
Abstract
In this paper, we explored the protective effect and physiochemical mechanism of He-Ne laser preillumination in enhancement of tall fescue seedlings tolerance to high salt stress. The results showed that salt stress greatly reduced plant growth, plant height, biomass, leaf development, ascorbate acid (AsA) and glutathione (GSH) concentration, the enzymatic activities, and gene expression levels of antioxidant enzymes such as catalase (CAT) and glutathione reductase (GR) and enhanced hydrogen peroxide (H2O2) content, superoxide radical (O2 ·−) generation rates, membrane lipid peroxidation, relative electrolyte leakage, the enzymatic activities, and gene expression levels of superoxide dismutase (SOD), ascorbate peroxidase (APX), and peroxidase (POD), compared with controls. However, He-Ne laser preillumination significantly reversed plant growth retardation, biomass loss, and leaves development decay induced by salt stress. And the values of the physiochemical parameters observed in salt-stressed plants were partially reverted or further increased by He-Ne laser. Salt stress had no obvious effect on the transcriptional activity of phytochromeB, whereas He-Ne laser markedly enhanced its transcriptional level. Preillumination with white fluorescent lamps (W), red light (RL) of the same wavelength, or RL, then far-red light (FRL) had not alleviated the inhibitory effect of salt stress on plant growth and antioxidant enzymes activities, suggesting that the effect of He-Ne laser on improved salt tolerance was most likely attributed to the induction of phytochromeB transcription activities by the laser preillumination, but not RL, FRL or other light sources. In addition, we also utilized sodium nitroprusside (SNP) as NO donor to pre-treat tall fescue seedlings at the same conditions, and further evaluated the differences of physiological effects between He-Ne laser and NO in increasing salt resistance of tall fescue. Taken together, our data illustrated that He-Ne laser preillumination contributed to conferring an increased tolerance to salt stress in tall fescue seedlings due to alleviating oxidative damage through scavenging free radicals and inducing transcriptional activities of some genes involved in plant antioxidant system, and the induction of phytochromeB transcriptional level by He-Ne laser was probably correlated with these processes. Moreover, this positive physiochemical effect seemed more effective with He-Ne laser than NO molecule.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Tall fescue (Festuca arundinacea Schreb.) is the predominant, perennial, cool-season turf and forage grass species grown or planted under diverse environmental conditions (Dong and Qu 2005). It is widely used for general purpose turf and low-maintenance grass cover and plays an important role in environmental protection and livestock production (Wang and Ge 2005; Ma et al. 2014). Accordingly, an increased tolerance to unfavorable environmental conditions such as drought, high soil salinity, extreme temperature, ultraviolet-B radiation, high light, air pollution, heavy metals, or combination of these conditions appears indispensable for tall fescue growth and development (Zsigmond et al. 2012). These extreme environmental conditions usually lead to abiotic stresses and the accumulation of reactive oxygen species (ROS) in plant cells, including singlet oxygen (O1/2), superoxide radical (O2 ·−), hydrogen peroxide (H2O2), and hydroxyl radical (HO·) (Moller and Sweetlove 2010; Zsigmond et al. 2012). The excessive amounts of ROS formation are considered to be harmful for plant cells, which will produce oxidative stress for plants resulting in the oxidization of cellular components, macromolecules such as DNA and protein, carbohydrates, lipids, and chlorophyll (Zsigmond et al. 2012). Thus, cellular damage and cell death will become inevitable. To control the low level of ROS and protect plant cells against abiotic stresses, plants always have evolved a brand spectrum of antioxidant defense mechanisms and cope with ROS production through the use of the certain radical-scavenger systems, including low molecular weight antioxidants such as ascorbate acid (AsA), reduced glutathione (GSH), carotenoids, α-tocopherol, and flavonoids, as well as antioxidant enzymes such as superoxide dismutase (SOD: EC 1.15.1.1), catalase (CAT: EC 1.11.1.6), peroxidase (POD: EC 1.11.1.7), ascorbate peroxidase (APX: EC 1.11.1.11) and NADPH-dependent GSSG reductase (GR: EC 1.6.4.2) (Mittler 2002; Apel and Hirt 2004; Sun et al. 2007). Since these ROS scavengers can react with superoxide radical or H2O2, and then produce H2O and O2 (Ramachandra et al. 2004; Sharma and Dietz 2009; Triantaphylidès and Havaux 2009), the toxicity effects for plants due to the excessive production of ROS will be detoxified. Therefore, high activities of antioxidant enzymes and high concentration of antioxidants are advantageous for plants against many environmental stressors, and can confer an enhanced tolerance to unfavorable growth conditions in plants.
Soil and water salinity are the major environmental stress that drastically decline crops growth and productivity in many countries and plague intensive food production with perhaps the greatest threat being to sustainable irrigated agriculture (Rengasamy 2006; Rozema and Flowers 2008; Yadav et al. 2011; Hasegawa 2013; Iqbal and Ashraf 2013). Today, soil and water salinity become more and more acute due to constantly increasing human water consumption and environmental contamination (Rozema and Flowers 2008; Hasegawa 2013). The increased salinization of arable land is expected to have devastating global effects within the next 15 years (Lima et al. 2014). Primary salt stress effects are hyperosmolality, which causes water deficit for plant growth. What is more important, the accumulation of intracellular ROS during salt stress changes the degree of lipid peroxidation, ion leakage, malondialdehyde (MDA) content, hydrogen peroxide levels, and superoxide radical generation rates, which appears to disturb the complex biochemical metabolism and physiological processes resulted in the significant decreases of plant growth and development, and if severe enough, leads to cell death. In preliminary studies, it could be overcome through traditional plant breeding or producing transgenic plants, but these biotechnological methods applied to develop salt-resistant plants or improve salt-tolerance of crops were not much successful, even after gene improvement (Dionisio-sese and Tobita 2000; Ottow et al. 2005; Bing et al. 2013; Ma et al. 2014). Then the use of some physical methods with low cost and low risk like He-Ne laser radiation for protecting plant cells against oxidative damage and enhancing plants salt tolerance could be the attractive solution to overcome the salinity problem.
He-Ne and CO2 laser, commonly employed in crops and many higher plants, play a positive role in accelerating plant growth and metabolism (Qiu et al. 2008; Qiu et al. 2010; Perveen et al. 2011; Qiu et al. 2013) and also protect plant seedlings against biotic and abiotic damages induced by a variety of unfavorable environmental stressors like fungal infection (Han et al. 2006; Dong et al. 2007), enhanced UV-B radiation (Chen 2008; Yang et al. 2012), chilling stress (Chen et al. 2010), osmotic stress (Qiu et al. 2010), drought stress (Qiu et al. 2008), and cadmium stress (Qiu et al. 2013). The laser treatment enhanced plants tolerance mainly through improving seeds germination rates, plant height, root length and numbers, seedlings biomass, enzymatic activities, and chlorophyll contents in seedlings subjected to environmental stressors (Laspina et al. 2005; Yang et al. 2012; Qiu et al. 2013). In addition, previous studies had also demonstrated that nitric oxide (NO), as a bioactive molecule involved in many physiological and biochemical processes, could protect seedlings against oxidative damage under various environmental stress forms (Laspina et al. 2005; Song et al. 2006; Shi et al. 2007; Zhao et al. 2008). However, the use of He-Ne laser radiation as a protective method against long-term high salt-induced oxidative damage has been poorly studied, including its effects on seedlings growth, leaf development, and biomass especially in salt-stressed seedlings. At present, little is known about stress-protective mechanisms of He-Ne laser, and whether He-Ne laser can protect plants through increasing enzymatic and non-enzymatic defense mechanisms to detoxify free radicals and reduce oxidative stress induced by salt stress is worthy to be further explored, then a series of experiments were carried out to test this hypothesis. We also firstly compared the differences of the physiological effects between He-Ne laser preillumination and NO pretreatment on salt-stressed plants.
It is well known that phytochromes (Phy) play an important role in the formation of plant resistance to oxidative stress and the development of stress-protective mechanisms in plants (Carvalho et al. 2011; Kreslavski et al. 2013). Among five types of Phy identified in higher plants now, phytochromeA (PhyA) and phytochromeB (PhyB) are basic types and also found in Festuca plants. PhyA is the primary photoreceptor perceiving and mediating various responses to far-red light (FRL). PhyB is the predominant phytochrome regulating responses to low energy red light (RL), including He-Ne laser (Smith 2000; Quail 2002). The content and state of the phytochrome significantly influences not only the accumulation of antioxidants with low molecular weight such as flavonoids, but also the activity of antioxidant enzymes, including peroxidase (Kreslavski et al. 2013). Thus, we presume that the phytochrome-signaling system is probably involved in the formation of stress resistance induced by He-Ne laser preillumination in salt-stressed plants. Therefore, we subsequently examined whether the transcriptional activity of PhyB, a common and important form of photoreceptors, was upregulated under a combination of He-Ne laser preillumination and salt stress.
The primary aim of this study is to evaluate the interactive effect of He-Ne laser preillumination and high salinity stress, and further elucidate the precise physiochemical mechanisms of He-Ne laser preillumination in enhancement of salt tolerance through alleviating salt-induced oxidative damage in tall fescue seedlings. And these results could potentially benefit turf-type grass breeding by improving plant growth and decreasing yield losses under high salinity conditions.
Materials and methods
Plant materials and treatment conditions
Mature seeds of tall fescue (Festuca arundinacea Schreb. Houndog5) were dehusked by stirring in 50 % sulfuric acid for 30 min and rinsed with distilled water for three times followed by 95 % ethanol, then surface sterilized with 6 % sodium hypochlorite supplemented with 0.1 % Tween-20 for 30 min and rinsed again with distilled water for five times (Wang and Ge 2005). The seeds were cultured on 1/2MS medium at a plant incubator for 3 days, at 24 ± 2 °C, 70 % relative humidity, a photoperiod 16/8 h day/night cycle. After germination, seedlings were transplanted into black plastic containers containing suitable volume of Hoagland’s nutrient solution (Xu et al. 2010). Each container contained 30 seedlings and nutrient solution was renewed once a week.
After 10 days of pre-culture, seedlings was stressed with 50, 100, and 150 mM NaCl for 10 days again, or pretreated by white fluorescent lamps (W), red light (RL), a combination of red light and far-red light (red light irradiation, then far-red light, RL + FRL), He-Ne laser (H), or SNP (S) for 7 days, respectively, prior to NaCl stress. At the same time, control seedlings were cultured in Hoagland’s nutrient solution and not treated with any stresses. SNP (Sigma, USA) was used as NO donor, and pretreatment concentration was 80, 100, and 120 μM, respectively. He-Ne laser was obtained with the He-Ne laser generator (wavelength λ = 632.8 nm, flux rate 5 mW mm−2 (Supplemental materials Figure 1 and Figure 2), initial beam diameter 2 mm and final diameter 100 mm via amplifier, made by Nanjing Laser Instrument Factory, Nanjing City, China), and the duration of irradiation was 2, 4, 6 min per day, respectively. The white fluorescent lamps (photosynthetic photo flux density (PPFD), 100 μmol m−2 s−1, 10 min), red light (λ RL = 633 nm, 10 min), or red light, then far-red light (λ RL = 633 nm, λ FRL = 720 nm, 10 min), obtained with the help of LEDs, were used as control experiments to demonstrate if the laser is responsible for the effect of improved salt tolerance. The light intensity was 100 μmol m−2 s−1 per day on the surface of leaves with different light sources. Laser and all fluorescence lamps preillumination were carried out at 06:00–06:30 a.m. in darkness to prevent the influence of stray light. To gain a better insight into the role of He-Ne laser radiation in accelerating plant growth and metabolism of tall fescue, He-Ne laser treatment alone was also applied in tall fescue seedlings.
The 20-day-old seedlings were grown in the Hoagland’s nutrient solution containing different concentrations of NaCl for another 10 days at 24 ± 2 °C, a relative humidity of 70 %, a photoperiod 16/8 h day/night cycle, and PPFD 100 μmol m−2 s−1. After 10 days of salt-stress treatment, tall fescue seedlings were harvested for further analysis.
Seedlings height and growth parameters measurements
Ten days after NaCl treatment, a total of 50 seedlings chosen randomly from each treatment were measured and the values of seedlings height, fresh weight, and dry weight were recorded. For plant growth measurement, treatments were performed after germination of 3 days, just at the early growth stages, and total coleoptile length was determined with 20-day-old seedlings. Growth was expressed as total coleoptile length/numbers of total seeds, indicating the change of growth rates of seedlings at the early growth stages. For plant height measurement, all treatments were carried out after pre-culture of 10 days followed with germination of 3 days and examined with 30-day-old seedlings from the base of the stem to the tip of the topmost fully expanded leaf of the plants using a meter stick, indicating the effect of the laser and salt stress on mature plant height.
Fresh weight was measured after just detaching the leaves from tall fescue seedlings. Dry weight was determined after drying the leaves at 80 °C in an oven for 48 h.
Leaves development determination
At the end of experiments, leaves development in response to salt stress and He-Ne laser preillumination was determined with Portable Leaf Area Meter (AM-300, ADC, UK) and indicated by leaf length, leaf width, and leaf area. Fresh leaves detached from 50 seedlings of each treatment were sampled for analysis.
Relative electrolyte leakage and lipid peroxidation measurements
Relative electrolyte leakage (REL) of tall fescue seedlings in different treatments was measured according to the method of Xu et al. (2010).
Membrane lipid peroxidation was estimated by malondialdehyde (MDA) contents in leaves of tall fescue, which was determined by the slightly modified method according to Xu et al. (2010). The fresh leaves (0.1 g) were homogenized in 1 ml 10 % trichloroacetic acid (TCA), and then the homogenate was centrifuged at 4000×g for 10 min. The incubation mixture contained 2 ml supernatant, 1 ml 0.6 % thiobarbituric acid (TBA), and the mixture was incubated in 100 °C for 15 min. The mixture was centrifuged again after cooled rapidly and the absorbance of the supernatant was measured at 532, 600 and 450 nm, respectively.
Hydrogen peroxide and superoxide radical determination
Hydrogen peroxide contents were determined by the peroxidase-coupled assay according to the method of Zhao et al. (2008).
Superoxide radical contents were measured by the slightly modified method according to Xu et al. (2010). Leaves of tall fescue (0.1 g) were homogenized with the appropriate volume of 65 mM phosphate buffer (pH 7.8), centrifuged at 10,000×g for 10 min. The whole supernatant was mixed with 1.5 ml of 60 mM phosphate buffer (pH 7.8) and 0.5 ml of 1 mM hydroxylamine hydrochloride. The mixture was incubated in 25 °C for 20 min and subsequently incubated with 2 ml of 17 mM 4-aminobenzene sulfonic acid and 2 ml of 7 mM α-naphthylamine at 30 °C for 30 min. The absorbance of the final solution was measured at 530 nm. The superoxide radical content was calculated based on the standard curve.
Antioxidant compounds concentration measurement
Extracts for glutathione (GSH) concentration measurement were prepared from fresh leaves (0.5 g) homogenized under ice-cold conditions in 3 ml extraction buffer containing 5 % sulphosalycilic acid (pH 2.0), 1 g polyvinylpyrrolidone (PVP). After centrifugation at 15,000×g for 15 min at 4 °C, the supernatants added 2.0 ml phosphate buffer (pH 7.7) and 0.5 ml 4 mM 5,5′-dithiobis(2-nitrobenzoic acid) (DTNB) was used for the following analysis. After incubation of 10 min at 25 °C, the absorbance was read at 412 nm. GSH concentration was calculated according to the standard curve (Anderson 1985).
Frozen leaves in liquid nitrogen (2.0 g) were homogenized with a mortar and pestle and extracted with 3 ml 6.5 % metaphosphoric acid containing 1 mM EDTA under ice-cold conditions. After centrifugation at 15,000×g for 15 min, the supernatant added with 1 mM DTT was used for estimation of total ascorbate (ascorbate and reduced ascorbate). The ascorbic acid (AsA) concentration was determined by a standard curve (Law et al. 1983).
Antioxidant enzymes activities assays
Leaves (1.0 g) frozen in liquid nitrogen were homogenized under ice-cold conditions in 5 ml 50 mM phosphate buffer (pH 7.8) containing 1 mM EDTA, 1 mM ascorbic acid, 1 mM DTT, and 2 % PVP. The homogenates were centrifuged at 12,000×g for 20 min at 4 °C and supernatant fraction was used for the enzymes activities assays.
Ascorbate peroxidase (APX) activity was measured immediately by monitoring the rate of ascorbate oxidation at 290 nm, as described by Nakano and Asada (1981). The 3-ml reaction mixture contained 50 mM potassium phosphate buffer (pH 7.0), 0.1 mM EDTA, 0.3 mM ascorbate, 1 mM H2O2, and 28 μl of the enzyme extract. One unit of APX was defined as the amount of enzyme that oxidized 1 μM ascorbate per min at 25 °C.
Total superoxide dismutase (SOD) activity was determined in the homogenates in terms of its ability to inhibit the photochemical reduction of nitroblue tetrazolium (NBT) (Rao and Sresty 2000). The reaction mixture included 50 mM phosphate buffer (pH 7.8), 13 mM methiony, 75 μM NBT, 0.1 mM EDTA, 2 μM riboflavin, and 100 μl of enzyme extract. The reaction was initiated by fluorescent lamp with intensity of 100 μmol m−2 s−1 for 15–20 min. After terminating the reaction, absorbance of the reaction mixture was read at 560 nm. The SOD activity was evaluated according to the amount of required that caused a 50 % inhibition of the rate of NBT photo-reduction under the assay conditions.
Peroxidase (POD) activity was assayed by following the increase in absorption at 470 nm due to the formation of oxidation of guaiacol according to the method of Zhang and Kirham (1994), using a reaction mixture containing 25 mM phosphate buffer (pH 7.0), 0.1 mM EDTA, 1.0 mM H2O2, 0.05 % guaiacol (w/v). and 20 μl enzyme extract.
Activity of catalase (CAT) was assayed as a decrease in absorbance at 240 nm for 1 min due to the decomposition of H2O2. The reaction mixture consisted of 25 mM sodium phosphate buffer (pH 7.0), 10 mM H2O2, and 100 μl enzyme extract. The reaction was initiated by adding 10 mM H2O2 (Zhang and Kirham 1994).
Activity of glutathione reductase (GR) was determined by the method of Schaedle and Bassham (1977), which depends on the rate of decrease in the absorbance of NADPH at 340 nm. The reaction solution consisted of 25 mM HEPES buffer (pH 7.0), 0.5 mM oxidized glutathione (GSSG), 0.12 mM NADPH, 0.2 mM EDTA, and 50 μl enzyme extract.
Total RNA extraction, cDNA synthesis, and real-time quantitative PCR
Total RNA from tall fescue seedlings in different treatment conditions was extracted using the Concert Plant RNA Reagent (Invitrogen, USA) according to the supplied protocol. The Oligo software was used for primer design based on the EST sequences of candidate proteins available in the NCBI. The sequences of primers were given in Table 1 and the GAPDH (glyceraldehyde-3-phosphate dehydrogenase) gene was used as an internal control.
Two micrograms of total RNA was reverse-transcribed into cDNA by RT-PCR (Biometra, TGRadient, Germany) using random hexamers and M-MLV Reverse Transcriptase (Promega, WI, USA). After reverse transcription, the products of each reaction were diluted five times to avoid potential primer interference in the following qRT-PCR reactions. Quantitative real-time PCR reaction was performed in a 20 μl of total volume containing 2 μl cDNA, 300 nM of each primer, and 10 μl of the SYBR Green PCR Master Mix (GeneCore BioTechnologies, Shanghai, China) through using a real-time PCR detection system (Model Corbett RG6000, Australia) according to the manufacturer’s instruction. The reaction cycling conditions for antioxidant enzyme genes were 95 °C for 2 min, 35 cycles at 95 °C for 15 s, 60 °C for 15 s, and 72 °C for 20 s. The qPCR conditions for PhyB gene were 95 °C for 3 min, 50 cycles at 95 °C for 30 s, 60 °C for 30 s, and 72 °C for 30 s. At the end of qPCR, dissociation curve analysis was performed to confirm the purity and specificity of amplification products. The comparative CT method was used to analyze the expression level of antioxidant enzyme genes of Sod, Pod, Cat, Gr, Apx, and phytochrome-signaling gene of PhyB (Livak and Schmittgen 2001).
Statistical analysis
Values from five replicates were expressed as means ± SD (standard deviation) and analyzed by the Student’s t test. The data in the figures or tables were considered significant difference at P < 0.05 according to Duncan’s multiple range test.
Results
Plant growth parameters measurement
In the present work, according to Fig. 1, we found that the phenotype alteration was significant in tall fescue seedlings under 150 mM NaCl treatment (Fig. 1a), and high salt-stressed plants showed a green-yellow phenotype before the two-leaf stage of tall fescue. After the two-leaf stage, the yellow leaves gradually turned green, and eventually approached normal color. After preillumination with He-Ne laser or SNP pretreatment, all leaves showed normal green color (Fig. 1b–d). And seedlings height and roots length (N + H and N + S group) were much greater than salt-stressed plants (N group), but still lower than that under normal conditions (CK group). However, preillumination of leaves with white fluorescent lamps, RL alone, or RL, then FRL had not alleviated the inhibitory effect of salt stress on plant phenotype characteristics and growth (Supplemental materials Figure 3A, B, and C). Therefore, we used He-Ne laser preillumination for subsequent experiments.

The phenotype of tall fescue seedlings under different growth conditions. a Tall fescue seedlings under different NaCl concentrations. b Tall fescue seedlings under a combination of He-Ne laser preillumination and salt stress. c Tall fescue seedlings under a combination of SNP pretreatment and salt stress. d The phenotype of tall fescue under different treatment conditions. CK control plants; N50, N100, N150 stress treatment with 50, 100, and 150 mM NaCl, respectively; N + H 2 , N + H 4 , N + H 6 He-Ne laser preillumination 2, 4, and 6 min per day prior to salt stress, respectively; N + S 80, N + S 100 , N + S 120 SNP (sodium nitroprusside) of 80, 100, and 120 μM pretreatment prior to salt stress, respectively; H the 4 min per day of He-Ne laser treatment alone without salt stress
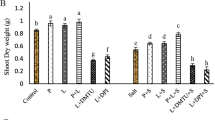
The effects of combined treatment with He-Ne laser and high salt stress on morphological characteristics of tall fescue were also evaluated by the estimation of agronomic characters, such as plant growth, biomass, and leaf development. As shown in Fig. 2, seedlings biomass (Fig. 2a–c) and plant height (Fig. 2d) significantly decreased in plants under 150 mM NaCl treatment, compared with control plants (P < 0.05). And plant growth (Fig. 2e) was also drastically inhibited. Plants pretreated with He-Ne laser or SNP prior to salt stress, however, were less affected. The 4 min per day of the laser (Fig. 2a) and 100 μM of SNP (Fig. 2b) were the most effective for the protection of tall fescue seedlings, respectively.

Growth parameters in tall fescue seedlings in response to combined treatment of He-Ne laser and salt stress. a Effects of different time of He-Ne laser preillumination on biomass (fresh weight and dry weight) in tall fescue. FW fresh weight, DW dry weight. b Effects of different SNP treatment concentration on biomass in tall fescue. c The changes of biomass in tall fescue under combined treatment of He-Ne laser or SNP and salt stress. d Seedlings height of tall fescue under combined treatment of He-Ne laser or SNP and salt stress. e Plant growth of tall fescue under combined treatment of He-Ne laser or SNP and salt stress. Values are means ± SD (n = 50), different letters followed with bars indicate significant differences at P < 0.05 according to Duncan’s multiple range test
Leaf development determination
The effects of He-Ne laser on leaves development of tall fescue are shown in Table 2. Salt stress significantly inhibited leaves development, as indicated by leaf length reduction of 54 %, leaf width reduction of 52 %, leaf area reduction of 77 %, and similar to SNP pretreatment, the inhibition was drastically alleviated by He-Ne laser preillumination (P < 0.05). But He-Ne laser treatment for non-salt stress plants had no obvious effects on leaves development.
Moreover, in this study, we found that the values of plant growth parameters and biomass of tall fescue pretreated with 4 min of He-Ne laser per day for 7 days were all slightly higher than that of plants pretreated with 100 μM SNP for 7 days (P > 0.05). In addition, under normal conditions, application of He-Ne laser did not greatly change plant growth parameters and biomass (H).
Effects of He-Ne laser preillumination on the relative electrolyte leakage and lipid peroxidation
Relative electrolyte leakage (REL) under 150 mM NaCl stress was the highest compared with other groups (Fig. 3a). And it was markedly decreased by 28.63 % with He-Ne laser preillumination prior to NaCl stress (P < 0.05). Furthermore, the decrease with He-Ne laser preillumination was slightly greater than SNP pretreatment (P > 0.05). The value of REL in the group receiving only He-Ne laser treatment was 11.97 %, similar to that of control (P > 0.05).

Effects of He-Ne laser preillumination on a relative electrolyte leakage and b malondialdehyde content (MDA) in tall fescue under salt stress. Values are means ± SD (n = 5) from five replicates. Different letters followed with bars are significantly different at P < 0.05
Since MDA content is usually used as an index of lipid peroxidation, it was measured in leaves of tall fescue under different treatments (Fig. 3b). In plants leaves from 150 mM NaCl treatment, NaCl induced an increase of 132.14 % in MDA content, as compared to controls (P < 0.05). He-Ne laser preillumination significantly alleviated the negative effects of salt stress on membrane lipid peroxidation. We also found that MDA content in plants with He-Ne laser preillumination was slightly lower than that with SNP. However, He-Ne laser treatment alone had no obvious influence on lipid peroxidation.
The levels of hydrogen peroxide and superoxide radical
According to the results of Fig. 4, H2O2 (Fig. 4a) and O2 ·− (Fig. 4b) content under salt stress increased to 172 and 146 % of the control, respectively. Our results also revealed that He-Ne laser preillumination could alleviate NaCl-induced oxidative stress by decreasing the accumulation of H2O2 and O2 ·−. In addition, under normal conditions, application of He-Ne laser had no significant effects on H2O2 and O2 ·− levels.

Effects of He-Ne laser preillumination on a levels of H2O2 and b superoxide radical (O2 ·−) in tall fescue under salt stress. Values are means ± SD (n = 5) calculated from five replicate experiments. Bars with different letters are significantly different at P < 0.05
Glutathione and ascorbic acid concentration
Compared to the control, a significant reduction was observed in concentration of GSH (Fig. 5a) and AsA (Fig. 5b) of tall fescue seedlings exposed to salt stress (P < 0.05). However, antioxidant compounds concentration remarkably increased in seedlings subjected to combined treatment (P < 0.05), especially with He-Ne laser preillumination. He-Ne laser treatment alone slightly enhanced GSH and AsA content in tall fescue.

Effects of He-Ne laser preillumination and salt stress on a glutathione (GSH) and b ascorbic acid (AsA) concentration in tall fescue. After 10 days of NaCl treatment, the first pair of fully expanded leaves was used for determination. Values are means ± SD (n = 5) from five replicates, and different letters followed with bars indicate significant differences (P < 0.05)
Antioxidant enzyme activities
In tall fescue seedlings under 150 mM NaCl stress, the production of ROS is the highest compared to other groups, and that not only depends on the lower soluble antioxidants contents, but also is related with the certain antioxidant enzyme activities. Therefore, activities of ROS-scavenging enzymes including SOD, POD, APX, CAT, and GR were determined under salt stress with or without He-Ne laser preillumination.
Table 3 illustrated the effects of 150 mM NaCl treatment on SOD, APX, POD, CAT, and GR activities in leaves of tall fescue, either with or without He-Ne laser preillumination or SNP pretreatment. Under salt stress conditions, activities of SOD, APX, and POD significantly increased by 70.64, 51.56, and 144.33 % in tall fescue seedlings. But He-Ne laser preillumination increased three enzymatic activities even more, 141.28, 66.41, and 195.07 % over the controls, respectively. No markedly changes compared with the control were observed when plants were treated with He-Ne laser alone, except for APX activity, but only 44.33 % over the control. The CAT activity was strongly inhibited under salt stress. However, compared with salt-stressed plants, He-Ne laser preillumination or SNP pretreatment could significantly increase CAT activity. NaCl treatment reduced GR activity, while He-Ne laser preillumination or SNP pretreatment also restored GR activity. Moreover, plants treated with He-Ne laser alone showed no significant change in activities of CAT and GR compared to control values.
Transcriptional levels of SOD, POD, CAT, GR, APX, and PhyB
As shown in Fig. 6, salt stress significantly induced a decrease in expression levels of CAT and GR genes, whereas an increase in transcript levels for POD, SOD, and APX genes. He-Ne laser preillumination significantly upregulated (P < 0.05) expression levels of all genes of SOD, POD, APX, CAT, and GR as compared with salt stressed seedlings (Fig. 6 N + H). These results suggested that enhanced gene expression was contributed to the increases in the activities of antioxidant enzymes, thus resulting in the protection against oxidative damage induced by salt stress.

Effects of He-Ne laser on antioxidant enzymes of superoxide dismutase (SOD), peroxidase (POD), catalase (CAT), glutathione reductase (GR), ascorbate peroxidase (APX), and PhytochromeB (PhyB) gene expression level in tall fescue under 150 mM NaCl stress. The relative transcriptional levels of SOD, POD, CAT, GR, APX, and PhyB taking control as 1 U. Bars are means ± SD (n = 5), different letters within column indicate significant differences (P < 0.05) according to Duncan’s multiple range test
In addition, the laser alone also slightly increased the transcript level of these genes as compared to the control. Our data implied that the laser preillumination had a positive role in increasing salt tolerance in plants under high saline conditions.
Salt stress had no significant effects on transcript levels of PhyB in leaves of tall fescue (Fig. 6f). However, He-Ne laser radiation greatly enhanced the transcript activity of PhyB in plants both under salt stress and normal condition (N + H and H group), suggesting the PhyB of higher transcriptional level induced by He-Ne laser was probably conductive to the improved salt tolerance due to its enhancement effect on antioxidants concentrations and antioxidant enzymes activities. For N + S group, transcript activity of PhyB was also similar with controls (CK).
To further make sure whether PhyB induced by the laser related with enhancement of salt tolerance, antioxidant enzymes activities were also determined in tall fescue seedlings preillumination with white fluorescent lamps, RL alone, or RL, then FRL. And the results demonstrated that preillumination with white fluorescent lamps, RL alone, or RL, then FRL had not increased the activities of these antioxidant enzymes compared to salt-stressed plants (Supplemental materials Table 1), suggesting that antioxidant enzymes genes and PhyB gene were not upregulated by white fluorescent lamps, RL alone, or RL, then FRL, despite PhyB was the red light photoreceptors. These results further implied that the induction of higher content of PhyB by He-Ne laser might have a link with improved salt tolerance induced by the laser.
Discussion
It is well known that the reactive oxygen species (ROS) formation will elevate in plant cells when exposed to various stressful conditions, including high salt stress (Laspina et al. 2005; Shi et al. 2007; Zhao et al. 2008; Xu et al. 2010), which leads to oxidative damage to the plants. Previous studies have reported that He-Ne laser, like NO molecule, could protect plant cells against cadmium-induced oxidative stress by detoxifying ROS (Beligni and Lamattina 2002; Qiu et al. 2013). However, there is still limited information about the morphological characteristics and physiochemical responses of plants to combined treatment with He-Ne laser preillumination and high salt stress.
The effects of combined treatment with He-Ne laser and high salt stress on morphological characteristics of tall fescue were significant. The 4 min per day of the laser (Figs. 1b, 2a) and 100 μM of SNP (Figs. 1c, 2b) were the most effective for the protection of tall fescue seedlings under salt stress, respectively. These results were in accordance with the results of Shi et al.(2007), who reported that NaCl stress reduced significantly the dry weight of both shoot and root of cucumber plants and the inhibition was alleviated by exogenous NO treatment, but in that case the use of lower SNP concentration (50 μM). However, in our study, preillumination of leaves with white fluorescent lamps, RL alone, or RL, then FRL had not alleviated the inhibitory effect of salt stress on plant growth and development (Supplemental materials Figure 3A, B and C). Therefore, the improved salt tolerance of seedlings should be attributed to He-Ne laser irradiation.
Since MDA content is usually used as an index of lipid peroxidation, it was measured as a biomaker of salt-induced oxidative stress (Fig. 3b). The results showed that He-Ne laser preillumination significantly alleviated the negative effects of salt stress on membrane lipid peroxidation. Therefore, He-Ne laser apparently acts as an efficient ROS scavenger or a membrane stabilizer. Since oxidative stress is generated through the formation of reactive oxygen species (ROS) in plants, hydrogen peroxide, and superoxide radical would remarkably accumulate in plant cells subjected to salt stress. In our study, the significant accumulation of H2O2 and O2 ·− in cells indicated that oxidative stress was induced by high concentration of NaCl in tall fescue seedlings. He-Ne laser preillumination could alleviate NaCl-induced oxidative stress by decreasing the accumulation of H2O2 and O2 ·−, and this result was in agreement with the observations of Qiu et al. in studying the role of He-Ne laser pretreatment on enhancing wheat seedlings tolerance to cadmium and osmotic stress (Qiu et al. 2010; Qiu et al. 2013). Under salt stress conditions, increased H2O2 content, the rate of O2 ·− generation and lipid peroxidation had been also reported in mitochondria of pea leaves, tomato leaves and roots, and cucumber roots (Gómez et al. 1999; Foissner et al. 2000; Mittova et al. 2004; Shi et al. 2007).
Plants have evolved a complex antioxidant system, such as antioxidant compounds and antioxidant enzymes, to avoid the harmful or negative effects of ROS on cellular constituents. Glutathione (GSH) and ascorbic acid (AsA) are the major antioxidants in protecting plants against oxidative damage due to the ROS formation, which can detoxify ROS by reacting with singlet oxygen, hydrogen superoxide, superoxide radical, OH−, and lipid hydroperoxide (Foyer and Halliwell 1976; Ramachandra et al. 2004). All together, in our study, the lower GSH and AsA concentration lead to the accumulation of H2O2 and O2 ·−, which resulted in oxidative stress for plants or plant cells. While He-Ne laser preillumination significantly alleviated cell injury caused by salt stress through increasing the supply or biosynthesis rate of these antioxidant compounds to detoxify excessive amounts of ROS production. Panda et al. (2011) and Qiu et al. (2013) have also reported that the higher GSH and AsA levels would contribute to elevating plants tolerance to cadmium-induced oxidative stress.
In tall fescue seedlings under 150 mM NaCl stress, the production of ROS is the highest compared to other groups, and that not only depends on the lower soluble antioxidants contents but also is related with the certain antioxidant enzyme activities. Therefore, activities of ROS-scavenging enzymes including SOD, POD, APX, CAT, and GR were determined under salt stress with or without He-Ne laser preillumination. Shi et al. (2007) have reported that the influence of salt stress on the antioxidant enzymes is related to the plant treatment time, plant tissues, plant species, and genotypes. The different influence of long-term salt stress on different ROS-scavenging enzymes activities had also been found in the present experiment.
It has been reported that ROS-scavenging enzymes activities markedly increased in cotton callus tissues (Vital et al. 2008), shoot cultures of rice (Fadzilla et al. 1997), and cucumber root (Shi et al. 2007) under saline conditions. Our results found that activities of SOD, POD, and APX would increase in tall fescue under NaCl treatment conditions, which implied that the plant antioxidant machinery might effectively struggle against salt stress. However, catalase (CAT) and glutathione reductase (GR) activities were inhibited or reduced by salt stress. The results were also in accordance with the results of Meneguzzo et al. (1999) in observations of wheat roots antioxidant enzymes response to increasing NaCl concentration. And these observations of our experiments further suggested that the influences of salt stress on the antioxidant enzymes activities are very complex. Taken together, since SOD converts the superoxide anion into H2O2, and CAT and other several peroxidases like APX then eliminated H2O2 production by conversion to H2O and O2, the high SOD and low CAT activities will lead to a high H2O2 accumulation, which exceeds APX detoxification capability despite APX activity increase greatly under salt stress. This result was supported by the reports of Laspina et al. (2005). He-Ne laser preillumination strongly enhanced values of SOD, POD, APX, CAT, and GR compared with the control. Furthermore, the values of antioxidant compounds contents and ROS-scavenging enzymes activities with 4 min per day of He-Ne laser preillumination for 7 days were all slightly higher than that with 100 μM SNP as NO donor pretreatment for 7 days in leaves of tall fescue. Therefore, the He-Ne laser preillumination can remarkably alleviate oxidative damage in plants by strongly increasing antioxidant compounds contents and ROS-scavenging enzymes activities.
Although we have shown that one of the effect of the laser on oxidative stress induced by salt stress is protected by its ability to increase antioxidant enzymes activities, little has been known about the expression alteration of antioxidant enzymes at the transcriptional levels. Figure 6 revealed that He-Ne laser preillumination significantly upregulated (P < 0.05) expression level of the genes SOD, POD, APX, CAT, and GR as compared with salt-stressed seedlings (Fig. 6, N + H). These results suggested that enhanced antioxidant enzymatic gene expression has contributed to the increases in the activities of antioxidant enzymes, thus resulting in the protection against oxidative damage induced by salt stress.
In addition, to further elucidate physiochemical mechanisms of He-Ne laser against oxidative stress induced by salt stress, we also examined the transcriptional activity of phytochromeB under the laser treatment. The enhanced transcriptional level of the phytochromeB chromophore induced by the laser was supposed to have a link with the increased contents of low molecular antioxidants (AsA and GSH, Fig. 5) and the activities of antioxidant enzymes (SOD, POD, APX, CAT, and GR, Table 3). These results have been supported by several studies (Kreslavski et al. 2012; Kreslavski et al. 2013). In addition, preillumination with white fluorescent lamps, RL alone, or RL, then FRL, but not He-Ne laser, had not increased the activities of antioxidant enzymes through enhancement of transcriptional levels of antioxidant enzymes and PhyB genes compared to salt-stressed plants in our control experiments (Supplemental materials Table 1). So we concluded that the protective effect of the laser against oxidative damage probably correlated with the induction of higher content of PhyB by He-Ne laser via an unknown mechanism (Fig. 7). Thus, the induction of higher content of PhyB by the laser was presumed to be contributed to improved salt tolerance. However, the detailed mechanism of the interactions between its protective effect and expression patterns of more genes involved in phy-signaling system, as well as the induction of active form of PhyB and antioxidants biosynthesis capacity, antioxidant enzymes activities, needs further be investigated in the subsequent experiment by using some respective mutants of rice or Arabidopsis, such as the hy2 mutants of Arabidopsis with the reduced synthesis of phyB chromopore.

Models for the regulation mechanisms of He-Ne laser-induced resistance of seedlings to saline conditions. An unknown receptor protein in the plasma membrane or cell wall was involved in the laser preillumination signal transduction to increase the transcriptional levels of genes (such as SOD, POD, APX, CAT, GR, and PhyB genes) that are involved in the modulation of antioxidant enzyme activities and antioxidant compounds’ biosynthesis. The activities of antioxidant enzymes and antioxidants concentration increased significantly, which caused a decrease of oxidative stress induced by salt stress. The inhibitory effect of plant growth was partially alleviated, thereby improving the resistance of seedling to salt stress. Another reason is that seedlings absorb more energy from low density of the laser than the controls, and the active form of PhyB formation processes is triggered due to the transformation of light energy to chemical energy in plants, and resulted in the enhancement of antioxidant enzyme activities and antioxidants biosynthesis capacity
The positive effects of laser on organisms have been widely studied in various plants, and the effects of laser on plants are mainly light/optical effects, electromagnetic effects, temperature effects, and pressure effects. However, the low-power laser of visible wavelength usually generates little heat and pressure. Qiu et al. (2008, 2013) found that the light/optical effects of the laser had no obvious repair effect on plants damage. Chen (2008) reported that the effect mechanism of the laser on the organisms is most likely attributed to its electromagnetic effect. The electromagnetic effect can influence the structures and activities of biological macromolecules, such as proteins and enzymes, nucleic acid, lipids, and various physiologically active substances including GSH and AsA. Some researchers have reported that the low power of He-Ne laser can activate many enzymes in plant cells, and most of these enzymes combined with some metal cationic, such as Fe2+, Mg2+, and Mn2+. The magnetic effects of the laser commonly interact with metal cationic and promote enzymes’ activities. It is assumed that the activation effects of laser on enzymes is possibly related with paramagnetism of metal atoms lies in enzyme structure, semiconductor character of enzyme, and transmission traits of many charged ions in biochemical reactions catalyzed by enzymes.
Another possible reason is that the physiologically active form of phytochromes (Phy), especially phytochromeB (PhyB), which is formed upon radiation with the laser of low power, can enhance the activities of antioxidant enzymes and increase the contents of antioxidant compounds. This hypothesis about the mechanism of the action of low energy laser has been confirmed in some studies. Kreslavski et al. (2013) reported that Phy is the key photoreceptors involved in many processes of photomorphogenesis, which is formed upon illumination with low-energy visible red light (RL), whereas PhyB is the predominant phytochrome regulating responses to RL. Hernandez et al. (2010) and Qiu et al. (2013) assumed that seeds or seedlings pretreated with low-power laser will absorb more energy from the surroundings than the control. The laser energy triggers physiological and biochemical processes in plants due to the transformation of light energy to chemical energy. Then the activation effect of He-Ne laser on enzymes and other active substances is most likely correlated with the physiologically active form of PhyB formation by the use of more chemical energy from He-Ne laser energy (Fig. 7).
Consequently, in our study, the physiological processes and biochemical metabolism in plants preillumination with He-Ne laser for 4 min (N + H) were accelerated, and finally, plants growth was enhanced, and plant height, biomass (Figs. 1 and 2), and leaf development (Table 2) were augmented notably (P < 0.05) due to the enhancement of antioxidant compounds biosynthesis capacity and antioxidant enzymes activities induced by He-Ne laser preillumination, partially by activating transcriptional levels of some antioxidant enzymes. Furthermore, the higher content of the physiologically active form of phytochromeB induced by the laser illumination was also most likely involved in these processes. Investigation of plant response to He-Ne laser preillumination under salt stress conditions could be conducive to the understanding of plant physiological response mechanisms to light (in the laser, red, and far-red region of visible spectrum) treatment. In conclusion, we speculate that the biological effect of low-power laser is not single but a compound function of many factors.
References
Anderson ME (1985) Determination of glutathione and glutathione disulfide in biological samples. Methods Enzymol 113:548–554
Apel K, Hirt H (2004) Reactive oxygen species: metabolism, oxidative stress and signal transduction. Annu Rev Plant Biol 55:373–399
Beligni MV, Lamattina L (2002) Nitric oxide interferes with plant photo-oxidative stress by detoxifying reactive oxygen species. Plant Cell Environ 25:737–748
Bing L, Feng CC, Li JL, Li XX, Zhao BC, Shen YZ, Huang ZJ, Ge RC (2013) Overexpression of the AtSTK gene increases salt, PEG and ABA tolerance in Arabidopsis. J Plant Biol 56:375–382
Carvalho RF, Campos ML, Azevedo RA (2011) The role of phytochrome in stress tolerance. J Plant Biol 53:920–929
Chen YP (2008) Isatis indigotica seedlings derived from laser stimulated seeds showed improved resistance to elevated UV-B. Plant Growth Regul 55:73–79
Chen YP, Jia JF, Yue M (2010) Effect of CO2 laser radiation on physiological tolerance of wheat seedlings exposed to chilling stress. Photochem Photobiol 86:600–605
Dionisio-sese MD, Tobita S (2000) Effects of salinity on sodium content and photosynthetic responses of rice seedlings differing in salt tolerance. J Plant Physiol 157:54–58
Dong SJ, Qu RD (2005) High efficiency transformation of tall fescue with Agrobacterium tumefaciens. Plant Sci 168:1453–1458
Dong SJ, Tredway LP, Shew HD, Wang GL, Sivamani E, Qu RD (2007) Resistance of transgenic tall fescue to two major fungal diseases. Plant Sci 173:501–509
Fadzilla NM, Finch RP, Burdon RH (1997) Salinity, oxidative stress and antioxidant responses in shoot cultures of rice. J Exp Bot 48:325–331
Foissner I, Wendehenne D, Langebartels C, Dumer J (2000) In vivo imaging of an elicitor-induced nitric oxide burst in tobacco. Plant J 23:817–824
Foyer CH, Halliwell B (1976) The presence of glutathione and glutathione reductase in chloroplasts: a proposed role in ascorbic acid metabolism. Planta 133:21–25
Gómez JM, Hernández JA, Jiménez A, Delrio LA, Sevilla F (1999) Differential response of antioxidative system of chloroplasts and mitochondria to long-term NaCl stress of pea plants. Free Radic Res 31:11–18
Han YH, Bonos SA, Clarke BB, Meyer WA (2006) Inheritance of resistance to gray leaf spot disease in perennial ryegrass. Crop Sci 46:1143–1148
Hasegawa PM (2013) Sodium (Na+) homeostasis and salt tolerance of plants. Environ Exp Bot 92:19–31
Hernandez AC, Dominiquez PA, Cruz OA, Ivanov R, Carballo CA, Zepeda BR (2010) Laser in agriculture. Int Agrophys 24:407–422
Iqbal M, Ashraf M (2013) Gibberellic acid mediated induction of salt tolerance in wheat plants: growth, ionic partitioning, photosynthesis, yield and hormonal homeostasis. Environ Exp Bot 86:76–85
Kreslavski VD, Shirshikova GN, Lyubimov VY, Shmarev AN, Boutanaev AM, Kosobryukhov AA, Schmitt FJ, Friedrich T, Allakhverdiev SI (2013) Effect of preillumination with red light on photosynthetic parameters and oxidant-/antioxidant balance in Arabidopsis thaliana in response to UV-A. J Photochem Photobiol B Biol 127:229–236
Kreslavski VD, Khristin MS, Shabnova NL, Lyubimov VY (2012) Preillumination of excised spinach leaves with red light increases the resistance of photosynthetic apparatus to UV radiation. Russ J Plant Physiol 59:717–723
Laspina NV, Groppa MD, Tomaro MT, Benavides MP (2005) Nitric oxide protects sunflower leaves against Cd-induced oxidative stress. Plant Sci 169:323–330
Law MY, Charles SA, Halliwell B (1983) The effect of hydrogen peroxide and paraquat on glutathione and ascorbic acid in spinach (spinacia oleracea) chloroplast. Biochem J 210:899–903
Lima RB, Santos TB, Vieira LGE, Ferrarese MLL, Ferrarese-Filho O, Donatti L, Boeger MRT, Petkowicz CLO (2014) Salt stress alters the cell wall polysaccharides and anatomy of coffee (Coffea arabica L.) leaf cells. Carbohydr Polym 112:686–694
Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real-time quantitative PCR and the 2−△△Ct method. Methods 25:402–408
Ma DM, Xu WR, Li HW, Jin FX, Guo LN, Wang J, Dai HJ, Xu X (2014) Co-expression of the Arabidopsis SOS genes enhances salt tolerance in transgenic tall fescue (Festuca arundinacea Schreb.). Protoplasma 251:219–231
Meneguzzo S, Navari-Izzo F, Izzo R (1999) Antioxidative responses of shoots and roots of wheat to increasing NaCl concentration. J Plant Physiol 155:274–280
Mittova V, Guy M, Tal M, Volokita M (2004) Salinity up-regulates the antioxidative system in root mitochondria and peroxisomes of the wild-tolerant tomato species Lycopersicon pennellii. J Exp Bot 55:1105–1113
Mittler R (2002) Oxidative stress, antioxidant and stress tolerance. Trends Plant Sci 7:405–410
Moller IM, Sweetlove LJ (2010) ROS signaling-specificity is required. Trends Plant Sci 15:370–374
Nakano Y, Asada K (1981) Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplast. Plant Cell Physiol 22:867–880
Ottow EA, Brinker M, Teichmann T, Fritz E, Kaiser W, Brosché M, Kangasjärvi J, Jiang XN, Polle A (2005) Populus euphratica displays apoplastic sodium accumulation, osmotic adjustment by decrease in calcium and soluble carbohydrates, and develops leaf succulence under salt stress. Plant Physiol 139:1762–1772
Panda P, Nath S, Chanu TT, Sharma GD, Panda SK (2011) Cadmium stress-induced oxidative stress and role of nitric oxide in rice (Oryza sativa L.). Acta Plant Physiol 33:1737–1747
Perveen R, Jamil Y, Ashraf M, Ali Q, Iabal M, Ahmad MR (2011) He-Ne laser induced improvement in biochemical, physiological, growth and yield characteristics in sunflower (Helianthus annuus L.). Photochem Photobiol 87:1453–1463
Qiu ZB, Liu X, Tian XJ, Yue M (2008) Effects of CO2 laser pretreatment on drought stress resistance in wheat. J Photochem Photobiol B Biol 90:17–25
Qiu ZB, Li JT, Yue M (2010) The damage repair role of He-Ne laser on wheat exposed to osmotic stress. Can J Plant Sci 90:691–698
Qiu ZB, Li JT, Zhang MM, Bi ZZ, Li ZL (2013) He-Ne laser pretreatment protects wheat seedlings against cadmium-induced oxidative stress. Ecotoxicol Environ Saf 88:135–141
Quail PH (2002) Phytochrome photosensory signaling networks. Nat Rev Mol Cell Biol 3:85–93
Ramachandra RA, Chaitanya KV, Vivekanandan M (2004) Drought-induced responses of photosynthesis and antioxidant metabolism in higher plants. J Plant Physiol 161:1189–1202
Rao KVM, Sresty TVS (2000) Antioxidant parameters in the seedlings of pigeon pea (Cajanus cajan L. Millspaugh) in response to Zn and Ni stress. Plant Sci 157:113–128
Rengasamy P (2006) World salinization with emphasis on Australia. J Exp Bot 57:1017–1023
Rozema J, Flowers T (2008) Crops for a salinized world. Science 322:1478–1480
Schaedle M, Bassham JA (1977) Chloroplast glutathione reductase. Plant Physiol 59:1011–1012
Sharma SS, Dietz KJ (2009) The relationship between metal toxicity and cellular redox imbalance. Trends Plant Sci 14:43–50
Shi QH, Ding F, Wang XF, Wei M (2007) Exogenous nitric oxide protect cucumber roots against oxidative stress induced by salt stress. Plant Physiol Biochem 45:42–550
Smith H (2000) Phytochrome and light signal perception by plants-an emerging synthesis. Nature 407:585–591
Song LL, Ding W, Zhao MG, Sun BT, Zhang LX (2006) Nitric oxide protects against oxidative stress under heat stress in the calluses from two ecotypes of reed. Plant Sci 171:449–458
Sun BT, Jing Y, Chen KM, Song LL, Chen FJ, Zhang LX (2007) Protective effect of nitric oxide on iron defficiency-induced oxidative stress in maize (Zea mays). J Plant Physiol 164:536–543
Triantaphylidès C, Havaux M (2009) Singlet oxygen in plants: production, detoxification and signaling. Trends Plant Sci 14:219–228
Vital SA, Fowler RW, Virgen A, Gossett DR, Banks SW, Rodriguez J (2008) Opposing roles for superoxide and nitric oxide in the NaCl stress-induced upregulation of antioxidant enzyme activities in cotton callus tissue. Environ Exp Bot 62:60–68
Wang ZY, Ge YX (2005) Agrobacterium-mediated high efficiency transformation of tall fescue (Festuca arundinacea). J Plant Physiol 162:103–113
Xu YF, Sun XL, Jin JW, Zhou H (2010) Protective effect of nitric oxide on high light-induced oxidative damage in leaves of tall fescue. J Plant Physiol 167:512–518
Yadav S, Irfan M, Ahmad A, Hayat S (2011) Causes of salinity and plant manifestations to salt stress: a review. J Environ Biol 32:667–685
Yang LY, Han R, Sun Y (2012) Damage repair effect of He-Ne laser on wheat exposed to enhanced ultraviolet-B radiation. Plant Physiol Biochem 57:218–221
Zhao L, He JX, Wang XM, Zhang LX (2008) Nitric oxide protects against polyethylene glycol-induced oxidative damage in two ecotypes of reed suspension cultures. J Plant Physiol 165:182–191
Zhang JX, Kirham MB (1994) Drought stress-induced changes in activities of superoxide dismutase, catalase and peroxidase in wheat species. Plant Cell Physiol 35:785–791
Zsigmond L, Szepesi Á, Tari I, Rigó G, Király A, Szabados L (2012) Over expression of the mitochondrial PPR40 gene improves salt tolerance in Arabidopsis. Plant Sci 182:87–93
Acknowledgments
This research was supported by the National Nature Science Foundation of the People’s Republic of China (grant no. 31301245), Shanxi Province Nature Science Foundation (grant no. 2014011028-5), and the Nature Science Foundation of Shanxi Normal University (grant no. ZR1414). The authors are grateful to the editors and reviewers for their helpful comments on earlier versions of the manuscript.
Conflict of interest
The authors declare that they have no conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling Editor: Peter Nick
Electronic supplementary material
Below is the link to the electronic supplementary material.
ESM 1
(DOC 3228 kb)
Rights and permissions
About this article

Cite this article
Gao, LM., Li, YF. & Han, R. He-Ne laser preillumination improves the resistance of tall fescue (Festuca arundinacea Schreb.) seedlings to high saline conditions. Protoplasma 252, 1135–1148 (2015). https://doi.org/10.1007/s00709-014-0748-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00709-014-0748-3