative stress (Liu et al. 2009; Fernandez-Real et al. 1998, 2002; Cooksey et al. 2010). High concentrations of ferritin (Gillum 2001; Lecube et al. 2008), transferrin (Podmore et al. 2016) and soluble transferrin receptor (sTfR) (Freixenet et al. 2009) have been observed in obesity and T2DM. This seems counterintuitive because sTfR levels are proportional to cellular expression of the membrane-associated transferrin receptors, reflecting cellular iron demands, and, therefore, the higher the body iron stores, the lower the sTfR concentrations. However, the findings about the direction of the relationship between cardiometabolic risk and sTfR are conflicting according to a recent meta-analysis (Orban et al. 2014). At least two studies reported positive associations of sTfR with waist circumference (Freixenet et al. 2009; Montonen et al. 2012), suggesting that the higher sTfR concentrations in abdominal obesity of hyperferritinemic subjects might be due to increased infiltration of visceral fat by macrophages. Unlike other cell types, macrophages increase their transferrin receptors as their cellular iron concentrations increase (Freixenet et al. 2009). In fact, circulating sTfR seems modulated by pleiotropic effects (Souto et al. 2003). This indicates that optimal concentration of iron parameters are required for glucose and lipid metabolism (Simcox and McClain 2013). Targeting iron metabolism has, therefore, become a compelling therapeutic strategy for the prevention and treatment of chronic diseases such as T2DM and CVD (Crielaard et al. 2017; Meng et al. 2017).
Carnosine has been shown to have chelating effects in addition to its anti-oxidative, anti-inflammatory and anti-glycating properties, thereby suggested for the prevention and treatment of T2DM and CVD (Baye et al. 2016; Boldyrev et al. 2013). In-vitro, supplementation with carnosine has prevented iron-mediated toxicity in endothelial and proximal tubular cells regardless of glucose levels, which the authors ascribed to chelation of iron (Zhang et al. 2016). Carnosine has also been shown to enhance the ferric reducing activity of human plasma by converting ferric to ferrous (Fe3+ to Fe2+), indicating its anti-oxidant ability in the presence of transition metal ions (Mozdzan et al. 2005). In streptozotocin induced diabetic rats, administration of carnosine reduced serum iron, glucose and lipid concentrations which could be due to its chelating effects or ability to scavenge free radicals and deactivate reactive oxygen species (Soliman et al. 2007). A recent human study also showed that carnosine supplementation decreased malondialdehyde levels, a marker of oxidative stress (Elbarbary et al. 2017). However, the studies showing the role of carnosine on iron and glycaemic parameters have been carried out either in vitro or in rodents. We and others have previously shown that carnosine supplementation improved glucose metabolism in humans (de Courten et al. 2016; Houjeghani et al. 2017; Elbarbary et al. 2017). To our knowledge, the effect of carnosine on markers of iron metabolism in humans has thus far not been investigated. Hence, we aimed to examine whether the chelating properties of carnosine have effect on iron parameters in non-diabetic overweight and obese individuals.
Materials and methods
Study design and population
Details of the study protocol have been reported elsewhere (de Courten et al. 2016; Baye et al. 2017; Regazzoni et al. 2016). This study is a secondary analysis of data from a pilot parallel-design, randomised, double-blind, placebo-controlled trial which was conducted in 30 healthy overweight and obese individuals at the Institute of Experimental Endocrinology, Slovak Academy of Sciences, Bratislava, Slovakia. All participants were non-diabetic based on a 75 g oral glucose tolerance test and had no sign of infection according to a physical examination and routine blood analyses. None of them were smokers, and had taken any medications or food supplements. All participants underwent a study protocol that involved anthropometric assessments, lipid profile and glycaemic measures, assessment of urinary carnosine concentrations and serum carnosinase-1 content and activity. Participants were randomly assigned either to receive 1 g oral carnosine (Flamma S.p.A, Italy) or identical placebo (1 g sucrose) twice daily for 12 consecutive weeks. All participants were asked to refrain from substantial life style changes throughout the study period as well as from vigorous physical activity and caffeine for 3 days prior their visits. Participants with significant weight change (≥ 5 kg) in the study period were excluded from the analysis. In this analysis, we have excluded three participants (1 carnosine, 2 placebo) for non-compliance with the protocol, and one additional participant (1 carnosine) due to unusually high iron parameters. All metabolic testing, blood and urine collections were performed after a 12 h overnight fast and 12 h after carnosine ingestion.
All participants provided written informed consent before enrolment into the study. The study protocol was approved by the Ethics Committee of the University Hospital Bratislava, Comenius University, Slovakia, and it adheres to the Ethical Declaration of Helsinki.
Assessment of dietary preference
Food preference questionnaire was used to determine the participants’ dietary preference for high-fat/low-fat foods based on 72 different food items. Participants rate each food hedonically on a nine-point Likert scale, with one-dislike extremely, five-neutral, and nine-like extremely. Fat preference score was computed by dividing high-fat score by low-fat score.
Measurement of urinary carnosine and creatinine concentrations and serum carnosinase activity and content
Urine and blood samples were collected in the fasting state using pre-cooled containers and tubes, respectively, centrifuged and stored at − 80 °C until analysis. Spot urine outcomes were corrected for creatinine concentrations. Urinary carnosine was measured using an internal standard and a triple quadrupole mass spectrometer (TSQ QUANTUM ULTRA, Thermo Scientific, Italy). Blank matrices for calibration curves set up were obtained by pooling urine from six volunteers aged 24–28 years who followed a lacto-ovo-vegetarian diet for 1 day before urine collection. Before pooling, biological fluids were analysed to ensure they did not contain the selected adduct above the 20% limit of quantitation. The internal standard, tyrosine hydroxylase (TH), was added at a final concentration of 35 µM for urine. Samples were then treated using analytical procedures as follows. Three independent samples were prepared for each level of the calibration curve and each was analysed in triplicate. Calibration curves were built by least square linear regression analysis by plotting the ratios between the peak areas of the analyte and the internal standard against the analyte’s nominal concentration. For liquid chromatography–electrospray ionisation–mass spectrometry (LC–ESI–MS) analysis, urine samples were treated as follows: aliquots of 150 µl were mixed with 150 µl of an aqueous solution of 4% trichloroacetic acid (TCA) (v/v) and spiked with TH to reach a final concentration of 35 µM. The sample was then centrifuged (14,000g, 10 min, 4 °C), to remove any particulate matter. The urinary supernatant was collected and an aliquot was injected in the chromatography system and analysed by LC–ESI–MS in multiple reaction monitoring protocol as previously described (Aldini et al. 2011).
Urine creatinine was measured by a colorimetric assay based on Jaffe reaction (Cayman, USA) (Heinegard and Tiderstrom 1973). The urine samples are diluted 1:10 with H2O and then treated with an alkaline picrate solution. When the creatinine reacts with the picrate, a yellow/orange coloration is formed. The colour intensity is proportional to creatinine concentration and it is measured at 500 nm. The sample creatinine concentration is determined using a creatinine standard curve. The colour derived by creatinine is then destroyed at acidic pH and the intensity is measured again as a factor of correction.
Serum carnosinase concentrations were measured using a sandwich enzyme-linked immunosorbent assay (ELISA) developed by Adelmann et al. (Adelmann et al. 2012), as per instructions. Serum carnosinase activity was quantified by fluorometric determination of liberated histidine after carnosine addition. Briefly, 10 mM carnosine (Flamma, Italy) was added to serum and the reaction was stopped after 10-min incubation at 37 °C by 600 mM TCA. For controls, TCA was added before carnosine. After centrifugation (4500 rpm, 15 min, RT), supernatant was added to a mixture of OPA (incomplete phtaldehyde with 0.2% 2-mercaptoethanol) and 4 M sodium hydroxide and fluorescence were determined after 40 min (excitation: 360 nm and emission: 465 nm).
Measurement of markers of iron metabolism
Serum iron concentration was measured by a standard commercial rate assay (Iron Kit Cat. No. 467910, Beckman Coulter Diagnostics, New South Wales, Australia). The coefficient of variation (CV) was 2.48% at 11.14 µmol/l and 3.8% at 43.56 µmol/l. The reference interval was 13–35 µmol/l. Both ferritin and transferrin concentrations were measured in plasma by standard commercial rate assays (Access/DXI Ferritin Reagent Pack Cat. No. 33020 and Transferrin Kit Cat. No. 467942, Beckman Coulter Diagnostics, New South Wales, Australia). The CV of ferritin was 4.99% at 41.21 μg/l and 8.27% at 117.51 μg/l. The reference interval for males aged above 10 years was 20–300 μg/l and for females aged 10–50 years was 15–80 μg/l. The CV of transferrin was 4.96% at 1.96 g/l and 4.64% at 2.56%, and its reference range was 2.0–3.6 g/l. Measurement of iron, ferritin and transferrin were performed using a Beckman Coulter DXC800 Analyser. Plasma sTfR concentrations were quantitatively determined by a sandwich immunoenzymatic assay (Access sTfR assay Cat. No. A32493, Beckman Coulter Diagnostics, New South Wales, Australia) using the Access Immunoassay Systems performed on a Beckman Coulter Unicel DXI 800 analyzer. The CV was 4.90% at 0.71 mg/l, 4.81% at 1.87 mg/l and 4.26% at 6.81 mg/l. The reference interval was 0–1.55 mg/l as recommended by Beckman Coulter. Total iron binding capacity was computed by multiplying the transferrin concentrations by 25.1; while iron saturation was calculated using the formula: (3.982 * iron)/transferrin. The reference intervals of iron binding capacity and iron saturation were 45–76 µmol/l and 15–46%, respectively.
Measurement of markers of inflammation
High-sensitivity C-reactive protein (hsCRP) was measured in serum using an immunoturbidimetric assay (Randox, UK). The reference range was 0.477–10 mg/dl. Serum IL-6 was quantified by a bead-based multiplex assay (LEGENDplex™ Human Metabolic Panel, Biolegend, San Diego, CA). The reference interval of IL-6 was 0–114.2 pg/ml. The CVs for hsCRP and IL-6 were < 10%.
Statistical analyses
The sample size calculation, where insulin sensitivity was the primary outcome of the trial, has been reported elsewhere (de Courten et al. 2016). Kolmogorov–Smirnov tests and visual inspection of probability and dot plots were used to assess normality of data. Appropriate data transformation was performed when needed. Results were presented as means ± standard deviations. Independent t tests were employed to determine baseline as well as change from baseline differences between carnosine and placebo groups. Pearson correlations were used to assess the relationships between markers of iron metabolism and carnosine measurements. Linear regressions were performed to determine the presence of associations between iron parameters and carnosine measurements after adjusting for age, sex and body mass index. Data analyses were performed using SAS Studio 3.4 (SAS Institute Inc., Cary, North Carolina).
Results
Baseline characteristics of the participants are presented in Table 1. Twenty-six participants were included in this analysis. Five of them were females (2 carnosine, 3 placebo). Overall mean age and body mass index of the participants were 42.7 ± 8.1 years and 31.9 ± 4.4 kg/m2, respectively, with no significant differences between carnosine and placebo groups (both p > 0.6). As previously described (de Courten et al. 2016), there were no baseline differences between groups in glycaemic parameters, blood pressure, lipid profiles, dietary preference, resting energy expenditure, and hsCRP (all p > 0.1). Both groups also did not differ in baseline urinary carnosine-creatinine ratio (0.12 ± 0.08 carnosine vs 0.11 ± 0.08 placebo, p = 0.8) or serum carnosinase-1 activity (1.8 ± 0.8 µmol/ml/hr carnosine vs 2.1 ± 0.6 µmol/ml/hr placebo, p = 0.4) or content (57.7 ± 23.5 ng/ml carnosine vs 65.2 ± 37.5 ng/ml placebo, p = 0.5). Markers of iron metabolism such as iron, ferritin, transferrin, sTfR, total iron binding capacity and iron saturation were not different between carnosine and placebo groups at baseline (all p > 0.2; Table 2). None of the iron parameters were different by gender (all p > 0.1). Change in dietary preference score did not differ between carnosine and placebo groups (mean difference ± standard deviation: − 0.72 ± 10.58 carnosine vs − 0.88 ± 27.3 placebo, p = 0.9).
Effect of carnosine supplementation on markers of iron metabolism
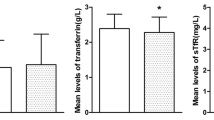
Carnosine supplementation significantly decreased the concentration of plasma sTfR compared to placebo (mean change difference ± standard error: − 0.07 ± 0.03 mg/l, p = 0.04; Table 2). Circulating serum iron, plasma ferritin, plasma transferrin, plasma iron binding capacity and serum iron saturation concentrations did not change after supplementation with carnosine compared to placebo (all p > 0.5; Table 2).
Associations between sTfR and carnosine measures and cardiometabolic parameters at follow-up
Plasma sTfR was correlated negatively with urinary carnosine-creatinine ratio (r = − 0.43, p = 0.02; Fig. 1) and positively with serum carnosinase-1 activity (r = 0.61, p = 0.0009; Fig. 2). These associations were remained significant after adjustment for age, sex, body mass index and intervention group (urinary carnosine-creatinine ratio; β-coefficient: − 0.06 nmol/ml, 95% confidence interval: − 0.12 to − 0.001, p = 0.04; serum carnosinase-1 activity; β-coefficient: 0.21 μmol/ml/hr; 95% confidence interval: 0.08–0.34; p = 0.003). Serum carnosinase-1 content was not related to plasma sTfR concentrations (r = 0.36, p = 0.07). Inflammatory markers such as hsCRP and IL-6, glycaemic or cardiovascular parameters were also not associated with plasma sTfR (all p > 0.1, data not shown).

Correlation graph shows the relationship between urinary carnosine-creatinine ratio and plasma soluble transferrin receptor concentrations at follow-up (n = 26)

Correlation graph shows the relationship between serum carnosinase-1 activity and plasma soluble transferrin receptor concentrations at follow-up (n = 26)
Discussion
We investigated whether supplementation with carnosine changes markers of iron metabolism in overweight and obese, otherwise healthy adults. We have shown for the first time that carnosine supplementation decreased plasma sTfR concentrations compared to placebo, and plasma sTfR was inversely associated with urinary carnosine-creatinine ratio and positively associated with serum carnosinase-1 activity at follow-up.
Soluble transferrin receptor is responsible for the cellular uptake of iron (Speeckaert et al. 2010), and is elevated in obesity (Freixenet et al. 2009) and insulin resistance (Fernandez-Real et al. 2007). Importantly, high sTfR was associated with increased risk of T2DM in overweight and obese individuals (Fernandez-Cao et al. 2017; Rajpathak et al. 2009b). Lifestyle intervention has been shown to reduce sTfR in high-risk groups. A study in overweight and obese children and adolescents showed a decrease in plasma sTfR after intervention with 8-month physical exercise compared to controls (Coimbra et al. 2017). Dietary weight loss with resistance training for 16 weeks also reduced serum sTfR in obese individuals compared to dietary weight loss alone and no weight loss (Fernandez-Real et al. 2009). Furthermore, a decrease in sTfR was observed after a 1-year dietary weight loss intervention in overweight and obese children compared to controls (Gong et al. 2014). In our study, we have demonstrated that concentration of plasma sTfR was reduced after carnosine supplementation compared to placebo. However, we did not find any association between sTfR and glycaemic parameters. This could be due to that all the participants were non-diabetic or the small sample size of the study. Nonetheless, we have previously shown that carnosine supplementation hampered insulin resistance and improved glucose tolerance in non-diabetic overweight and obese adults with and without impaired glucose tolerance (de Courten et al. 2016). Our data suggest that carnosine could be another strategy to prevent or delay the development of insulin resistance and T2DM in high risk groups via reducing sTfR concentrations.
Soluble transferrin receptor expression on erythroid precursors is determined by cellular iron demand (Speeckaert et al. 2010). Circulating sTfR concentration in serum reflects total body sTfR concentration (Skikne 2008). It has been suggested as a better marker of iron overload in obese individuals (Mast et al. 1998). High concentration of sTfR was commonly reported in obese people (Fernandez-Cao et al. 2017; Freixenet et al. 2009; Coimbra et al. 2017; Sharma et al. 2009; Lecube et al. 2006), which could be due to functional iron deficiency, a state in which there is insufficient iron incorporation into erythroid precursors in the case of apparently adequate body iron stores (Macdougall et al. 1989). Carnosine has been suggested to downregulate transferrin receptor protein expression in vitro (Zhang et al. 2016). In humans, low plasma carnosinase activity promotes circulating carnosine after supplementation with carnosine (Everaert et al. 2012). Consistent with this, we find that sTfR was associated inversely with urinary carnosine–creatinine ratio after adjustment for age, sex and body mass index. We have previously shown that carnosine supplementation increased urinary carnosine concentrations, but did not change serum carnosinase 1 activity or content (de Courten et al. 2016). Our findings may suggest that carnosine supplementation may improve iron uptake by erythroid precursors, thereby reducing the concentration of sTfR. Future mechanistic studies are needed to confirm this.
We report that sTfR was positively associated with serum carnosinase-1 activity after adjustment for age, sex and body mass index. Recent studies have shown that carnosinase-1 activity is modulated through an allosteric mechanism involving cysteine residue at position 102 (Cys102) (Peters et al. 2017). In particular, thiol-containing compounds, such as glutathione and cysteine, reduce carnosinase-1 activity through the formation of mixed disulfides. Since sTfR is involved in the redox homeostasis and hence in the protein cysteinylation process, we suggest that carnosinase-1 activity may be modulated allosterically by the formation of Cys102 mixed disulfides, a reaction which is most likely related to plasma sTfR concentrations.
Carnosine has been shown to chelate heavy metal ions such as cooper and zinc (Boldyrev et al. 2013). This has been shown to inhibit oxidative and inflammatory pathways (Nagai et al. 2012). In-vitro, carnosine prevented iron mediated toxicity in renal tubular cells likely due to chelating effects of carnosine (Zhang et al. 2016). Carnosine administration also resulted in decrease in serum iron concentration and improved glucose metabolism in streptozotocin induced diabetic rats (Soliman et al. 2007). In our study, however, carnosine supplementation did not change iron concentrations, which suggests that carnosine has no direct chelating effects on iron in healthy overweight or obese individuals.
Iron as a transition metal can generate reactive oxygen species via Fenton reaction, leads to the formation of a variety of toxic by-products including hydroxynonenal, and induces oxidative stress (Liu et al. 2009). sTfR to ferritin ratio was associated with urinary 8-hydroxydeoxyguanosine excretion, a marker of oxidative damage in vivo (Tuomainen et al. 2007). Furthermore, markers of oxidative stress such as malondialdehyde and total reactive oxygen species were positively correlated with serum iron parameters in patients with β-thalassemia (Choudhuri et al. 2013). Carnosine has the ability to interrupt the production of hydroxyl radicals by trapping the Fenton reaction (Velez et al. 2008; Price et al. 2001), decrease malondialdehyde formations (Liu et al. 2008), and inhibit oxidative reactions (Nagai et al. 2012), which could subsequently improve iron and glucose metabolism. It has also been shown that carnosine is the most effective and selective hydroxynonenal sequestering agent available (Aldini et al. 2014). The anti-oxidant properties of carnosine could, therefore, be a possible mechanism for the reduced sTfR.
Inflammatory cytokines including interleukins and tumour necrosis factor alpha have been shown to upregulate sTfR expression in vitro (Tsuji et al. 1991; Johnson et al. 2004). sTfR was positively associated with interleukin-6 (Coimbra et al. 2017) and C-reactive protein concentrations (Sharma et al. 2009). In rodents, carnosine suppressed the production and release of interleukin-6 and tumour necrosis factor alpha as well as C-reactive protein (Yan et al. 2009; Tsai et al. 2010). A recent human study in patients with T2DM showed that tumour necrosis factor alpha was reduced after carnosine supplementation (Houjeghani et al. 2017). In our study, we did not find any association of sTfR with IL-6 and hsCRP. This may be due to that all participants had no sign of inflammation at enrolment or the small sample size of the study. Although future studies are needed to confirm the exact mechanism, it may be possible that the reduction of chronic low-grade inflammation after carnosine supplementation may downregulate the expression of sTfR.
Advanced glycation end products (AGEs) have been shown to associate with iron overload (Choudhuri et al. 2013). AGEs are also known to contribute to increased oxidative stress and inflammation (Vlassara 2001). A recent study in patients with β-thalassemia showed positive associations between serum AGE concentrations and iron indices (Choudhuri et al. 2013). We have not measured AGEs in our study. Nonetheless, several experimental studies demonstrated that carnosine has the ability to inhibit AGEs formation produced through a non-enzymatic glycation and a direct reaction with reactive carbonyl species generated by lipid and sugar oxidation (Baye et al. 2016). In uremic patients undergoing peritoneal dialysis, supplementation with carnosine has also been shown to prevent AGEs formation by acting as a sacrificial molecule protecting protein amino groups from glycation by highly reactive carbonyl compounds (Alhamdani et al. 2007). This finding suggests that the antiglycating ability of carnosine may contribute to the reduction in sTfR concentration; however, this has to be confirmed by future mechanistic studies.
In this pilot clinical trial, comprehensive metabolic profiling of the study participants were performed; rigorous randomisation process was employed; allocation of treatment to the intervention groups was concealed; healthy participants who did not take any medications or supplements were studied; and no baseline differences between groups were observed, which all restricts the effect of confounding on reported outcome measures. We have measured several markers of iron metabolism including iron, ferritin, transferrin, sTfR, total iron binding capacity and iron saturation. We did not measure hepcidin concentrations and the participants’ actual dietary intake, which could be a limitation of this study. The small sample size of the study could be another limitation of this trial, where sample size calculation was not computed based on the outcome of iron parameters. The study was performed in overweight or obese individuals; hence, results cannot be extrapolated to another target groups, including individuals with normal weight. Larger sample size trials are therefore needed to confirm the effect of carnosine supplementation on iron metabolism.
In conclusion, we have demonstrated for the first time that carnosine supplementation reduced plasma sTfR in healthy overweight and obese humans. This suggests that carnosine may improve iron metabolism in high-risk groups and thereby prevent and delay the development of T2DM and CVD; however, future larger clinical trials are required to confirm our findings.
References
Adelmann K, Frey D, Riedl E, Koeppel H, Pfister F, Peters V, Schmitt CP, Sternik P, Hofmann S, Zentgraf HW, Navis G, van den Born J, Bakker SJ, Kramer BK, Yard BA, Hauske SJ (2012) Different conformational forms of serum carnosinase detected by a newly developed sandwich ELISA for the measurements of carnosinase concentrations. Amino Acids 43(1):143–151. https://doi.org/10.1007/s00726-012-1244-8
Aldini G, Orioli M, Rossoni G, Savi F, Braidotti P, Vistoli G, Yeum KJ, Negrisoli G, Carini M (2011) The carbonyl scavenger carnosine ameliorates dyslipidaemia and renal function in Zucker obese rats. J Cell Mol Med 15(6):1339–1354. https://doi.org/10.1111/j.1582-4934.2010.01101.x
Aldini G, Carini M, Yeum KJ, Vistoli G (2014) Novel molecular approaches for improving enzymatic and nonenzymatic detoxification of 4-hydroxynonenal: toward the discovery of a novel class of bioactive compounds. Free Radic Biol Med 69:145–156. https://doi.org/10.1016/j.freeradbiomed.2014.01.017
Alhamdani MS, Al-Azzawie HF, Abbas FK (2007) Decreased formation of advanced glycation end-products in peritoneal fluid by carnosine and related peptides. Perit Dial Int 27(1):86–89
Baye E, Ukropcova B, Ukropec J, Hipkiss A, Aldini G, de Courten B (2016) Physiological and therapeutic effects of carnosine on cardiometabolic risk and disease. Amino Acids 48(5):1131–1149. https://doi.org/10.1007/s00726-016-2208-1
Baye E, Ukropec J, de Courten MPJ, Vallova S, Krumpolec P, Kurdiova T, Aldini G, Ukropcova B, de Courten B (2017) Effect of carnosine supplementation on the plasma lipidome in overweight and obese adults: a pilot randomised controlled trial. Sci Rep 7(1):17458. https://doi.org/10.1038/s41598-017-17577-7
Boldyrev AA, Aldini G, Derave W (2013) Physiology and pathophysiology of carnosine. Physiol Rev 93(4):1803–1845. https://doi.org/10.1152/physrev.00039.2012
Choudhuri S, Dutta D, Sen A, Chowdhury IH, Mitra B, Mondal LK, Saha A, Bhadhuri G, Bhattacharya B (2013) Role of N-epsilon- carboxy methyl lysine, advanced glycation end products and reactive oxygen species for the development of nonproliferative and proliferative retinopathy in type 2 diabetes mellitus. Mol Vis 19:100–113
Coimbra S, Catarino C, Nascimento H, Ines Alves A, Filipa Medeiros A, Bronze-da-Rocha E, Costa E, Rocha-Pereira P, Aires L, Seabra A, Mota J, Ferreira Mansilha H, Rego C, Santos-Silva A, Belo L (2017) Physical exercise intervention at school improved hepcidin, inflammation, and iron metabolism in overweight and obese children and adolescents. Pediatr Res 82(5):781–788. https://doi.org/10.1038/pr.2017.139
Cooksey RC, Jones D, Gabrielsen S, Huang J, Simcox JA, Luo B, Soesanto Y, Rienhoff H, Abel ED, McClain DA (2010) Dietary iron restriction or iron chelation protects from diabetes and loss of beta-cell function in the obese (ob/ob lep-/-) mouse. Am J Physiol Endocrinol Metab 298(6):E1236–E1243. https://doi.org/10.1152/ajpendo.00022.2010
Crielaard BJ, Lammers T, Rivella S (2017) Targeting iron metabolism in drug discovery and delivery. Nat Rev Drug Discov. https://doi.org/10.1038/nrd.2016.248
de Courten B, Jakubova M, de Courten MP, Kukurova IJ, Vallova S, Krumpolec P, Valkovic L, Kurdiova T, Garzon D, Barbaresi S, Teede HJ, Derave W, Krssak M, Aldini G, Ukropec J, Ukropcova B (2016) Effects of carnosine supplementation on glucose metabolism: pilot clinical trial. Obesity (Silver Spring) 24(5):1027–1034. https://doi.org/10.1002/oby.21434
Elbarbary NS, Ismail EAR, El-Naggar AR, Hamouda MH, El-Hamamsy M (2017) The effect of 12 weeks carnosine supplementation on renal functional integrity and oxidative stress in pediatric patients with diabetic nephropathy: a randomized placebo-controlled trial. Pediatr Diabetes. https://doi.org/10.1111/pedi.12564
Everaert I, Taes Y, De Heer E, Baelde H, Zutinic A, Yard B, Sauerhofer S, Vanhee L, Delanghe J, Aldini G, Derave W (2012) Low plasma carnosinase activity promotes carnosinemia after carnosine ingestion in humans. Am J Physiol Renal Physiol 302(12):F1537–F1544. https://doi.org/10.1152/ajprenal.00084.2012
Fernandez-Cao JC, Arija V, Aranda N, Basora J, Diez-Espino J, Estruch R, Fito M, Corella D, Salas-Salvado J (2017) Soluble transferrin receptor and risk of type 2 diabetes in the obese and nonobese. Eur J Clin Invest 47(3):221–230. https://doi.org/10.1111/eci.12725
Fernandez-Real JM, Ricart-Engel W, Arroyo E, Balanca R, Casamitjana-Abella R, Cabrero D, Fernandez-Castaner M, Soler J (1998) Serum ferritin as a component of the insulin resistance syndrome. Diabetes Care 21(1):62–68
Fernandez-Real JM, Lopez-Bermejo A, Ricart W (2002) Cross-talk between iron metabolism and diabetes. Diabetes 51(8):2348–2354
Fernandez-Real JM, Moreno JM, Lopez-Bermejo A, Chico B, Vendrell J, Ricart W (2007) Circulating soluble transferrin receptor according to glucose tolerance status and insulin sensitivity. Diabetes Care 30(3):604–608. https://doi.org/10.2337/dc06-1138
Fernandez-Real JM, Izquierdo M, Moreno-Navarrete JM, Gorostiaga E, Ortega F, Martinez C, Idoate F, Ricart W, Ibanez J (2009) Circulating soluble transferrin receptor concentration decreases after exercise-induced improvement of insulin sensitivity in obese individuals. Int J Obes (Lond) 33(7):768–774. https://doi.org/10.1038/ijo.2009.99
Freixenet N, Remacha A, Berlanga E, Caixas A, Gimenez-Palop O, Blanco-Vaca F, Bach V, Baiget M, Sanchez Y, Felez J, Gonzalez-Clemente JM (2009) Serum soluble transferrin receptor concentrations are increased in central obesity. Results from a screening programme for hereditary hemochromatosis in men with hyperferritinemia. Clin Chim Acta 400(1–2):111–116. https://doi.org/10.1016/j.cca.2008.10.019
Gillum RF (2001) Association of serum ferritin and indices of body fat distribution and obesity in Mexican American men—the Third National Health and Nutrition Examination Survey. Int J Obes Relat Metab Disord 25(5):639–645. https://doi.org/10.1038/sj.ijo.0801561
Gong L, Yuan F, Teng J, Li X, Zheng S, Lin L, Deng H, Ma G, Sun C, Li Y (2014) Weight loss, inflammatory markers, and improvements of iron status in overweight and obese children. J Pediatr 164(4):795–800.e792. https://doi.org/10.1016/j.jpeds.2013.12.004
Heinegard D, Tiderstrom G (1973) Determination of serum creatinine by a direct colorimetric method. Clin Chim Acta 43(3):305–310
Houjeghani S, Kheirouri S, Faraji E, Jafarabadi MA (2017) l-carnosine supplementation attenuated fasting glucose, triglycerides, advanced glycation end products and tumor necrosis factor alpha levels in patients with type 2 diabetes: a double-blind placebo-controlled randomized clinical trial. Nutr Res. https://doi.org/10.1016/j.nutres.2017.11.003
Johnson D, Bayele H, Johnston K, Tennant J, Srai SK, Sharp P (2004) Tumour necrosis factor alpha regulates iron transport and transporter expression in human intestinal epithelial cells. FEBS Lett 573(1–3):195–201. https://doi.org/10.1016/j.febslet.2004.07.081
Lecube A, Carrera A, Losada E, Hernandez C, Simo R, Mesa J (2006) Iron deficiency in obese postmenopausal women. Obesity (Silver Spring) 14(10):1724–1730. https://doi.org/10.1038/oby.2006.198
Lecube A, Hernandez C, Pelegri D, Simo R (2008) Factors accounting for high ferritin levels in obesity. Int J Obes (Lond) 32(11):1665–1669. https://doi.org/10.1038/ijo.2008.154
Liu WH, Liu TC, Yin MC (2008) Beneficial effects of histidine and carnosine on ethanol-induced chronic liver injury. Food Chem Toxicol 46(5):1503–1509. https://doi.org/10.1016/j.fct.2007.12.013
Liu Q, Sun L, Tan Y, Wang G, Lin X, Cai L (2009) Role of iron deficiency and overload in the pathogenesis of diabetes and diabetic complications. Curr Med Chem 16(1):113–129
Macdougall IC, Hutton RD, Cavill I, Coles GA, Williams JD (1989) Poor response to treatment of renal anaemia with erythropoietin corrected by iron given intravenously. BMJ 299(6692):157–158
Mast AE, Blinder MA, Gronowski AM, Chumley C, Scott MG (1998) Clinical utility of the soluble transferrin receptor and comparison with serum ferritin in several populations. Clin Chem 44(1):45–51
Meng G, Yang H, Bao X, Zhang Q, Liu L, Wu H, Du H, Xia Y, Shi H, Guo X, Liu X, Li C, Su Q, Gu Y, Fang L, Yu F, Sun S, Wang X, Zhou M, Jia Q, Guo Q, Song K, Huang G, Wang G, Wu Y, Niu K (2017) Increased serum ferritin levels are independently related to incidence of prediabetes in adult populations. Diabetes Metab 43(2):146–153. https://doi.org/10.1016/j.diabet.2016.07.028
Montonen J, Boeing H, Steffen A, Lehmann R, Fritsche A, Joost HG, Schulze MB, Pischon T (2012) Body iron stores and risk of type 2 diabetes: results from the European Prospective Investigation into Cancer and Nutrition (EPIC)-Potsdam study. Diabetologia 55(10):2613–2621. https://doi.org/10.1007/s00125-012-2633-y
Mozdzan M, Szemraj J, Rysz J, Nowak D (2005) Antioxidant properties of carnosine re-evaluated with oxidizing systems involving iron and copper ions. Basic Clin Pharmacol Toxicol 96(5):352–360. https://doi.org/10.1111/j.1742-7843.2005.pto_03.x
Nagai R, Murray DB, Metz TO, Baynes JW (2012) Chelation: a fundamental mechanism of action of AGE inhibitors, AGE breakers, and other inhibitors of diabetes complications. Diabetes 61(3):549–559. https://doi.org/10.2337/db11-1120
Orban E, Schwab S, Thorand B, Huth C (2014) Association of iron indices and type 2 diabetes: a meta-analysis of observational studies. Diabetes Metab Res Rev 30(5):372–394. https://doi.org/10.1002/dmrr.2506
Oshiro S, Morioka MS, Kikuchi M (2011) Dysregulation of iron metabolism in Alzheimer’s disease, Parkinson’s disease, and amyotrophic lateral sclerosis. Adv Pharmacol Sci 2011:378278. https://doi.org/10.1155/2011/378278
Peters V, Schmitt CP, Weigand T, Klingbeil K, Thiel C, van den Berg A, Calabrese V, Nawroth P, Fleming T, Forsberg E, Wagner AH, Hecker M, Vistoli G (2017) Allosteric inhibition of carnosinase (CN1) by inducing a conformational shift. J Enzyme Inhib Med Chem 32(1):1102–1110. https://doi.org/10.1080/14756366.2017.1355793
Podmore C, Meidtner K, Schulze MB, Scott RA, Ramond A, Butterworth AS, Di Angelantonio E, Danesh J, Arriola L, Barricarte A, Boeing H, Clavel-Chapelon F, Cross AJ, Dahm CC, Fagherazzi G, Franks PW, Gavrila D, Grioni S, Gunter MJ, Gusto G, Jakszyn P, Katzke V, Key TJ, Kuhn T, Mattiello A, Nilsson PM, Olsen A, Overvad K, Palli D, Quiros JR, Rolandsson O, Sacerdote C, Sanchez-Cantalejo E, Slimani N, Sluijs I, Spijkerman AM, Tjonneland A, Tumino R, van der DL A, van der Schouw YT, Feskens EJ, Forouhi NG, Sharp SJ, Riboli E, Langenberg C, Wareham NJ (2016) Association of multiple biomarkers of iron metabolism and type 2 diabetes: the EPIC-interact study. Diabetes Care 39(4):572–581. https://doi.org/10.2337/dc15-0257
Price DL, Rhett PM, Thorpe SR, Baynes JW (2001) Chelating activity of advanced glycation end-product inhibitors. J Biol Chem 276(52):48967–48972. https://doi.org/10.1074/jbc.M108196200
Rajpathak SN, Crandall JP, Wylie-Rosett J, Kabat GC, Rohan TE, Hu FB (2009a) The role of iron in type 2 diabetes in humans. Biochim Biophys Acta 1790(7):671–681. https://doi.org/10.1016/j.bbagen.2008.04.005
Rajpathak SN, Negassa A, Kabat GC, Wylie-Rosett J, Rohan TE, Crandall J, Diabetes Prevention Program Research G (2009b) Elevated body iron stores predict the conversion from impaired glucose tolerance to type 2 diabetes. Diabetes Obes Metab 11(5):472–479. https://doi.org/10.1111/j.1463-1326.2008.00985.x
Regazzoni L, de Courten B, Garzon D, Altomare A, Marinello C, Jakubova M, Vallova S, Krumpolec P, Carini M, Ukropec J, Ukropcova B, Aldini G (2016) A carnosine intervention study in overweight human volunteers: bioavailability and reactive carbonyl species sequestering effect. Sci Rep 6:27224. https://doi.org/10.1038/srep27224
Sharma AP, McKenna AM, Lepage N, Nieuwenhuys E, Filler G (2009) Relationships among serum iron, inflammation, and body mass index in children. Adv Pediatr 56:135–144. https://doi.org/10.1016/j.yapd.2009.08.014
Simcox JA, McClain DA (2013) Iron and diabetes risk. Cell Metab 17(3):329–341. https://doi.org/10.1016/j.cmet.2013.02.007
Skikne BS (2008) Serum transferrin receptor. Am J Hematol 83(11):872–875. https://doi.org/10.1002/ajh.21279
Soliman KM, Mohamed AM, Metwally NS (2007) Attenuation of some metabolic deteriorations induced by diabetes mellitus using carnosine. J Appl Sci 7:2252–2260
Souto JC, Remacha A, Buil A, Almasy L, Blangero J, Fontcuberta J (2003) Genetic determinants of iron metabolism plasma phenotypes and their relationship with risk of thrombosis. Haematologica 88(12):1436–1438
Speeckaert MM, Speeckaert R, Delanghe JR (2010) Biological and clinical aspects of soluble transferrin receptor. Crit Rev Clin Lab Sci 47(5–6):213–228. https://doi.org/10.3109/10408363.2010.550461
Tsai SJ, Kuo WW, Liu WH, Yin MC (2010) Antioxidative and anti-inflammatory protection from carnosine in the striatum of MPTP-treated mice. J Agric Food Chem 58(21):11510–11516. https://doi.org/10.1021/jf103258p
Tsuji Y, Miller LL, Miller SC, Torti SV, Torti FM (1991) Tumor necrosis factor-alpha and interleukin 1-alpha regulate transferrin receptor in human diploid fibroblasts. Relationship to the induction of ferritin heavy chain. J Biol Chem 266(11):7257–7261
Tuomainen TP, Loft S, Nyyssonen K, Punnonen K, Salonen JT, Poulsen HE (2007) Body iron is a contributor to oxidative damage of DNA. Free Radic Res 41(3):324–328. https://doi.org/10.1080/10715760601091642
Velez S, Nair NG, Reddy VP (2008) Transition metal ion binding studies of carnosine and histidine: biologically relevant antioxidants. Colloids Surf B Biointerfaces 66(2):291–294. https://doi.org/10.1016/j.colsurfb.2008.06.012
Vlassara H (2001) The AGE-receptor in the pathogenesis of diabetic complications. Diabetes Metab Res Rev 17(6):436–443
Yan SL, Wu ST, Yin MC, Chen HT, Chen HC (2009) Protective effects from carnosine and histidine on acetaminophen-induced liver injury. J Food Sci 74(8):H259–H265. https://doi.org/10.1111/j.1750-3841.2009.01330.x
Zhang S, Ntasis E, Kabtni S, van den Born J, Navis G, Bakker SJ, Kramer BK, Yard BA, Hauske SJ (2016) Hyperglycemia does not affect iron mediated toxicity of cultured endothelial and renal tubular epithelial cells: influence of l-carnosine. J Diabetes Res 2016:8710432. https://doi.org/10.1155/2016/8710432
Zheng Y, Li XK, Wang Y, Cai L (2008) The role of zinc, copper and iron in the pathogenesis of diabetes and diabetic complications: therapeutic effects by chelators. Hemoglobin 32(1–2):135–145. https://doi.org/10.1080/03630260701727077
Acknowledgements
We thank the volunteers for their participation in the trial. We also thank Professor Wim Derave for performing the carnosinase measurements. This study was supported by the Grant Agency of the Slovak Academy of Sciences VEGA 2/0107/18, Slovak Research and Development Agency SRDA (APVV) 15/0253, Royal Australasian College of Physicians, Diabetes Australia Research Trust and Foundation for High Blood Pressure Research. Carnosine supplement was received from Flamma SPa, Italy. EB is a recipient of the Monash Graduate and Monash International Postgraduate Scholarships. BdC is supported by a National Heart Foundation Future Leader Fellowship (100864).
Author information
Authors and Affiliations
Contributions
EB conceived the study idea, performed data analysis and interpretation, wrote first draft of the manuscript and revised subsequent drafts until publication. BdC designed the study, contributed to the study idea and data analysis, and critically reviewed the manuscript. MPJdC co-designed the study with BdC and contributed to the review of the manuscript. JU & BU assisted with design of the study, performed all clinical measurement and contributed to review of the manuscript. TK & PK contributed to the clinical study performance and data collection. JMFR contributed to review of the manuscript. GA performed to laboratory measurement and reviewed the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare they have no conflict of interest.
Ethical approval
All procedures performed in studies involving human participants were in accordance with the ethical standards of the institutional and/or national research committee and with the 1964 Helsinki declaration and its later amendments or comparable ethical standards.
Informed consent
Written informed consent was obtained from all individual participants included in the study.
Additional information
Handling Editor: S. P. Baba.
Rights and permissions
About this article

Cite this article
Baye, E., Ukropec, J., de Courten, M.P.J. et al. Carnosine supplementation reduces plasma soluble transferrin receptor in healthy overweight or obese individuals: a pilot randomised trial. Amino Acids 51, 73–81 (2019). https://doi.org/10.1007/s00726-018-2623-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00726-018-2623-6