
Abstract
p24 family proteins have been known for a long time, but their functions have remained elusive. However, they are emerging as essential regulators of protein trafficking along the secretory pathway, influencing the composition, structure, and function of different organelles in the pathway, especially the ER and the Golgi apparatus. In addition, they appear to modulate the transport of specific cargos, including GPI-anchored proteins, G-protein-coupled receptors, or K/HDEL ligands. As a consequence, they have been shown to play specific roles in signaling, development, insulin secretion, and the pathogenesis of Alzheimer’s disease. The search of new putative ligands may open the way to discover new functions for this fascinating family of proteins.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
p24 family proteins were first described by Wada et al. (1991). Over the years, and despite their extensive characterization in mammals, yeast, and, more recently, in plants, the functions of these proteins have remained elusive. However, there is no doubt that p24 proteins play important roles in transport along the secretory pathway. The conventional secretory pathway involves the transport of newly synthesized proteins from the ER via the Golgi apparatus to the plasma membrane or the cell surface, but also includes a lateral route from the Golgi apparatus to endosomes and lysosomes (the vacuole in plants).
The so-called early secretory pathway involves bidirectional transport between the ER and the Golgi apparatus and is mediated by coat protein complex I (COPI)-coated and coat protein complex II (COPII)-coated vesicles (Brandizzi and Barlowe 2013). COPII vesicles are formed at specialized regions of the ER, termed ER export sites (ERES), and sequester proteins for transport to the Golgi apparatus, while COPI vesicles are formed at the Golgi apparatus and are involved in intra-Golgi transport and in retrograde transport from the Golgi apparatus to the ER (Brandizzi and Barlowe 2013). p24 proteins have been shown to be efficiently packaged within COPI- and COPII-coated vesicles and to cycle between the compartments of the early secretory pathway. Based on these properties and on their topology, they have long been proposed to function as cargo receptors and in quality control in transport along the secretory pathway. The identification of putative cargos has however been difficult. However, an increasing amount of functional studies have recently become available, which suggest that p24 proteins are key players in the regulation of the secretory pathway and play important specific functions in different organisms.
The p24 family
Phylogeny and nomenclature
The p24 proteins constitute a family of type-I transmembrane proteins of ~24 kDa. Based on sequence homology, p24 proteins can be classified in four subfamilies, named α, β, δ, and γ (Dominguez et al. 1998) (Fig. 1). The number of p24 proteins of each subfamily varies among species (Table 1). The four p24 subfamilies are present in yeast and all animals but plants seem to have members of only p24β and p24δ subfamilies. In general, vertebrates contain ten p24 family members (Schuiki and Volchuk 2012). Phylogenetic analysis of vertebrate p24 proteins showed that the p24α and p24δ subfamilies have a common origin as also appears to be the case for p24β and p24γ subfamilies (for an excellent report on the phylogeny of p24 proteins, see Strating et al. (2009)). The p24α and p24γ subfamilies in most vertebrates have several members while in contrast, the p24β and p24δ subfamilies contain only one single member (Fig. 1). Within each of the four vertebrate p24 subfamilies, the degree of amino acid sequence identity is high, being the p24γ subfamily the one that shows largest variability (Theiler et al. 2014). The yeast, Drosophila and vertebrate p24 subfamilies have expanded independently (Fig. 1). The plant p24δ subfamily seems to have greatly expanded independently from the fungi/animals. In the case of Arabidopsis, p24 proteins of the delta subfamily have been proposed to belong to two different subclasses (which correspond to the two main branches of this subfamily), the δ1 subclass (comprising p24δ3–6) and the δ2 subclass (comprising p24δ7–11) (Chen et al. 2012).

Unrooted tree of the p24 protein family. A multiple alignment of the p24 proteins sequence was constructed using T-Coffee, and the tree was generated from this alignment using the Molecular Evolutionary Genetics Analysis software (MEGA, version 6.06) with the Neighbor-Joining method. The four p24 subfamilies (α, β, γ, and δ) are highlighted by color background shading. Hs Homo sapiens, Mm Mus musculus, Sc Saccharomyces cerevisiae, Dm Drosophila melanogaster, At Arabidopsis thaliana, Os Oryza sativa (see Table 1 for p24 protein nomenclature)
In the past, several nomenclatures for vertebrate p24 proteins have been introduced and they are still being used in parallel (Table 1). p24 proteins were originally named according to their apparent molecular weight in SDS-PAGE (22-24 kDa) (i.e., gp25L, emp24, erv25, p24, p23). As new members were discovered, they were named, by analogy, p26, p27, p28, although their deduced molecular mass was also 22–24 kDa. In the vertebrate protein database, most p24 proteins are annotated as TMED (transmembrane emp24 domain containing protein, emp24 being the first yeast p24 protein identified) (Schimmoller et al. 1995). Dominguez et al. (1998) proposed a more systematic nomenclature involving the use of a Greek letter (α, β, δ, and γ) to identify the subfamily, followed by a number (starting with the first-discovered member). In this review, we have adopted this nomenclature to avoid confusion and to facilitate their identification within the family but since many of the original/unconventional names given to many p24 family members are well-established and they are extensively used in many publications (i.e., p23 in mammals, Erv25 or Emp24 in yeast, or Drosophila members), they have also been used in this article.
Tissue-specific and regulated expression
Most p24 proteins are ubiquitously expressed, although a few of them are expressed in a tissue-specific manner and show regulated expression. These specific expression patterns may reflect specialized functions for these p24 proteins, for instance, the transport of a specific set of cargo proteins.
Strating et al. (2009) showed by RT-PCR that in mouse, eight out of ten p24 transcripts are widely expressed, except for p24α1 and p24γ5 that show restricted expression patterns. Cell-type-specific and selectively induced expression has also been found for some members of the Xenopus p24 family during background color adaptation (Rotter et al. 2002). In Drosophila, the temporal and spatial expression patterns of the p24 genes have been assayed, and it was confirmed that the majority of Drosophila p24 genes are widely expressed and some of them have developmental, tissue-specific, or sex-specific expression and functions (see below) (Boltz et al. 2007). Furthermore, it has been shown that expression of some Drosophila p24 genes is mediated by CrebA/Creb3-like family of bZip transcription factors (Fox et al. 2010).
p24 proteins are highly expressed in secretory cell types such as exocrine, endocrine, and neural cells. Several p24 proteins have been detected in insulinoma cell lines and pancreatic islets and expression levels positively correlated with insulin abundance, suggesting that they are required for insulin biosynthesis and secretion (Hosaka et al. 2007; Zhang and Volchuk 2010; Wang et al. 2012) (see “p24 proteins, insulin secretion, and diabetes”).
In humans, the expression of TMP21 (p24δ1) has been recently studied in detail. It has been shown that the nuclear factor of activated T cells (NFAT) plays a very important role in regulation of human TMP21 and that the expression of TMP21 is at high levels in the heart, liver, lung, kidney, and adrenal gland; moderate levels in the brain, pancreas, prostate gland, testicle, small intestine, colon, stomach, gall bladder, thyroid gland, and trachea; and low levels in the skeletal muscle, skin, and lymphonodus (Liu et al. 2011; Xie et al. 2014). Interestingly, the levels of TMP21 (p24δ1) in the brain decline during postnatal development and reduced levels have been found in individuals with Alzheimer’s disease (Vetrivel et al. 2008) (see “p24 proteins and Alzheimer’s disease”).
In Arabidopsis, five p24 genes (p24δ4, p24δ5, p24δ9, p24β2, and p24β3) have high/medium levels of expression in different tissues, according to public microarray databases. In contrast, p24δ11 has low and flower-specific expression (Zimmermann et al. 2004). The Arabidopsis p24β2 gene is upregulated in response to ER stress (Kamauchi et al. 2005), as has been described for TMED3 (p24γ4) in mammals (Hartley et al. 2010).
Properties and trafficking of p24 proteins
Protein domains
p24 proteins are type I transmembrane proteins, with a short (13–20 residues) cytosolic C-terminal tail, a single transmembrane domain and a large luminal region, including the GOLD domain (see below), a linker region and a coiled-coil domain (Fig. 2a). p24 proteins contain, at the N-terminus, a β-strand-rich globular domain which is also present in several proteins related to Golgi dynamics and secretion (including animal Sec14 proteins, p24 proteins, and Golgi complex-associated protein of 60 kDa (GCP60)-like proteins), and has therefore been named the Golgi dynamics (GOLD) domain (Anantharaman and Aravind 2002). The GOLD domain is predicted to mediate diverse protein-protein interactions (Anantharaman and Aravind 2002) and has been suggested to be involved in cargo recognition, but is also often observed in sugar- and lipid-binding proteins (Gaskell et al. 1995). The GOLD domain of all p24 proteins contains two cysteine residues, suggesting the existence of a disulfide bridge. The alpha-helical coiled-coil domain has been shown to be involved in oligomerization of p24 proteins (Ciufo and Boyd, Emery et al. 2000; Montesinos et al. 2012; Liaunardy-Jopeace et al. 2014). The coiled-coil domains are located at similar positions relative to the transmembrane domains, allowing the interaction between different p24 proteins (Emery et al. 2000) (see below). However, the coiled-coil domain (and not the GOLD domain) has been recently shown to be also important for recognition and transport of GPI-anchored proteins (see “Transport of cargo”) (Theiler et al. 2014).

Properties of p24 proteins. a Domain organization of p24 proteins. The structure of p24 proteins includes two luminal domains, the Golgi dynamics (GOLD) domain and a coiled-coil (CC) domain, separated by a linker region, a single transmembrane (TM) domain and a short cytoplasmic C-terminal tail which contain signals for binding COPI and COPII subunits in the form of a ΦFXXBBXn motif, where Φ is a bulky hydrophobic residue (F/Y/L/I), B is a basic residue and n is ≥2 (see text for details). b Proposed model for a dynamic equilibrium between monomeric, dimeric, and heterotetrameric forms of p24 family members (see text for details)
The transmembrane domain of p24 proteins contains polar residues and a conserved glutamine at the membrane-cytosol interface. Interestingly, the transmembrane domain of p24 (p24β1) has been shown to interact specifically with one single sphingomyelin species, SM18 (Contreras et al. 2012). This interaction depends on a sphingolipid-binding motif, which is also present in several G-protein-coupled receptors (Bjorkholm et al. 2014), putative cargoes of p24 proteins (see “Transport of cargo”), and has been postulated to modulate the equilibrium between monomeric and oligomeric states of p24 proteins (Contreras et al. 2012).
The cytosolic tail of p24 proteins contains signals for binding of COPI and COPII subunits allowing p24 proteins to be efficiently sorted within COPI and COPII vesicles for their bidirectional transport between the ER and the Golgi apparatus (Aniento et al. 2006). Figure 3 shows the amino acid sequence of the cytosolic C-terminus of several members of the different subfamilies. All of them have a conserved phenylalanine residue (often as part of a diaromatic motif) and many of them also contain a dibasic motif, in the form of a ΦFXXBB(X)n motif, where Φ is a bulky hydrophobic residue, B is a basic residue, X can be any amino acid, and n ≥ 2 (Figs. 2a and 3). There are conflicting reports concerning as to which p24 proteins actually bind COPI subunits, the sorting signals involved in this interaction and the COPI subunits which interact with these sorting signals. Nevertheless, it is clear that the dilysine motif present in members of the alpha and delta subfamilies is directly involved in COPI binding (Bethune et al. 2006; Popoff et al. 2011; Jackson et al. 2012; Ma and Goldberg 2013; Gao et al. 2014). On the other hand, the ΦF motif present in all p24 proteins has been shown to bind COPII subunits for their efficient ER export in COPII vesicles (Dominguez et al. 1998; Belden and Barlowe 2001; Barlowe 2003; Contreras et al. 2004b; Aniento et al. 2006). In addition, several reports have also shown that this motif is also involved in binding of COPI subunits, probably in cooperation with the dilysine motif (Fiedler et al. 1996; Sohn et al. 1996; Dominguez et al. 1998; Goldberg 2000; Belden and Barlowe 2001; Contreras et al. 2004b; Aniento et al. 2006). Vertebrate p24 proteins of the alpha subfamily contain a “classical” KKXX motif (Fig. 3). In the case of p25 (p24α2), it has been shown to bind as a dimer to the γ-COP subunit of coatomer and as a monomer to the α/β’/ε-COP B-subcomplex of coatomer (as do other proteins containing KKXX motifs) (Bethune et al. 2006; Popoff et al. 2011; Ma and Goldberg 2013). As a consequence, p25 (p24α2) has the highest affinity for coatomer (when compared with p24 proteins from the other subfamilies) (Bethune et al. 2006). The three yeast members in this subfamily, Erp1p, Erp5p, and Erp6p, have the dilysine motif in the -3,-5 position (Fig. 3), but this KXKXX motif can also bind coatomer very efficiently (Teasdale and Jackson 1996). Therefore, all p24 proteins of the alpha subfamily are expected to be efficiently incorporated in COPI vesicles for their Golgi-to-ER transport. The cytosolic tail of members of the delta subfamily shows significant differences among different organisms (Fig. 3). In mammals, Drosophila, Xenopus, or zebrafish, the dilysine motif is present in the -4,-5 position and the diaromatic motif is in the -8,-9 position. These p24 delta proteins have been shown to bind also COPI subunits (Fiedler et al. 1996, Sohn et al. 1996, Dominguez et al. 1998, Bethune et al. 2006). While it seems that monomers of these proteins have a much lower affinity for coatomer when compared with members of the alpha subfamily, coatomer binding is very similar when they are present as dimers (Bethune et al. 2006). The only member of the delta subfamily in yeast (Erv25p) contains KH instead of the KK motif, but it also binds COPI subunits (Belden and Barlowe 2001). Interestingly, all members of the delta subfamily in plants contain a dilysine motif in the -3,-4 position and a diaromatic motif in the -7,-8 position (Fig. 3), as happens with members of the p24 alpha subfamily (except yeast members). The dilysine motif in plant p24δ proteins has been shown to bind ADP-ribosylation factor 1 (ARF1) and COPI subunits, while the diaromatic motif has been shown to bind COPII subunits but also cooperates with the dilysine motif in COPI binding. As a consequence, plant p24 delta proteins bind COPI with much higher affinity than COPII (Contreras et al. 2004a, b). Members of the p24 beta subfamily lack a dilysine motif but all of them contain a ΦF motif which allows binding of COPII subunits (Dominguez et al. 1998; Belden and Barlowe 2001; Contreras et al. 2004b; Aniento et al. 2006). The C-terminal residues in animal p24 beta proteins is R(K)RVV, while in plants is S(N)RV. Actually, the C-terminal RV motif has been proposed to be involved in the ER export of plant p24 beta proteins (Chen et al. 2012). However, the use of a GFP tag at the C-terminus, just behind the short cytoplasmic tail (which contains the sorting signals for COPI/COPII binding), might have had an influence in the trafficking and localization of these fusion proteins. This is the reason why most XFP-p24 constructs used to investigate p24 protein trafficking (Blum et al. 1996; Majoul et al. 2001; Simpson et al. 2006; Blum and Lepier 2008; Langhans et al. 2008; Montesinos et al. 2012, 2013) have the XFP tag at the luminal N-terminus (immediately behind the signal sequence). The gamma subfamily shows clearly the greatest variability in the sequence of the C-terminal tail, which is consistent with the overall variability in their sequence, but all of them contain a ΦF motif which presumably allows binding of COPII subunits for their ER export.

Amino acid sequence of the cytosolic tail of p24 proteins from the different subfamilies. Classical dilysine residues are shown in red, other basic residues are shown in green and ΦF motifs are shown in blue. The asterisk indicates the position of the conserved phenylalanine residue present in the cytosolic tail of all p24 proteins
Posttranslational modifications and degradation
Several reports have shown differential glycosylation of p24 family members. In rat liver, p24α2 was found to be sensitive to treatment with N-glycosidase F (which cleaves all N-linked glycans at the site of attachment to asparagine) and neuraminidase (which removes sialic acid residues), but Endo H resistant, suggesting that p24α2 is modified by glycosylation enzymes of the medial- and trans-Golgi to acquire complex oligosaccharides, including terminal ones in the form of sialic acid (Dominguez et al. 1998). In another study, the glycosylation of different p24 family members was compared in HeLa cells, showing that p24γ3 (gp27) and p24α2 (GMP25) were glycosylated, in contrast to p23 (p24δ1), p24 (p24β1), and p26 (p24γ4) (Fullekrug et al. 1999). p24γ3 acquired complex oligosaccharides and sialic acid and became Endo H resistant, which indicates that p24γ3 is also modified by glycosylation enzymes of the medial- and trans-Golgi. However, p24α2 remained Endo H sensitive in HeLa cells (Fullekrug et al. 1999), in contrast with the situation in rat liver, where p24α2 was found to be EndoH resistant (Dominguez et al. 1998). More recently, as shown by sensitivity to N-glycosidase F, p24α2 in human embryo kidney (HEK) cells has also been found to be glycosylated, and it was Endo H resistant, as in rat liver. In addition, p24α2 has been postulated to be phosphorylated (Liu et al. 2015). Tmp21 (p24δ1) extracted from chondrocytes was found to be sialylated (Osiecka-Iwan et al. 2014), in contrast to the situation in HeLa cells, where p24δ1 is not glycosylated (Fullekrug et al. 1999). This is the first report of a tissue-specific distribution of a sialylated p24 protein. A novel p24δ isoform, Rrt6, has recently been identified and is the only S. cerevisiae p24 protein which is inducibly expressed and receives N-linked glycosylation (Hirata et al. 2013). To our knowledge, there are no reports on the glycosylation of p24 proteins in plant cells, although Arabidopsis members of the p24δ1 subclass (p24δ3-p24δ6) contain a putative N-glycosylation motif which is not present in members of the p24δ2 subclass (p24δ7-p24δ11).
There are only a couple of reports on the mechanisms involved in the degradation of p24 proteins. Liu et al. (2008) found that TMP21/p23 (p24δ1) has a short half-life of around 3 h and is degraded by the ubiquitin-proteasome pathway, as it is also the case for the other components of the presenilin-associated complex (see below). In contrast to TMP21, gp27 (p24γ3) was found to have a long half-life (Fullekrug et al. 1999). Recently, the stability of p24 proteins of the beta subfamily in plants was investigated. Transient expression experiments showed that coexpression with p24δ5 increased enormously the stability of p24β2 (as compared with p24β2 expression in the absence of p24δ5), possibly because p24δ5 holds p24β2 in the early secretory pathway (Montesinos et al. 2012). In addition, it was found that the protein levels of p24β2 and p24β3 were insensitive to treatment with the proteasomal inhibitor MG-132, but increased in a very significant manner upon treatment with E-64, an inhibitor of cysteine proteases. Therefore, it was speculated that the low stability of these proteins when expressed alone may be a consequence of increased degradation by cysteine proteases upon their transport to the prevacuolar compartment and the vacuole (Montesinos et al. 2013).
Oligomerization of p24 proteins
p24 proteins can interact with each other through their coiled-coil domains (Ciufo and Boyd, Emery et al. 2000; Montesinos et al. 2012; Liaunardy-Jopeace et al. 2014) to form different types of oligomeric complexes, which are important not only for their trafficking and localization but also for their stability. However, the actual composition of these putative complexes is still controversial. Several approaches have been used to characterize putative p24 complexes. Early experiments using sedimentation density gradient centrifugation already suggested that p24 proteins may exist as large complexes (Dominguez et al. 1998).
Coimmunoprecipitation experiments have been used to demonstrate the interaction between p23 (p24δ1) and p24 (p24β1) in CHO cells (Gommel et al. 1999). In yeast, immunoprecipitates of Erp1p (p24α) were found to contain also Erp2p (p24γ), Emp24p (p24β), and Erv25p (p24δ), suggesting that these proteins form a heterotetrameric complex, containing members of the four p24 subfamilies, which was named the yeast p24 complex (Marzioch et al. 1999). In parallel, immunoprecipitates of gp27 (p24γ3) in HeLa cells were found to contain GMP25 (p24α2), p24 (p24β1), and p23 (p24δ1), suggesting that p24 proteins in animal cells also form heterotetramers including members of the four p24 subfamilies (Fullekrug et al. 1999). Pull-down experiments have also been used to demonstrate the interaction of p23 (p24δ1) with p24 (p24β1), p25 (p24α2), and p28 (p24γ2) in CHO cells (Fujita et al. 2011). In contrast, pull-down experiments in HeLa cells showed that p28 (p24γ2) interacts with p23 (p24δ1) and p25 (p24α2) but not with p24 (p24β1) (Koegler et al. 2010). More recent pull-down experiments in yeast showed the interaction of the three p24γ isoforms with Emp24p (p24β), Erv25p (p24δ) and with different p24α isoforms, suggesting that yeast p24 complexes should contain Emp24p (p24β), Erv25p (p24δ) and different combinations of p24α and γ isoforms (Hirata et al. 2013).
Another approach to investigate the composition of putative p24 complexes is to investigate how the deletion of one p24 family member affects the stability of other p24 family members. In this respect, experiments using yeast mutants were used to show that the levels of Emp24p (p24β) and Erv25p (p24δ) were interdependent (Belden and Barlowe 1996). This type of approach has also been widely used in animal cells. In mouse, a p23 (p24δ1) heterozygous mutant showed a reduction in the protein levels of p25 (p24α2) and p26 (p24γ4) (Denzel et al. 2000). p23 silencing in N2a neuroblastoma and HeLa cells also caused reduced protein levels of p24 (p24β1), p25 (p24α2), and tp24 (p24γ1) (Vetrivel et al. 2007). In a mammalian cell line that stably expresses inducible temperature-sensitive GPI-fused proteins, p23 (p24δ1) silencing caused a parallel decrease in the levels of p24 (p24β1) (Takida et al. 2008). Protein levels of TMED9 (p24α2), TMED7 (p24γ3), and TMED10 (p24δ1) were reduced in a mouse mutant line lacking TMED2 (p24β1) (Jerome-Majewska et al. 2010). Knocking down p24δ1 in insulinoma cell lines also reduced the endogenous levels of both p24β1 and p24α2 (Zhang and Volchuk 2010). Knockdown of p23 (p24δ1) in Chinese hamster ovary (CHO) cells caused a reduction in protein levels of p25 (p24α2), p24 (p24β1), and p28 (p24γ2) (Fujita et al. 2011). In CHO cell lines, knockdown of p24δ1 caused a reduction in protein levels of p24α2, p24β1, and p24γ2, suggesting that a complex may be formed between these proteins. However, the levels of other p24γ isoforms (p24γ1, p24γ3, and p24γ4) were also reduced upon p24δ1 knockdown, suggesting that different p24γ subunits may be part of p24 complexes containing p24α, p24β and p24δ subunits (Theiler et al. 2014). Altogether, these experiments suggest that p24 proteins form heterotetrameric complexes including members of the four p24 subfamilies, with the gamma subunit being the most variable component of these complexes. In contrast, knockdown of p28 (p24γ2) in HeLa and HepG2 cells caused reduced levels of p23 (p24δ1) and p25 (p24α2), but not of p24 (p24β1) (Koegler et al. 2010). Interestingly, in Drosophila, knockdown of a single p24 protein caused both reduction and upregulation of several p24 transcripts (Buechling et al. 2011).
A number of reports argue against p24 proteins forming stable heterotetramers. Gel filtration experiments in yeast showed that a significant proportion of Erp1p (p24α), Erp2p (p24γ), Emp24p (p24β), and Erv25p (p24δ) coeluted at a molecular mass of ≈100 kDa, consistent with these proteins forming a heterotetramer (Marzioch et al. 1999), but each of the four proteins was also found to elute at a lower-molecular-mass, mostly consistent with p24 dimers. Indeed, in the absence of Erp1p (p24α) and Erp2p (p24γ), most Emp24p (p24β) and Erv25p (p24δ) were present as dimers, suggesting that these two proteins can form stable complexes in the absence of the two other p24 family members (Marzioch et al. 1999). Gel-filtration experiments in HeLa cells suggested that p24 proteins exist mostly as monomers and dimers of different composition depending on their subcellular localization (Jenne et al. 2002). Therefore, the available data could be consistent with a model where p24 proteins do not exist in a stable complex all of the time. Instead, it is more likely that they exist in a dynamic equilibrium between complexes and individual proteins (Fig. 2b), and perhaps the postulated heterotetramer could be a dimer of dimers, as proposed by Ciufo and Boyd (2000). Actually, dimers composed of p24β and p24δ proteins have been extensively characterized. In this respect, yeast Erv25p (p24δ) and Emp24p (p24β) can be efficiently incorporated into COPII vesicles for ER exit without the presence of Erp1p (p24α) and Erp2p (p24γ) (Belden and Barlowe 1996, 2001), while in animals, p23 (p24δ1) and p24 (p24β1) can form a complex which is transported from the ER to the Golgi apparatus (Gommel et al. 1999, Emery et al. 2000). In both cases, the dilysine motif present in the p24δ member (Erv25p or p23) was postulated to be responsible for retrograde Golgi-to-ER transport of both complexes (Gommel et al. 1999, Belden and Barlowe 2001). Indeed, p24β/δ dimers appear to be part of the molecular mechanism involved in the biogenesis of COPI vesicles (as described in “Biogenesis of COPI vesicles”).
It is not clear which is the contribution of the α/γ subunits to the function of putative p24 complexes. In this respect, Emp24p (p24β) and Erv25p (p24δ) have long been considered as the constitutive members of the yeast p24 complex (Marzioch et al. 1999) and to be responsible for most cellular p24 activity, as tested by their capacity to prevent secretion of the ER resident Kar2 (Belden and Barlowe 2001). However, it has been recently proposed that p24α and p24γ subunits in yeast are also essential for p24 function and that active p24 complexes should contain at least one each of p24α and p24γ isoforms as well as the invariant p24β and p24δ subunits (Hirata et al. 2013). More recently, p24 proteins in mammals have been proposed to form different hetero-oligomeric complexes differing in their p24γ subunit whereby the composition of these complexes appeared to determine the cargo specificity (Theiler et al. 2014). Indeed, p24γ2, but not other p24γ subunits, was found to be involved in transport of GPI-APs and the interaction between GPI-APs and p24γ2 involved the α-helical region in p24γ2.
Plants contain only members of the beta and delta subfamilies, and therefore, any complex should be made with only these two types of p24 proteins. We have characterized the protein levels of different p24 family members in mutants lacking a single p24 protein (Montesinos et al. 2012, 2013). The p24δ5 or p24δ4 (p24δ1 subclass) mutants had similar levels of p24δ5 or p24δ4, respectively, but both had reduced levels of p24δ9 (p24δ2 subclass), p24β2, and p24β3. Interestingly, the p24δ10 (p24δ2 subclass) mutant showed increased protein levels of the closely related p24δ9 (δ2 subclass), probably induced in the absence of p24δ10, and no decrease in the levels of p24δ5 (δ1 subclass) or p24β proteins. This indicates that the increased proteins levels of p24δ9 may compensate for the lack of p24δ10 in the p24δ10 mutant. Silencing of p24β2 or p24β3 caused reduced proteins levels of p24β3 or p24β2, respectively, and reduced levels of p24δ5 and p24δ9. These results suggest an interdependence in the protein levels of p24δ proteins from the two subclasses and the two members of the p24β subfamily. On the other hand, coimmunoprecipitation and pull-down experiments also suggested the existence of a direct interaction between p24δ5 (p24δ1 subclass), p24δ9 (p24δ2 subclass), p24β2, and p24β3 (Montesinos et al. 2012, 2013).
p24 trafficking and localization
p24 proteins have been shown to localize to the compartments of the early secretory pathway, including the endoplasmic reticulum (ER), the ER–Golgi intermediate compartment (ERGIC), the cis-Golgi network (CGN), or the Golgi apparatus (Stamnes et al. 1995; Belden and Barlowe 1996; Blum et al. 1996; Sohn et al. 1996; Nickel et al. 1997; Rojo et al. 1997; Dominguez et al. 1998; Fullekrug et al. 1999; Gommel et al. 1999; Emery et al. 2000; Rojo et al. 2000). They are also major constituents of both coat protein I (COPI)-coated (Stamnes et al. 1995; Sohn et al. 1996; Gommel et al. 1999) and COPII-coated (Schimmoller et al. 1995; Belden and Barlowe 1996) vesicles, which facilitate their bidirectional transport between the ER and the Golgi apparatus. In Arabidopsis, immunogold labeling on cryosections showed that p24 proteins of the delta subfamily (including p24δ4, p24δ5, and p24δ9) localized mainly to ER membranes, although some labeling was also found at the cis-Golgi. In contrast, p24 proteins of the beta subfamily (p24β2 and p24β3) localized mainly to the Golgi apparatus, with some labeling also at ER membranes (Montesinos et al. 2012, 2013).
p24 proteins have also been found to localize to membranes different from the ER–Golgi interface, including peroxisomes (Marelli et al. 2004), secretory granules (Hosaka et al. 2007), and even to the plasma membrane (Chen et al. 2006; Blum and Lepier 2008; Langhans et al. 2008), and to modulate transport of G-protein-coupled receptors or Toll-like receptors to the plasma membrane (see “Transport of cargo”).
The relative contribution of the C-terminal tail and the luminal domains to trafficking of p24 proteins is controversial. The localization of a p23 (p24δ1)/p24 (p24β1) dimer was shown to be independent of the KKLIE motif in the cytosolic tail of p23 but instead required the coiled-coil domains in both proteins (Emery et al. 2000). This is consistent with the role of the CC domain in allowing oligomerization of p24 proteins, which has a strong influence in their trafficking. However, Blum and Lepier (2008) showed that the minimal requirement for cycling of p23 in the early secretory pathway was the transmembrane domain and the cytoplasmic tail with the KKLIE motif. In contrast, the luminal domain in p23 was not necessary for ER–Golgi transport but was required for post-Golgi transport of p23 to the plasma membrane. The Golgi localization of TMED7 (p24γ3) has been shown recently to require its C-terminal tail and was proposed to depend on its diphenylalanine motif. However, none of the deletion mutants used in this report contained only the transmembrane domain and the cytosolic tail (with or without the FF motif) (Liaunardy-Jopeace et al. 2014).
In Arabidopsis, the cytosolic tail in p24δ5, containing a KK motif, was found to be necessary and sufficient for its trafficking between ER and Golgi, since a deletion mutant containing the transmembrane domain and the cytosolic tail (and thus lacking the luminal coiled-coil and GOLD domains) was able to cycle between these compartments and to localize at the ER at steady state (Montesinos et al. 2012). In contrast, the coiled-coil domain, which was shown to be required for its interaction with p24β2, was also required for post-Golgi trafficking of p24δ5 to the vacuole (Montesinos et al. 2012). We have proposed that hetero-oligomeric complexes between Arabidopsis p24 proteins from the p24β and p24δ subfamilies may cycle in the early secretory pathway, although the stoichiometry and composition of these complexes remains to be established. However, by a combination of biochemical approaches and coexpression experiments, we have proposed that “anterograde” (ER–Golgi) complexes should include p24β2, which has been shown to facilitate transport of both p24δ5 (δ1 subclass) and p24δ9 (δ2 subclass) to the Golgi apparatus. p24β3 may bind to complexes containing p24β2/p24δ5 or p24β2/p24δ9 for its transport to the Golgi apparatus. On the other hand, “retrograde” (Golgi–ER) complexes should contain p24δ proteins (for sorting into COPI vesicles), probably including members from the δ1 and δ2 subclasses. p24β3 may be recycled to the ER in complexes containing both p24δ5 and p24δ9 (Montesinos et al. 2012, 2013).
Functions of p24 proteins
In yeast, p24 proteins do not seem to be essential for vesicular transport, since a mutant strain lacking all p24 members showed only mild phenotypic alterations (Springer et al. 2000), including defects in the transport of GPI-APs (see below). In contrast, a homozygous knockout of p23 (p24δ1) in mice is lethal in an early embryonic stage (Denzel et al. 2000). In yeast, deletion of p24 proteins activates the UPR (Belden and Barlowe 2001), which may help to alleviate the transport defects caused by the loss of p24 function (Aguilera-Romero et al. 2008). In particular, the UPR causes the upregulation of several genes involved in Golgi-to-ER retrograde transport, including genes coding for coatomer subunits and the K/HDEL receptor ERD2 (Travers et al. 2000). Loss of p24 function in Drosophila also activates an ER stress response (Boltz and Carney 2008). However, it is possible that activation of such ER stress responses in higher eukaryotes is not sufficient to compensate for the loss of p24 function.
In this section, we will summarize the main functions which have been attributed to p24 proteins, including basic aspects of membrane trafficking but also more specific functions in different cell types.
Biogenesis of COPI vesicles
Over the years, a wealth of reports indicate that p24 proteins are key molecular players in the formation of COPI vesicles from Golgi membranes (for reviews, see (Popoff et al. 2011; Jackson 2014)). First, p23 (p24δ1) and p24 (p24β1) have been shown to be highly enriched in Golgi-derived COPI vesicles, in amounts stoichiometric to ADP-ribosylation factor 1 (ARF1), the small GTPase involved in the formation of COPI vesicles, and coatomer (Sohn et al. 1996, Gommel et al. 1999). In vitro, budding of COPI vesicles from liposomes with a composition resembling Golgi membranes was shown to require coatomer, ARF1, GTP, and the cytoplasmic tail of p23 (p24δ1) (Bremser et al. 1999). Using an in vitro Golgi vesicle budding assay, it has been shown that p24 proteins are also required for COPI vesicle formation in yeast (Aguilera-Romero et al. 2008). The cytoplasmic tail of p23 (p24δ1) has been shown to interact with the GDP-bound form of ARF1, leading to the proposal that p23 (p24δ1) acts as the ARF1 receptor (Gommel et al. 1999). However, FRET experiments showed that both p24a (p24β1) and p23 (p24δ1) interact with ARF1 (Majoul et al. 2001).
As mentioned before, the cytosolic tail of p24 proteins also have the ability to interact with coatomer (Harter et al. 1996; Bethune et al. 2006; Dominguez et al. 1998; Contreras et al. 2004a). The coatomer complex is composed of seven subunits (α/β/β’/γ/δ/ε/ζ), which are recruited en bloc onto Golgi membranes (Hara-Kuge et al. 1994). However, it is believed to include two subcomplexes, the outer layer or B-subcomplex (α/β’/ε) and the inner layer or F-subcomplex (β/γ/δ/ζ) (Jackson 2014). All p24 proteins have been proposed to bind as dimers to two independent sites in the γ-COP subunit, while p25 (and perhaps other p24 proteins containing a classical dilysine motif), can also bind as a monomer to the B-subcomplex (Bethune et al. 2006). Binding of p23 to the gamma COP subunit has been proposed to cause a conformational change that is transmitted to the alpha subunit leading to coatomer polymerization (Reinhard et al. 1999; Langer et al. 2008). The cytosolic tail of p24 (p24β1)—but not that of p23—has also been shown to interact with ARF GTPase-activating protein (GAP)1 and cause a direct inhibition of ARF-GAP1-mediated GTP hydrolysis on ARF1, thus preventing premature uncoating and allowing cargo selection within COPI vesicles to take place (Goldberg 2000; Lanoix et al. 2001; Majoul et al. 2001).
Altogether, these results are consistent with the following model for the participation of p24 proteins in COPI vesicle formation (Popoff et al. 2011; Jackson 2014) (Fig. 4). Recruitment of ARF1 to membranes involves a direct interaction of ARF1-GDP with dimers of p24 proteins (probably p24β/p24δ dimers). Upon recruitment of ARF-GEFs, ARF1 is activated by GDP/GTP exchange. This causes a conformational change in ARF1, which exposes its N-terminal amphipathic and myristoylated helix allowing membrane insertion of ARF1-GTP and its dissociation from p24 proteins (Franco et al. 1996, Antonny et al. 1997). p24 proteins can then interact with coatomer (either to the γ-subunit of the F-subcomplex or to the B-subcomplex), which also interacts with ARF1-GTP. In particular, ARF1-GTP has been shown to interact with the F-subcomplex of coatomer, which has two binding sites for ARF1-GTP (Yu et al. 2012; Jackson 2014). p24α and p24γ proteins can also be recruited at this stage, perhaps forming a tetramer with p24β/p24δ dimers. Interaction with p24 proteins leads to coatomer polymerization and the formation of COPI vesicles (Popoff et al. 2011). p24 proteins also control coat depolymerization inhibiting GTP hydrolysis in ARF1, which is a prerrequisite for uncoating (Goldberg 2000, Lanoix et al. 2001). p24 proteins can also interact with the K/HDEL receptor ERD2, which is efficiently incorporated within COPI vesicles and has also been proposed to be involved in a variety of interactions which contribute to COPI vesicle formation (Majoul et al. 2001; Montesinos et al. 2014). These interactions are triggered upon binding of the KDEL ligand to ERD2 which takes place at the slightly acidic pH of the cis-Golgi (Paroutis et al. 2004; Martiniere et al. 2013; Shen et al. 2013). Ligand binding induces ERD2 oligomerization (Majoul et al. 2001) and facilitates the interaction between ERD2 and ARF-GAP in vivo (Majoul et al. 2001), previously shown biochemically (Aoe et al. 1997, 1998), as well as the interaction between ERD2 and ARF1 or between ARF1 and ARF-GAP (Aoe et al. 1997; Majoul et al. 2001). ERD2 also interacts with coatomer, thus contributing to the formation of COPI vesicles enriched in ERD2 and KDEL ligands (Majoul et al. 2001) (Fig. 4). The interaction between ERD2 and coatomer involves a KKXSXXX motif which is only active when serine-209 is phosphorylated by protein kinase A (Cabrera et al. 2003). Such a phosphorylation is likely to occur at the Golgi apparatus, although this has not yet been demonstrated.

p24 proteins and COPI vesicle formation. p24 proteins are involved in a number of events important for COPI vesicle formation, including the recruitment of ARF1 to the Golgi membrane (1), the interaction with the K/HDEL receptor ERD2 (2) and the interaction with coatomer, which also binds to ARF1, ERD2 and other dilysine cargo (3). These array of interactions facilitates coatomer polymerization and the formation of COPI vesicles (4)
Arabidopsis p24δ5 and p24δ9 have also been shown to interact with both ARF1-GDP and coatomer. In the case of p24δ5, this interaction involves both the dilysine motif in the -3,-4 position and the diphenylalanine motif in the -7,-8 position, acting cooperatively. This suggests that all Arabidopsis p24 proteins of the delta subfamily have the ability to recruit ARF1 and coatomer and be involved in the formation of COPI vesicles (Contreras et al. 2004a; Montesinos et al. 2012, 2014). In addition, we have shown recently that Arabidopsis p24δ5 and p24δ9 (but not p24β2) interact with ERD2, thus contributing to sorting of ERD2 within COPI vesicles and retrieval of K/HDEL ligands (Montesinos et al. 2014) (see below).
p24 proteins may also play a role in the formation of COPII vesicles, although this has not yet been firmly established. It is known that p24 proteins are enriched in COPII vesicles (Schimmoller et al. 1995; Belden and Barlowe 1996) and their cytoplasmic tails contain signals for binding of COPII subunits, in particular the Sec23/Sec24 subunits, thus allowing their efficient sorting within COPII vesicles (Dominguez et al. 1998; Miller et al. 2003; Contreras et al. 2004b). However, COPII vesicles can still form in the absence of p24 proteins in vitro (Matsuoka et al. 1998), although they are devoid of specific cargos, which emphasize the putative role of p24 proteins as cargo receptors, which is discussed in the following section. On the other hand, p24 proteins have been proposed to influence the formation of COPII vesicles by altering the physical properties of the ER membrane due to their abundance and asymmetric localization (the bulk of the protein residing predominantly in the ER lumen) (Copic et al. 2012). This would also be the case for GPI-anchored proteins, which are well-characterized cargos of p24 proteins (as discussed in the following section), and which reside exclusively in the ER lumen. A very recent report suggests that asymmetrically distributed membrane proteins, such as p24 proteins, require the scaffolding function of the cargo adaptor Lst1p (a Sec24p homolog) to form larger COPII vesicles which can then accommodate “difficult” cargo proteins (D'Arcangelo et al. 2015).
Transport of cargo
p24 proteins have long been proposed to function as cargo receptors for transport along the secretory pathway. However, in contrast to many classical cargo receptors, which are involved in the transport of soluble proteins, most putative p24 cargos described so far appear to be lipid-linked proteins or membrane proteins. On the other hand, secretion of invertase, a soluble secreted protein, was found to be delayed in emp24 and erv25 yeast mutants (Schimmoller et al. 1995; Belden and Barlowe 1996; Marzioch et al. 1999), but not in other p24 deletion mutants (Marzioch et al. 1999). In this section, we describe the best characterized putative cargos of p24 proteins.
GPI-anchored proteins
Early studies in yeast already suggested a role of p24 proteins in the transport of glycosylphosphatidylinositol (GPI)-anchored proteins (GPI-APs). In particular, ER-to-Golgi transport of Gas1p (a GPI-AP) was found to require Emp24p (p24β) (Schimmoller et al. 1995) and Erv25p (p24δ) (Belden and Barlowe 1996), but also Erp1p (p24α) and Erp2p (p24γ), the other members of the putative p24 complex in yeast (Marzioch et al. 1999). Indeed, p24 proteins were shown to interact directly with Gas1p facilitating their inclusion in ER-derived COPII vesicles (Muniz et al. 2000). In yeast, GPI-anchored proteins and other secretory proteins have been proposed to exit the ER in different types of COPII vesicles. Using in vitro assays to reconstitute the formation of ER-derived vesicles, the GPI-anchored proteins Gas1p and Yps1p were found in vesicles different from those containing other secretory proteins (like the amino acid permease Gap1p, glycosylated pro-α-factor, or alkaline phosphatase) (Muniz et al. 2000). In yeast, cargo concentration at ERES can be visualized using a temperature-sensitive sec31-1 (COPII) mutant, which blocks ER exit. Using this approach, GPI-anchored proteins were found to concentrate in specific ERES different from those containing other secretory proteins (e.g., glycosylated pro-α-factor or the hexose transporter Hxt1p), and exit the ER in a process that required the p24 protein Emp24p (Castillon et al. 2011). Emp24p could be specifically cross-linked to several GPI-anchored proteins (Gas1p, Yps1p, Ccw14-GFP, Cwp2-GFP) but not to two unrelated transmembrane proteins, Gap1p or Hxt1p (Castillon et al. 2011).
Using the sec31-1 mutant, it was also found that the p24 complex is not required to concentrate GPI-APs into ERES. In addition, ER exit of GPI-APs was shown to involve a specific interaction with the specialized COPII isoform Lst1p (Castillon et al. 2011). Based on these results, it has been proposed that the yeast p24 complex, rather than participating in sorting of GPI-APs within COPII vesicles, may instead function as an adaptor connecting remodeled GPI-APs with the COPII coat to facilitate their incorporation within COPII vesicles. Indeed, a very recent report suggests that binding of remodeled cargo induces the p24 complex to specifically recruit the COPII subunit Lst1p, for the specific ER exit of GPI-APs (Manzano-Lopez et al. 2015). In addition, the p24 complex may also contribute to retrograde Golgi-to-ER transport of unremodeled GPI-APs (Castillon et al. 2011).
p24 proteins have also been shown to participate in efficient ER-to-Golgi transport of GPI-APs in mammals (Takida et al. 2008, Bonnon et al. 2010). In Chinese hamster ovary (CHO) cells, silencing of p23 (p24δ1) caused a significant delay in transport to the plasma membrane of vesicular stomatitis virus G protein (VSVG) with a GPI-anchor but not that of non-GPI-linked VSVG (Takida et al. 2008). In addition, p23 knockdown caused a delayed maturation of decay-accelerating factor (DAF), a well-characterized GPI-anchored protein in human cells, suggesting that p23 (p24δ1) is involved in ER–Golgi transport of GPI-anchored proteins in human cells (Takida et al. 2008). In HeLa cells, p23 (p24δ1) or p24 (p24β1) silencing (but not p25 silencing) caused a significant inhibition of ER-to-Golgi transport of the human GPI-anchored protein CD59 or the folate receptor alpha, a GPI-anchored protein carrying three N-glycans, but was without effect in the transport of the human transferrin receptor, a type I transmembrane protein which also carries three N-glycans, suggesting a selective function of p23 (p24δ1) and p24 (p24β1) in ER-to-Golgi transport of human GPI-APs (Bonnon et al. 2010). ER-to-Golgi transport of the GPI-AP CD59 was found to be selective (rather than bulk-flow) and COPII-dependent and required specifically the Sec24 isoforms Sec24C/D, but not Sec24A/B. p24 (p24β1) and p23 (p24δ1) were found to directly interact with GPI-anchored proteins and a fraction of p24 and p23 was found to copartition with CD59 in lipid rafts (Bonnon et al. 2010). In addition, cholesterol depletion caused the accumulation of both CD59 and p24 proteins (but not ERGIC-53) in the ER (Bonnon et al. 2010). These results constituted the first evidence that in mammals there are two different types of COPII vesicles for ER-to-Golgi transport and that GPI-anchored proteins may form at specific ERES, perhaps with a specific lipid composition. Sorting of GPI-APs to ERES has been shown to require proper structural remodeling and involves association with p23 (p24δ1) and p24 (p24β1) for efficient ER exit of GPI-APs (Fujita et al. 2011). Sorting of GPI-APs to COPII vesicles was impaired in p23 (p24δ1) knockdown cells, suggesting that concentration of GPI-APs at ERES and packaging into COPII vesicles is dependent upon p24 proteins, in contrast to the situation in yeast.
Binding of GPI-anchored proteins (as binding to K/HDEL cargos to the K/HDEL receptor; see below) has been shown to be pH-dependent, being maximal at neutral pH and much lower at slightly acidic pH (Fujita et al. 2011). This is consistent with a model where p24 proteins bind correctly remodeled GPI-anchored proteins at the neutral pH of the ER and dissociates them at the mildly acidic pH of the Golgi apparatus. This way, GPI-APs can be transported to the cell surface while p24 complexes can be recycled to the ER in COPI vesicles (Fujita et al. 2011). Recently, it has been shown that specific binding of p24γ2 to GPI-APs involves its α-helical region, but not the GOLD domain (Theiler et al. 2014).
Wnt proteins
Wnt proteins are secreted, lipid-modified glycoproteins that control animal development. Wnts are cotranslationally imported into the ER lumen where they are palmitoylated at conserved cysteine residues by Porcupine, a membrane bound O-acyltransferase (Tanaka et al. 2000, Gross and Boutros 2013). In Drosophila, p24 family members were tested for a function in secretion of Wingless (Wg), a Wnt Drosophila protein. Two of them, Éclair (p24α) and Emp24 (p24β) were found to be necessary for ER export and secretion of Wg (Port et al. 2011). However, only Emp24 (and not Éclair) was found to directly interact with Wg. In another report, ER export of Wg and Wnt inhibitor of Dorsal (WntD) was shown to require Opossum (Opm), a conserved member of the p24γ subfamily and two other Drosophila p24 proteins, CHOp24 (Emp24, p24β) and p24-1 (p24γ). Interestingly, Opm was found to interact with Wingless but not with WntD, which is not palmitoylated as other Wnts (Buechling et al. 2011). p24 proteins have also been shown to be required for transport of Wnt proteins in mammalian cells, as shown by knockdown of TMED5 (p24γ2), a human homologue of Opm, in HEK293T cells (Buechling et al. 2011). The Drosophila p24δ protein Baiser has also recently been shown to interact also with Wg (as CHOp24, but in contrast to Éclair) and to colocalize with Wg and the ER v-SNARE Sec22, suggesting that Baiser may be involved in packaging Wg in COPII vesicles (Li et al. 2015).
Both Wnt ligands and GPI-APs are anchored to the luminal leaflet of the ER membrane and therefore cannot directly associate with cytosolic COPII proteins. By associating with p24 proteins, these lipid-linked cargos can be selectively incorporated within COPII vesicles. In this respect, it has been proposed that GPI-APs and Wnt proteins may use similar p24 complexes (probably containing p24γ2) for their ER-to-Golgi transport (Kinoshita et al. 2013).
G-protein-coupled receptors (GPCRs)
PAR-2, a protease-activated receptor, was the first cargo protein shown to be recognized by p24 proteins (in particular p24β1 and p24δ1) in mammalian systems. Protease-activated receptors (including PAR-1 and PAR-2) are G-protein-coupled receptors which are activated by irreversible proteolytic cleavage by serine proteases (for a review, see (Zhao et al. 2014)). Upon activation, PAR-2 is rapidly sorted to early endosomes and then transported to the lysosomes for degradation. PAR-2 resensitization involves repopulation of cell-surface receptors from presynthesized and newly synthesized PAR-2 molecules stored in the Golgi apparatus (Zhao et al. 2014). Transport of PAR-2 molecules from the Golgi to the plasma membrane has been shown to be controlled by p24A (p24β1) and p23 (p24δ1) (Luo et al. 2007, 2011). By biochemical approaches, p24A (p24β1) was shown to interact with PAR-2 in HEK293 cells, rat astrocytes, and rat cortical neurons (Luo et al. 2007). This interaction involved the second extracellular loop of PAR-2 and the N-terminal region of p24A (p24β1), including the GOLD domain but also the linker region. Actually, the GOLD domain was found to be necessary but not sufficient for the interaction with PAR-2, although it is possible that the linker region is simply needed for a proper folding of the GOLD domain (Luo et al. 2007). Colocalization experiments showed that the interaction between p24A or p23 and PAR-2 takes place at the Golgi apparatus (Luo et al. 2007, 2011). Activation of PAR-2 (with a specific peptide agonist) reduced significantly the interaction with p24A, suggesting that p24A dissociates from PAR-2 after receptor activation. PAR-2 activation was also shown to activate ARF1. Based on these results, a model has been proposed where p24A (p24β1) holds PAR-2 at the Golgi apparatus in unstimulated cells. Upon PAR-2 activation, ARF1-GDP can interact with p24A and be recruited to Golgi membranes. GTP/GDP exchange in ARF1 (catalyzed by a brefeldin A-sensitive ARF1-GEF) leads to dissociation of p24A (p24β1) and PAR-2, allowing transport of PAR-2 to the plasma membrane (Luo et al. 2007). p24A (p24β1) has been shown to bind several other GPCRs, including PAR-1, but also several nucleotide receptors as well as a μ-opiod receptor (Luo et al. 2011). Interestingly, both p24 (p24β1) and several 7TM GPCRs contain sphingolipid-binding motifs suggesting a regulation of these proteins by sphingolipids (Bjorkholm et al. 2014).
Toll-like receptors
Toll-like receptor 4 (TLR-4), the receptor for the bacterial product endotoxin, is a key regulator of innate immunity and inflammation (Bryant et al. 2010). TLR-4 functions at the plasma membrane, where it is activated by the glycolipid lipopolysaccharide (LPS) and internalized into early endosomes. However, trafficking of TLR-4 in the secretory pathway is less well characterized. The p24 family member TMED7 (p24γ3) has been shown to form dimers or oligomers (via its coiled-coil domain) which are able to interact with TLR-4 (Liaunardy-Jopeace et al. 2014). Overexpresion of TMED7 caused increased cell surface expression of TLR-4 while silencing of TMED7 produced the opposite effect, suggesting that TMED7 is required for trafficking of TLR-4 to the cell surface. However, it is not clear whether TMED7 is directly involved in ER exit of TLR-4 and how both proteins dissociate to allow transport of TLR-4 to the cell surface (Liaunardy-Jopeace et al. 2014; Liaunardy-Jopeace and Gay 2014).
p24 proteins and ER retrieval of K/HDEL ligands
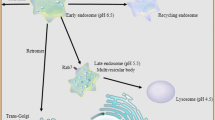
p24 delta proteins have been shown to be involved in Golgi-to-ER transport of K/HDEL ligands bound to the K/HDEL receptor ERD2 via a direct interaction between p24 delta proteins and ERD2 (Majoul et al. 2001; Montesinos et al. 2014). FRET experiments in animal cells showed that the interaction between p23 (p24δ1) and ERD2 takes place at the Golgi complex. Once the KDEL ligand reaches the ER, p23 and ERD2 no longer interact (Majoul et al. 2001). In Arabidopsis, p24 proteins of the delta subfamily (p24δ5 and p24δ9) were shown to interact with two different K/HDEL receptors, ERD2a and ERD2b. This interaction required the GOLD domain in p24δ5 and was optimal at acidic pH but very low at neutral pH, consistent with the interaction taking place at the Golgi apparatus (Montesinos et al. 2014). p24δ5 was also found to inhibit the secretion of HDEL ligands, but not constitutive secretion, showing a role for p24δ5 in retrograde Golgi-to-ER transport of K/HDEL ligands. In addition, p24δ5 and ERD2 were shown to interact with ARF1 and COPI proteins in a pH-dependent manner, being optimal at acidic pH and very low at neutral pH. ERD2 and p24δ5 can also interact with the Sec23 subunit of the COPII coat, but this interaction is optimal at neutral pH (Montesinos et al. 2014).
Altogether, pH-dependent interactions seem to play an essential role for bidirectional transport of ERD2 and K/HDEL ligands. The slightly acidic pH of the Golgi apparatus is also required for binding of K/HDEL ligands to ERD2. Ligand binding to ERD2 induces ERD2 oligomerization and triggers the interaction between ERD2 and ARF1, ERD2 and ARF-GAP, and ARF1 and ARF-GAP. The acidic pH of the Golgi apparatus also favors the interaction between p24 proteins and ERD2 and the interaction of both ERD2 and p24 proteins with ARF1 and COPI proteins, leading to the formation of COPI vesicles enriched in K/HDEL ligands (Fig. 4). Once in the ER lumen, at neutral pH, p24 proteins dissociate from ERD2 and ERD2 from K/HDEL ligands.
Other putative p24 cargos in plants
In plants, a p24 protein named CYB, based in its phenotype (accumulation in cytoplasmic bodies), which in the standard nomenclature corresponds to p24δ4, has been proposed to be required for trafficking of GLL23, a putative myrosinase-associated protein (Jancowski et al. 2014). Myrosinase is a β-glucosidase present in the Brassicaceae which hydrolyzes glucosinolates to produce a variety of products with varying toxicity to pathogens and insect herbivores, such as isothiocyanates, epithionitriles, nitriles, or thiocyanates (Kliebenstein et al. 2005). In this respect, myrosinase-associated proteins, such as GLL23, may affect the product composition of glucosinolate hydrolysis (Agee et al. 2010). GLL23 has also been shown to be part of defense-related PYK 10 complexes that form upon cell damage (Nagano et al. 2008). p24δ4 was proposed to function as a cargo receptor for efficient ER-export of GLL23, perhaps to prevent an accumulation of this protein in the ER, where it may be detrimental for ER organization. In this respect, the cyb mutation was found to affect the GOLD domain in p24δ4, which is presumably involved in cargo recognition (Jancowski et al. 2014).
p24 proteins and organization of the early secretory pathway
Several reports have suggested that p24 proteins may be involved in the structure and organization of the compartments of the early secretory pathway, including the formation of ER exit sites (ERES), the structure of the ER and the ERGIC and the biogenesis and maintenance of the Golgi apparatus. Using a cell-free system to study the assembly of transitional ER and the formation of ER exit sites (ERES), p24α2 was suggested to play a role in this process, being the first proposal of a possible structural role for p24 family proteins (Lavoie et al. 1999). This is consistent with the enrichment of p24 proteins in COPII vesicles (Belden and Barlowe 1996, Marzioch et al. 1999) and the dramatic reduction of ER-derived vesicles in a yeast Emp24p (p24β) KO strain (Stamnes et al. 1995).
Emery et al. (2000) found that overexpression of p23 (p24δ1) led to the loss of normal Golgi morphology with a fragmentation of Golgi ribbon and the appearance of smaller Golgi fragments (i.e., Golgi mini-stacks). These results were consistent with the alteration in Golgi morphology obtained after overexpression of GFP-tagged p23 (p24δ1) and p24 (p24β1) (Blum et al. 1999). Based on these observations, it has been proposed that p23 (p24δ1) may have a morphogenic function which contributes to the organization of the Golgi apparatus (Emery et al. 2000). Indeed, the inactivation of one allele of p23 (p24δ1) in mice was shown to induce structural changes in the Golgi apparatus, with dilation of the Golgi cisternae which was more prominent at the rim (Denzel et al. 2000). p23 (p24δ1) overexpression has also been shown to cause Golgi fragmentation in a subset of neurons (Gong et al. 2011), consistent with the results of Emery et al. (2000). Silencing of p25 (p24α2) in HeLa cells caused a reduction in the number of ERGIC clusters and destabilized the ERGIC without affecting the number of ER exit sites (labeled with Sec31). It also led to Golgi fragmentation, with the Golgi ribbon converted to Golgi mini-stacks. However, the cisternae were not swollen and cisternal stacking remained intact, which suggested a normal cis-trans topology (Mitrovic et al. 2008). p25 silencing also caused COPI dissociation from Golgi membranes, but the dispersed Golgi remained associated to Golgi matrix proteins. Silencing of p28 (p24γ2) in human hepatocellular carcinoma (HepG2) did not change the morphology of the ER or the ERGIC (in contrast to p25 silencing), but caused Golgi fragmentation, showing Golgi mini-stacks with apparently unchanged cis-trans topology (Koegler et al. 2010). However, COPI subunits remained Golgi-associated in p28-depleted cells, in contrast to p25-silenced cells. Interestingly, this Golgi fragmentation did not cause an alteration of anterograde or retrograde traffic (Koegler et al. 2010). Knockdown experiments of p24A (p24β1) in HEK-293 cells also showed a disruption of the Golgi apparatus, as judged by a diffuse cytosolic staining of the Golgi marker GM130 (Luo et al. 2007).
Using a mutant version of p25 lacking the KKXX motif, it has been proposed that p24 proteins may form specialized membrane domains rich in cholesterol (Emery et al. 2003). This would be consistent with the partial partitioning of p23 and p25 in lipid rafts in ER membranes, as described above.
To our knowledge, there are no reports on the putative roles of p24 proteins in the organization of the compartments of the early secretory pathways in plant cells. Transient expression of XFP-tagged p24 proteins in tobacco protoplasts or N. benthamiana leaves did not show any obvious alteration in the morphology of the compartments of the secretory pathway, even under conditions of overexpression (Langhans et al. 2008, Montesinos et al. 2012, 2013). Single p24 KO mutants in Arabidopsis also did not show any obvious ultrastructural alterations (Montesinos et al. 2012), perhaps because of functional redundancy with other p24 family members.
p24 proteins in physiology and pathology
p24 proteins have recently become very popular recently because of their implication in a variety of specific functions in animal cells, including early embryonic mouse development and morphogenesis of the mouse embryo and placenta, insulin biosynthesis and subsequent secretion in pancreatic beta cells, or amyloid precursor metabolism and pathogenesis of Alzheimer’s disease, as described in this section.
p24 proteins in development
Several studies have shown that p24 proteins have a role in development. In mice, the loss of p23/TMED10 (p24δ1), that also led to reduced levels of p24α2 and p24γ4 proteins, resulted in early embryonic lethality (Denzel et al. 2000). The loss of TMED2 (p24β1) caused death at midgestation and prior to death, the embryos showed developmental delay and absence of the labyrinth layer of the placenta. The loss of TMED2 also led to reduced levels of TMED9 (p24α2) and to the loss of p24γ3 and p23/TMED10. Since the mutant embryos lacking TMED10 arrest and die prior Tmed2 mutants, it was concluded that individual p24 proteins are essential for distinct stage and tissue-specific functions during development (Jerome-Majewska et al. 2010).
As it has been said above, Drosophila p24 transcripts show a variety of spatial, temporal and tissue-specific expression patterns. Characterization of different p24 mutants and the use of RNA interference (RNAi) to reduce p24 expression during development have both shown that most Drosophila p24 genes are required during development (Boltz et al. 2007, Saleem et al. 2012). These studies also showed that every p24 except CG9308 (p24β) function in some aspects of reproduction and have a role both in male and female reproduction. For instance, mutation in loj (p24γ), eca (p24α), or bai (p24δ) result in females that are unable to oviposit and eca and bai males that have reduced fertility. Interestingly, neural expression of p24s has been involved in regulating female behavior as loj expression in octopaminergic neuron has been shown to rescue the loj mutant oviposition defect (Boltz et al. 2007, Saleem et al. 2012).
Additional D. melanogaster genetic approaches have been performed to investigate p24 functions in development that involved Wnt signaling. As described before, it has been shown that transport of Wnt proteins requires p24 proteins (Buechling et al. 2011; Port et al. 2011). Using large-scale RNA interference (RNAi) screens, it was found that knockdown of specific p24 proteins leads to a reduction in secretion of Wg although no effects on general protein secretion were detected. Further in vivo experiments showed that the knockdown of these p24 proteins in the wing imaginal disc resulted in the accumulation of Wg in producing cells. In addition, a reduction in Wg target gene expression and defects in the formation of wing margin tissue were observed (Buechling et al. 2011; Port et al. 2011).
p24 proteins, insulin secretion, and diabetes
Some p24 proteins have been shown to be abundantly expressed in secretory cell types. This is the case of Tmp21 (p24δ1) and p24A (p24β1) in rat pancreatic acinar cells (Blum et al. 1996), p23 (p24δ1) in endocrine-derived cell lines (Hosaka et al. 2007), rat islets or insulinoma cell lines (Zhang and Volchuk 2010) or TMED6 (p24γ3) in rat pancreatic islets (Wang et al. 2012). p23, which mainly localized to cis-Golgi membranes, was also found to partially localize to the membrane of secretory granules in pancreatic β-cells (Hosaka et al. 2007), raising the possibility that p24 proteins may also regulate post-Golgi trafficking events in these cells.
Knocking down p24δ1 in insulinoma cell lines, which caused a concomitant reduction of other p24 family members (p24β1 and p24α2), decreased glucose-stimulated insulin secretion, with no effect on basal secretion. It inhibited proinsulin biosynthesis and decreased total cellular insulin content (Zhang and Volchuk 2010). The same reduction in insulin secretion was observed upon knockdown of TMED6 (p24γ3) (Wang et al. 2012). These results suggest that p24 proteins (including p24δ1 and p24γ3) are required for insulin biosynthesis and subsequent secretion in pancreatic β-cells (Zhang and Volchuk 2010). However, a direct interaction between pro-insulin and p24 proteins has not been demonstrated. Interestingly, TMED6 gene expression has been found to be decreased in diabetic rats, suggesting a role for TMED6 in diabetes.
p24 proteins and Alzheimer’s disease
Alzheimer’s disease is characterized by the cerebral deposition of beta-amyloid (Aβ) peptides in senile plaques. Aβ peptides are generated by sequential cleavages of the amyloid-β precursor protein (APP) by β- and γ-secretase activities (Thinakaran and Koo 2008). APP is first cleaved within its luminal domain by β-secretase to generate a soluble fragment (which is secreted to the extracellular space) and a membrane-bound C-terminal fragment. The latter is then cleaved by the γ-secretase at several sequences, such as the γ-site, to produce Aβ40/42, and the ε-site, which results in the release of the cytosolic amyloid intracellular domain (Thinakaran and Koo 2008). It has been shown that the γ- and ε-secretase activities are part of a multimeric membrane protein complex, the presenilin complex (Iwatsubo 2004). Interestingly, this complex has been found to contain the p24 protein p23/TMP21 (p24δ1), which modulates γ-secretase activity (Chen et al. 2006). p23/TMP21 has been shown to interact with the presenilin complex via its transmembrane domain (Pardossi-Piquard et al. 2009) and to cause inhibition of γ-secretase (but not ε-secretase) activity (Chen et al. 2006). As a consequence, silencing of p23/TMP21 increased the production of both Aβ40 and Aβ42 in human embryonic kidney (HEK-293) cells (Chen et al. 2006, Dolcini et al. 2008) and in N2a neuroblastoma cells (Vetrivel et al. 2007). In addition, p23/TMP21 was found to inhibit transport of APP in the secretory pathway and to reduce cell surface accumulation of fully glycosylated APP (Vetrivel et al. 2007). p24α2, which has been shown recently to localize to pre-synaptic terminals in the mammalian brain (Liu et al. 2015), has also been shown to cooperate with p24δ1 in attenuating Aβ production, in contrast to p24β2, p24γ3 or p24γ4. This activity requires the dilysine motif in the cytosolic tail of both proteins (Hasegawa et al. 2010), which highlights the importance of sorting determinants in the cytosolic tail of p24 proteins for their trafficking and function. In summary, both p24δ1 and p24α2 may well be involved in the pathogenesis of Alzheimer’s disease modulating Aβ production. As mentioned before, the steady-state levels of p23 have been shown to be high during embryonic development and to decline during postnatal development (Vetrivel et al. 2008). Besides, the steady-state levels of p23 have been shown to be reduced in the brain of individuals with AD, which may explain the enhanced beta-amyloid production (Vetrivel et al. 2008). Therefore, increasing p23 expression has been proposed as a potential strategy to reduce Aβ levels (Gong et al. 2011).
p24 proteins and apoptosis
Protein kinase Cδ (PKCδ), a kinase widely implicated in apoptosis and inhibition of cell cycle progression, has been shown to interact with p23/Tmp21 (p24δ1) (Wang and Kazanietz 2010, Wang et al. 2011). By interacting with the C1 domain of PKCδ, p23/Tmp1 has been proposed to retain PKCδ in a perinuclear compartment (presumably the Golgi apparatus) and to prevent its transport to the plasma membrane, thereby reducing its ability for activation in response to stimuli and thus modulating its apoptotic function (Wang et al. 2011).
Concluding remarks
p24 family proteins are essential regulators of protein trafficking along the secretory pathway, regulating the composition, structure, and function of the different organelles of the pathway, mainly the ER and the Golgi apparatus. In addition, they appear to modulate the transport of specific cargos, thus playing specific roles in signaling, development, insulin secretion, or the pathogenesis of Alzheimer’s disease. As a consequence, they have recently been proposed to be potential targets for disease treatment. In contrast, very little is known about the specific functions of p24 proteins in plants since functional studies are still very scarce.
References
Agee AE, Surpin M, Sohn EJ, Girke T, Rosado A, Kram BW, Carter C, Wentzell AM, Kliebenstein DJ, Jin HC, Park OK, Jin H, Hicks GR, Raikhel NV (2010) MODIFIED VACUOLE PHENOTYPE1 is an Arabidopsis myrosinase-associated protein involved in endomembrane protein trafficking. Plant Physiol 152:120–132
Aguilera-Romero A, Kaminska J, Spang A, Riezman H, Muniz M (2008) The yeast p24 complex is required for the formation of COPI retrograde transport vesicles from the Golgi apparatus. J Cell Biol 180:713–720
Anantharaman V, Aravind L (2002) The GOLD domain, a novel protein module involved in Golgi function and secretion. Genome Biol 3: research0023
Aniento F, Matsuoka K, Robinson DG (2006) ER-to-Golgi transport: The COPII-pathway. In: Robinson DG (ed) The Plant Endoplasmatic Reticulum. Springer-Verlag, Berlin-Heidelberg, pp 99–124
Antonny B, Beraud-Dufour S, Chardin P, Chabre M (1997) N-terminal hydrophobic residues of the G-protein ADP-ribosylation factor-1 insert into membrane phospholipids upon GDP to GTP exchange. Biochemistry 36:4675–4684
Aoe T, Cukierman E, Lee A, Cassel D, Peters PJ, Hsu VW (1997) The KDEL receptor, ERD2, regulates intracellular traffic by recruiting a GTPase-activating protein for ARF1. EMBO J 16:7305–7316
Aoe T, Lee AJ, van Donselaar E, Peters PJ, Hsu VW (1998) Modulation of intracellular transport by transported proteins: insight from regulation of COPI-mediated transport. Proc Natl Acad Sci U S A 95:1624–1629
Barlowe C (2003) Signals for COPII-dependent export from the ER: what's the ticket out? Trends Cell Biol 13:295–300
Belden WJ, Barlowe C (2001) Distinct roles for the cytoplasmic tail sequences of Emp24p and Erv25p in transport between the endoplasmic reticulum and Golgi complex. J Biol Chem 276:43040–43048
Belden WJ, Barlowe C (1996) Erv25p, a component of COPII-coated vesicles, forms a complex with Emp24p that is required for efficient endoplasmic reticulum to Golgi transport. J Biol Chem 271:26939–26946
Bethune J, Kol M, Hoffmann J, Reckmann I, Brugger B, Wieland F (2006) Coatomer, the coat protein of COPI transport vesicles, discriminates endoplasmic reticulum residents from p24 proteins. Mol Cell Biol 26:8011–8021
Bjorkholm P, Ernst AM, Hacke M, Wieland F, Brugger B, von Heijne G (2014) Identification of novel sphingolipid-binding motifs in mammalian membrane proteins. Biochim Biophys Acta 1838:2066–2070
Blum R, Feick P, Puype M, Vandekerckhove J, Klengel R, Nastainczyk W, Schulz I (1996) Tmp21 and p24A, two type I proteins enriched in pancreatic microsomal membranes, are members of a protein family involved in vesicular trafficking. J Biol Chem 271:17183–17189
Blum R, Lepier A (2008) The luminal domain of p23 (Tmp21) plays a critical role in p23 cell surface trafficking. Traffic 9:1530–1550
Blum R, Pfeiffer F, Feick P, Nastainczyk W, Kohler B, Schafer KH, Schulz I (1999) Intracellular localization and in vivo trafficking of p24A and p23. J Cell Sci 112(Pt 4):537–548
Boltz KA, Carney GE (2008) Loss of p24 function in Drosophila melanogaster causes a stress response and increased levels of NF-kappaB-regulated gene products. BMC Genomics 9:212
Boltz KA, Ellis LL, Carney GE (2007) Drosophila melanogaster p24 genes have developmental, tissue-specific, and sex-specific expression patterns and functions. Dev Dyn 236:544–555
Bonnon C, Wendeler MW, Paccaud JP, Hauri HP (2010) Selective export of human GPI-anchored proteins from the endoplasmic reticulum. J Cell Sci 123:1705–1715
Brandizzi F, Barlowe C (2013) Organization of the ER-Golgi interface for membrane traffic control. Nat Rev Mol Cell Biol 14:382–392
Bremser M, Nickel W, Schweikert M, Ravazzola M, Amherdt M, Hughes CA, Sollner TH, Rothman JE, Wieland FT (1999) Coupling of coat assembly and vesicle budding to packaging of putative cargo receptors. Cell 96:495–506
Bryant CE, Spring DR, Gangloff M, Gay NJ (2010) The molecular basis of the host response to lipopolysaccharide. Nat Rev Microbiol 8:8–14
Buechling T, Chaudhary V, Spirohn K, Weiss M, Boutros M (2011) p24 proteins are required for secretion of Wnt ligands. EMBO Rep 12:1265–1272
Cabrera M, Muniz M, Hidalgo J, Vega L, Martin ME, Velasco A (2003) The retrieval function of the KDEL receptor requires PKA phosphorylation of its C-terminus. Mol Biol Cell 14:4114–4125
Castillon GA, Aguilera-Romero A, Manzano-Lopez J, Epstein S, Kajiwara K, Funato K, Watanabe R, Riezman H, Muniz M (2011) The yeast p24 complex regulates GPI-anchored protein transport and quality control by monitoring anchor remodeling. Mol Biol Cell 22:2924–2936
Chen F, Hasegawa H, Schmitt-Ulms G, Kawarai T, Bohm C, Katayama T, Gu Y, Sanjo N, Glista M, Rogaeva E, Wakutani Y, Pardossi-Piquard R, Ruan X, Tandon A, Checler F, Marambaud P, Hansen K, Westaway D, St George-Hyslop P, Fraser P (2006) TMP21 is a presenilin complex component that modulates gamma-secretase but not epsilon-secretase activity. Nature 440:1208–1212
Chen J, Qi X, Zheng H (2012) Subclass-specific localization and trafficking of Arabidopsis p24 proteins in the ER-Golgi interface. Traffic 13:400–415
Ciufo LF, Boyd A (2000) Identification of a lumenal sequence specifying the assembly of Emp24p into p24 complexes in the yeast secretory pathway. J Biol Chem 275:8382–8388
Contreras FX, Ernst AM, Haberkant P, Bjorkholm P, Lindahl E, Gonen B, Tischer C, Elofsson A, von Heijne G, Thiele C, Pepperkok R, Wieland F, Brugger B (2012) Molecular recognition of a single sphingolipid species by a protein's transmembrane domain. Nature 481:525–529
Contreras I, Ortiz-Zapater E, Aniento F (2004a) Sorting signals in the cytosolic tail of membrane proteins involved in the interaction with plant ARF1 and coatomer. Plant J 38:685–698
Contreras I, Yang Y, Robinson DG, Aniento F (2004b) Sorting signals in the cytosolic tail of plant p24 proteins involved in the interaction with the COPII coat. Plant Cell Physiol 45:1779–1786
Copic A, Latham CF, Horlbeck MA, D'Arcangelo JG, Miller EA (2012) ER cargo properties specify a requirement for COPII coat rigidity mediated by Sec13p. Science 335:1359–1362
D'Arcangelo JG, Crissman J, Pagant S, Copic A, Latham CF, Snapp EL, Miller EA (2015) Traffic of p24 Proteins and COPII Coat Composition Mutually Influence Membrane Scaffolding. Curr Biol 25:1296–1305
Denzel A, Otto F, Girod A, Pepperkok R, Watson R, Rosewell I, Bergeron JJ, Solari RC, Owen MJ (2000) The p24 family member p23 is required for early embryonic development. Curr Biol 10:55–58
Dolcini V, Dunys J, Sevalle J, Chen F, Guillot-Sestier MV, St George-Hyslop P, Fraser PE, Checler F (2008) TMP21 regulates Abeta production but does not affect caspase-3, p53, and neprilysin. Biochem Biophys Res Commun 371:69–74
Dominguez M, Dejgaard K, Fullekrug J, Dahan S, Fazel A, Paccaud JP, Thomas DY, Bergeron JJ, Nilsson T (1998) gp25L/emp24/p24 protein family members of the cis-Golgi network bind both COP I and II coatomer. J Cell Biol 140:751–765
Emery G, Parton RG, Rojo M, Gruenberg J (2003) The trans-membrane protein p25 forms highly specialized domains that regulate membrane composition and dynamics. J Cell Sci 116:4821–4832
Emery G, Rojo M, Gruenberg J (2000) Coupled transport of p24 family members. J Cell Sci 113(Pt 13):2507–2516
Fiedler K, Veit M, Stamnes MA, Rothman JE (1996) Bimodal interaction of coatomer with the p24 family of putative cargo receptors. Science 273:1396–1399
Fox RM, Hanlon CD, Andrew DJ (2010) The CrebA/Creb3-like transcription factors are major and direct regulators of secretory capacity. J Cell Biol 191:479–492
Franco M, Chardin P, Chabre M, Paris S (1996) Myristoylation-facilitated binding of the G protein ARF1GDP to membrane phospholipids is required for its activation by a soluble nucleotide exchange factor. J Biol Chem 271:1573–1578
Fujita M, Watanabe R, Jaensch N, Romanova-Michaelides M, Satoh T, Kato M, Riezman H, Yamaguchi Y, Maeda Y, Kinoshita T (2011) Sorting of GPI-anchored proteins into ER exit sites by p24 proteins is dependent on remodeled GPI. J Cell Biol 194:61–75
Fullekrug J, Suganuma T, Tang BL, Hong W, Storrie B, Nilsson T (1999) Localization and recycling of gp27 (hp24gamma3): complex formation with other p24 family members. Mol Biol Cell 10:1939–1955
Gao C, Cai Y, Wang Y, Kang BH, Aniento F, Robinson DG, Jiang L (2014) Retention mechanisms for ER and Golgi membrane proteins. Trends Plant Sci 19:508–515
Gaskell A, Crennell S, Taylor G (1995) The three domains of a bacterial sialidase: a beta-propeller, an immunoglobulin module and a galactose-binding jelly-roll. Structure 3:1197–1205
Goldberg J (2000) Decoding of sorting signals by coatomer through a GTPase switch in the COPI coat complex. Cell 100:671–679
Gommel D, Orci L, Emig EM, Hannah MJ, Ravazzola M, Nickel W, Helms JB, Wieland FT, Sohn K (1999) p24 and p23, the major transmembrane proteins of COPI-coated transport vesicles, form hetero-oligomeric complexes and cycle between the organelles of the early secretory pathway. FEBS Lett 447:179–185
Gong P, Roseman J, Fernandez CG, Vetrivel KS, Bindokas VP, Zitzow LA, Kar S, Parent AT, Thinakaran G (2011) Transgenic neuronal overexpression reveals that stringently regulated p23 expression is critical for coordinated movement in mice. Mol Neurodegener 6: 87-1326-6-87
Gross JC, Boutros M (2013) Secretion and extracellular space travel of Wnt proteins. Curr Opin Genet Dev 23:385–390
Hara-Kuge S, Kuge O, Orci L, Amherdt M, Ravazzola M, Wieland FT, Rothman JE (1994) En bloc incorporation of coatomer subunits during the assembly of COP-coated vesicles. J Cell Biol 124:883–892
Harter C, Pavel J, Coccia F, Draken E, Wegehingel S, Tschochner H, Wieland F (1996) Nonclathrin coat protein gamma, a subunit of coatomer, binds to the cytoplasmic dilysine motif of membrane proteins of the early secretory pathway. Proc Natl Acad Sci U S A 93:1902–1906
Hartley T, Siva M, Lai E, Teodoro T, Zhang L, Volchuk A (2010) Endoplasmic reticulum stress response in an INS-1 pancreatic beta-cell line with inducible expression of a folding-deficient proinsulin. BMC Cell Biol 11:59
Hasegawa H, Liu L, Nishimura M (2010) Dilysine retrieval signal-containing p24 proteins collaborate in inhibiting gamma-cleavage of amyloid precursor protein. J Neurochem 115:771–781
Hirata R, Nihei C, Nakano A (2013) Isoform-selective oligomer formation of Saccharomyces cerevisiae p24 family proteins. J Biol Chem 288:37057–37070
Hosaka M, Watanabe T, Yamauchi Y, Sakai Y, Suda M, Mizutani S, Takeuchi T, Isobe T, Izumi T (2007) A subset of p23 localized on secretory granules in pancreatic beta-cells. J Histochem Cytochem 55:235–245
Iwatsubo T (2004) The gamma-secretase complex: machinery for intramembrane proteolysis. Curr Opin Neurobiol 14:379–383
Jackson LP (2014) Structure and mechanism of COPI vesicle biogenesis. Curr Opin Cell Biol 29:67–73
Jackson LP, Lewis M, Kent HM, Edeling MA, Evans PR, Duden R, Owen DJ (2012) Molecular basis for recognition of dilysine trafficking motifs by COPI. Dev Cell 23:1255–1262
Jancowski S, Catching A, Pighin J, Kudo T, Foissner I, Wasteneys GO (2014) Trafficking of the myrosinase-associated protein GLL23 requires NUC/MVP1/GOLD36/ERMO3 and the p24 protein CYB. Plant J 77:497–510
Jenne N, Frey K, Brugger B, Wieland FT (2002) Oligomeric state and stoichiometry of p24 proteins in the early secretory pathway. J Biol Chem 277:46504–46511
Jerome-Majewska LA, Achkar T, Luo L, Lupu F, Lacy E (2010) The trafficking protein Tmed2/p24beta(1) is required for morphogenesis of the mouse embryo and placenta. Dev Biol 341:154–166
Kamauchi S, Nakatani H, Nakano C, Urade R (2005) Gene expression in response to endoplasmic reticulum stress in Arabidopsis thaliana. FEBS J 272:3461–3476
Kinoshita T, Maeda Y, Fujita M (2013) Transport of glycosylphosphatidylinositol-anchored proteins from the endoplasmic reticulum. Biochim Biophys Acta 1833:2473–2478
Kliebenstein DJ, Kroymann J, Mitchell-Olds T (2005) The glucosinolate-myrosinase system in an ecological and evolutionary context. Curr Opin Plant Biol 8:264–271
Koegler E, Bonnon C, Waldmeier L, Mitrovic S, Halbeisen R, Hauri HP (2010) p28, a novel ERGIC/cis Golgi protein, required for Golgi ribbon formation. Traffic 11:70–89
Langer JD, Roth CM, Bethune J, Stoops EH, Brugger B, Herten DP, Wieland FT (2008) A conformational change in the alpha-subunit of coatomer induced by ligand binding to gamma-COP revealed by single-pair FRET. Traffic 9:597–607
Langhans M, Marcote MJ, Pimpl P, Virgili-Lopez G, Robinson DG, Aniento F (2008) In vivo trafficking and localization of p24 proteins in plant cells. Traffic 9:770–785
Lanoix J, Ouwendijk J, Stark A, Szafer E, Cassel D, Dejgaard K, Weiss M, Nilsson T (2001) Sorting of Golgi resident proteins into different subpopulations of COPI vesicles: a role for ArfGAP1. J Cell Biol 155:1199–1212
Lavoie C, Paiement J, Dominguez M, Roy L, Dahan S, Gushue JN, Bergeron JJ (1999) Roles for alpha(2)p24 and COPI in endoplasmic reticulum cargo exit site formation. J Cell Biol 146:285–299
Li X, Wu Y, Shen C, Belenkaya TY, Ray L, Lin X (2015) Drosophila p24 and Sec22 regulate Wingless trafficking in the early secretory pathway. Biochem Biophys Res Commun. doi:10.1016/j.bbrc.2015.04.151
Liaunardy-Jopeace A, Bryant CE, Gay NJ (2014) The COP II adaptor protein TMED7 is required to initiate and mediate the delivery of TLR4 to the plasma membrane. Sci Signal 7:ra70
Liaunardy-Jopeace A, Gay NJ (2014) Molecular and cellular regulation of toll-like receptor-4 activity induced by lipopolysaccharide ligands. Front Immunol 5:473
Liu L, Fujino K, Nishimura M (2015) Pre-synaptic localization of the gamma-secretase-inhibiting protein p24alpha2 in the mammalian brain. J Neurochem 133:422–431
Liu S, Bromley-Brits K, Xia K, Mittelholtz J, Wang R, Song W (2008) TMP21 degradation is mediated by the ubiquitin-proteasome pathway. Eur J Neurosci 28:1980–1988
Liu S, Zhang S, Bromley-Brits K, Cai F, Zhou W, Xia K, Mittelholtz J, Song W (2011) Transcriptional Regulation of TMP21 by NFAT. Mol Neurodegener 6:21
Luo W, Wang Y, Reiser G (2011) Proteinase-activated receptors, nucleotide P2Y receptors, and mu-opioid receptor-1B are under the control of the type I transmembrane proteins p23 and p24A in post-Golgi trafficking. J Neurochem 117:71–81
Luo W, Wang Y, Reiser G (2007) p24A, a type I transmembrane protein, controls ARF1-dependent resensitization of protease-activated receptor-2 by influence on receptor trafficking. J Biol Chem 282:30246–30255
Ma W, Goldberg J (2013) Rules for the recognition of dilysine retrieval motifs by coatomer. EMBO J 32:926–937
Majoul I, Straub M, Hell SW, Duden R, Soling HD (2001) KDEL-cargo regulates interactions between proteins involved in COPI vesicle traffic: measurements in living cells using FRET. Dev Cell 1:139–153
Manzano-Lopez J, Perez-Linero AM, Aguilera-Romero A, Martin ME, Okano T, Silva DV, Seeberger PH, Riezman H, Funato K, Goder V, Wellinger RE, Muniz M (2015) COPII coat composition is actively regulated by luminal cargo maturation. Curr Biol 25:152–162
Marelli M, Smith JJ, Jung S, Yi E, Nesvizhskii AI, Christmas RH, Saleem RA, Tam YY, Fagarasanu A, Goodlett DR, Aebersold R, Rachubinski RA, Aitchison JD (2004) Quantitative mass spectrometry reveals a role for the GTPase Rho1p in actin organization on the peroxisome membrane. J Cell Biol 167:1099–1112
Martiniere A, Bassil E, Jublanc E, Alcon C, Reguera M, Sentenac H, Blumwald E, Paris N (2013) In vivo intracellular pH measurements in tobacco and Arabidopsis reveal an unexpected pH gradient in the endomembrane system. Plant Cell 25:4028–4043
Marzioch M, Henthorn DC, Herrmann JM, Wilson R, Thomas DY, Bergeron JJ, Solari RC, Rowley A (1999) Erp1p and Erp2p, partners for Emp24p and Erv25p in a yeast p24 complex. Mol Biol Cell 10:1923–1938
Matsuoka K, Orci L, Amherdt M, Bednarek SY, Hamamoto S, Schekman R, Yeung T (1998) COPII-coated vesicle formation reconstituted with purified coat proteins and chemically defined liposomes. Cell 93:263–275
Miller EA, Beilharz TH, Malkus PN, Lee MC, Hamamoto S, Orci L, Schekman R (2003) Multiple cargo binding sites on the COPII subunit Sec24p ensure capture of diverse membrane proteins into transport vesicles. Cell 114:497–509
Mitrovic S, Ben-Tekaya H, Koegler E, Gruenberg J, Hauri HP (2008) The cargo receptors Surf4, endoplasmic reticulum-Golgi intermediate compartment (ERGIC)-53, and p25 are required to maintain the architecture of ERGIC and Golgi. Mol Biol Cell 19:1976–1990
Montesinos JC, Langhans M, Sturm S, Hillmer S, Aniento F, Robinson DG, Marcote MJ (2013) Putative p24 complexes in Arabidopsis contain members of the delta and beta subfamilies and cycle in the early secretory pathway. J Exp Bot 64:3147–3167
Montesinos JC, Pastor-Cantizano N, Robinson DG, Marcote MJ, Aniento F (2014) Arabidopsis p24delta5 and p24delta9 facilitate Coat Protein I-dependent transport of the K/HDEL receptor ERD2 from the Golgi to the endoplasmic reticulum. Plant J 80:1014–1030
Montesinos JC, Sturm S, Langhans M, Hillmer S, Marcote MJ, Robinson DG, Aniento F (2012) Coupled transport of Arabidopsis p24 proteins at the ER-Golgi interface. J Exp Bot 63:4243–4261
Muniz M, Nuoffer C, Hauri HP, Riezman H (2000) The Emp24 complex recruits a specific cargo molecule into endoplasmic reticulum-derived vesicles. J Cell Biol 148:925–930
Nagano AJ, Fukao Y, Fujiwara M, Nishimura M, Hara-Nishimura I (2008) Antagonistic jacalin-related lectins regulate the size of ER body-type beta-glucosidase complexes in Arabidopsis thaliana. Plant Cell Physiol 49:969–980
Nickel W, Sohn K, Bunning C, Wieland FT (1997) p23, a major COPI-vesicle membrane protein, constitutively cycles through the early secretory pathway. Proc Natl Acad Sci U S A 94:11393–11398
Osiecka-Iwan A, Niderla-Bielinska J, Hyc A, Moskalewski S (2014) Rat chondrocyte-associated antigen identified as sialylated transmembrane protein Tmp21 belonging to the p24 protein family. Calcif Tissue Int 94:348–352
Pardossi-Piquard R, Bohm C, Chen F, Kanemoto S, Checler F, Schmitt-Ulms G, St George-Hyslop P, Fraser PE (2009) TMP21 transmembrane domain regulates gamma-secretase cleavage. J Biol Chem 284:28634–28641
Paroutis P, Touret N, Grinstein S (2004) The pH of the secretory pathway: measurement, determinants, and regulation. Physiology (Bethesda) 19:207–215
Popoff V, Adolf F, Brugger B, Wieland F (2011) COPI budding within the Golgi stack. Cold Spring Harb Perspect Biol 3:a005231
Port F, Hausmann G, Basler K (2011) A genome-wide RNA interference screen uncovers two p24 proteins as regulators of Wingless secretion. EMBO Rep 12:1144–1152
Reinhard C, Harter C, Bremser M, Brugger B, Sohn K, Helms JB, Wieland F (1999) Receptor-induced polymerization of coatomer. Proc Natl Acad Sci U S A 96:1224–1228
Rojo M, Emery G, Marjomaki V, McDowall AW, Parton RG, Gruenberg J (2000) The transmembrane protein p23 contributes to the organization of the Golgi apparatus. J Cell Sci 113(Pt 6):1043–1057
Rojo M, Pepperkok R, Emery G, Kellner R, Stang E, Parton RG, Gruenberg J (1997) Involvement of the transmembrane protein p23 in biosynthetic protein transport. J Cell Biol 139:1119–1135
Rotter J, Kuiper RP, Bouw G, Martens GJ (2002) Cell-type-specific and selectively induced expression of members of the p24 family of putative cargo receptors. J Cell Sci 115:1049–1058
Saleem S, Schwedes CC, Ellis LL, Grady ST, Adams RL, Johnson N, Whittington JR, Carney GE (2012) Drosophila melanogaster p24 trafficking proteins have vital roles in development and reproduction. Mech Dev 129:177–191
Schimmoller F, Singer-Kruger B, Schroder S, Kruger U, Barlowe C, Riezman H (1995) The absence of Emp24p, a component of ER-derived COPII-coated vesicles, causes a defect in transport of selected proteins to the Golgi. EMBO J 14:1329–1339
Schuiki I, Volchuk A (2012) Diverse roles for the p24 family of proteins in eukaryotic cells. Biomol Concepts 3:561–570
Shen J, Zeng Y, Zhuang X, Sun L, Yao X, Pimpl P, Jiang L (2013) Organelle pH in the Arabidopsis endomembrane system. Mol Plant 6:1419–1437
Simpson JC, Nilsson T, Pepperkok R (2006) Biogenesis of tubular ER-to-Golgi transport intermediates. Mol Biol Cell 17:723–737
Sohn K, Orci L, Ravazzola M, Amherdt M, Bremser M, Lottspeich F, Fiedler K, Helms JB, Wieland FT (1996) A major transmembrane protein of Golgi-derived COPI-coated vesicles involved in coatomer binding. J Cell Biol 135:1239–1248
Springer S, Chen E, Duden R, Marzioch M, Rowley A, Hamamoto S, Merchant S, Schekman R (2000) The p24 proteins are not essential for vesicular transport in Saccharomyces cerevisiae. Proc Natl Acad Sci U S A 97:4034–4039
Stamnes MA, Craighead MW, Hoe MH, Lampen N, Geromanos S, Tempst P, Rothman JE (1995) An integral membrane component of coatomer-coated transport vesicles defines a family of proteins involved in budding. Proc Natl Acad Sci U S A 92:8011–8015
Strating JR, Martens GJ (2009) The p24 family and selective transport processes at the ER-Golgi interface. Biol Cell 101:495–509
Strating JR, van Bakel NH, Leunissen JA, Martens GJ (2009) A comprehensive overview of the vertebrate p24 family: identification of a novel tissue-specifically expressed member. Mol Biol Evol 26:1707–1714
Takida S, Maeda Y, Kinoshita T (2008) Mammalian GPI-anchored proteins require p24 proteins for their efficient transport from the ER to the plasma membrane. Biochem J 409:555–562
Tanaka K, Okabayashi K, Asashima M, Perrimon N, Kadowaki T (2000) The evolutionarily conserved porcupine gene family is involved in the processing of the Wnt family. Eur J Biochem 267:4300–4311
Teasdale RD, Jackson MR (1996) Signal-mediated sorting of membrane proteins between the endoplasmic reticulum and the golgi apparatus. Annu Rev Cell Dev Biol 12:27–54
Theiler R, Fujita M, Nagae M, Yamaguchi Y, Maeda Y, Kinoshita T (2014) The alpha-helical region in p24gamma2 subunit of p24 protein cargo receptor is pivotal for the recognition and transport of glycosylphosphatidylinositol-anchored proteins. J Biol Chem 289:16835–16843
Thinakaran G, Koo EH (2008) Amyloid precursor protein trafficking, processing, and function. J Biol Chem 283:29615–29619
Travers KJ, Patil CK, Wodicka L, Lockhart DJ, Weissman JS, Walter P (2000) Functional and genomic analyses reveal an essential coordination between the unfolded protein response and ER-associated degradation. Cell 101:249–258
Vetrivel KS, Gong P, Bowen JW, Cheng H, Chen Y, Carter M, Nguyen PD, Placanica L, Wieland FT, Li YM, Kounnas MZ, Thinakaran G (2007) Dual roles of the transmembrane protein p23/TMP21 in the modulation of amyloid precursor protein metabolism. Mol Neurodegener 2:4
Vetrivel KS, Kodam A, Gong P, Chen Y, Parent AT, Kar S, Thinakaran G (2008) Localization and regional distribution of p23/TMP21 in the brain. Neurobiol Dis 32:37–49
Wada I, Rindress D, Cameron PH, Ou WJ, Doherty JJ 2nd, Louvard D, Bell AW, Dignard D, Thomas DY, Bergeron JJ (1991) SSR alpha and associated calnexin are major calcium binding proteins of the endoplasmic reticulum membrane. J Biol Chem 266:19599–19610
Wang H, Kazanietz MG (2010) p23/Tmp21 differentially targets the Rac-GAP beta2-chimaerin and protein kinase C via their C1 domains. Mol Biol Cell 21:1398–1408
Wang H, Xiao L, Kazanietz MG (2011) p23/Tmp21 associates with protein kinase Cdelta (PKCdelta) and modulates its apoptotic function. J Biol Chem 286:15821–15831
Wang X, Yang R, Jadhao SB, Yu D, Hu H, Glynn-Cunningham N, Sztalryd C, Silver KD, Gong DW (2012) Transmembrane emp24 protein transport domain 6 is selectively expressed in pancreatic islets and implicated in insulin secretion and diabetes. Pancreas 41:10–14
Xie J, Yang Y, Li J, Hou J, Xia K, Song W, Liu S (2014) Expression of tmp21 in normal adult human tissues. Int J Clin Exp Med 7:2976–2983
Yu X, Breitman M, Goldberg J (2012) A structure-based mechanism for Arf1-dependent recruitment of coatomer to membranes. Cell 148:530–542
Zhang L, Volchuk A (2010) P24 Family Type 1 Transmembrane Proteins are Required for Insulin Biosynthesis and Secretion in Pancreatic Beta-Cells. FEBS Lett 584:2298–2304
Zhao P, Metcalf M, Bunnett NW (2014) Biased signaling of protease-activated receptors. Front Endocrinol (Lausanne) 5:67
Zimmermann P, Hirsch-Hoffmann M, Hennig L, Gruissem W (2004) GENEVESTIGATOR. Arabidopsis microarray database and analysis toolbox. Plant Physiol 136:2621–2632
Acknowledgments
F.A. and M.J.M. were supported by the Ministerio de Economía y Competitividad (grant no. BFU2012_33883) and Generalitat Valenciana (ISIC/2013/004, ACOMP/2014/202, and FPA/2015/074). J.C.M. and N.P. were recipients of a fellowship from Ministerio de Educación (FPU program).
Conflict of interest
The authors declare that they have no conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling Editor: David Robinson
Rights and permissions
About this article

Cite this article
Pastor-Cantizano, N., Montesinos, J.C., Bernat-Silvestre, C. et al. p24 family proteins: key players in the regulation of trafficking along the secretory pathway. Protoplasma 253, 967–985 (2016). https://doi.org/10.1007/s00709-015-0858-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00709-015-0858-6