
Abstract
In this study, 150 cornelian cherry accessions originated from natural population on Stara Planina Mountain (Serbia) were characterized using a set of 31 morphological traits. Besides its characterizing and quantifying the variability of its morphological traits, the objective of this study was to identify intra- and inter-population variability and relationships among the accessions and to detect the most useful variables for discrimination. The evaluation was carried out over a period of 3 years. In order to evaluate the phenotypic variability, descriptive statistics, principal components analysis (PCA) and cluster analysis were used. Coefficient of variation (CV) showed the highest values for the number of flower buds per fruiting twig, while the lowest CV being evident in traits involving flowers and stones. PCA shows high discrimination capabilities of variables measured. Most of these variables were fruit and stone characteristics. PCA also evaluated the compactness and the peculiarity of each accession, thus providing information on the mixing level among them within the space of their morphological traits. Cluster analysis revealed grouping of accessions in three main clusters, where clusters with large number of accessions undergo sub-clustering. Separation into clusters and sub-clusters was not in a function of a locality or altitude, but in a function of genotypes. The results of this study indicate that these accessions must be conserved as valuable genetic resources determine and characterize its biodiversity. Also, it was found very important to enrich the cornelian cherry gene pool in order to find several valuable well-adapted genotypes suitable for the production.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Genus Cornus L. (commonly known as dogwoods) contains about 45 species distributed throughout the regions of the northern hemisphere (Murrell 1993). Most species used as ornamentals, and only a few species are grown for their fruits, such as Cornus mas L., cornelian cherry, due to its better fruit characteristics (Seeram et al. 2002). This species primarily grows in the temperate zone of Eurasia, and it is highly tolerant to diverse abiotic and biotic conditions. It ranged from a shrub to a small tree with edible olive-shaped fruits. Fruits of this species mainly used for food, medicine, ornamental and honey plant (Mamedov and Craker 2002). Fruits are typically red whit a sweet–sour taste and rich in tannins, sugar and organic acid (Güleryüz et al. 1998; Demir and Kalyoncu 2003; Ercisli et al. 2011), as well as phenol, ascorbic acid and anthocyanin (Rop et al. 2010; Hassanpour et al. 2011). In its distribution areal, cornelian cherry production is increasing because of its high use value where mostly genotypes from natural populations are used. Most individuals occurred spontaneously as a result of open pollination, which varies widely in terms of productivity and fruit characteristics. Long-term human selection has also given rise to a great diversity of trees. In order to overcome this problem, first, it is necessary to make a selection and multiplication of selected genotypes from the native population and then to improve its properties through the different breeding programs.
Inclusion of cornelian cherry in commercial growing may be beneficial for many reasons: diversification of agriculture in the region, development of new typical agricultural products and their increased offer at local markets, introduction into food-processing industry, acquisition of new source of income for local growers and family farms, and preservation of agro-biodiversity. But in the commercial fruit production, standardized cultivars are necessary. From these reasons, selection of cornelian cherry genotypes from natural populations is an important part of breeding programs in many countries such as Turkey (Pirlak et al. 2003; Yilmaz et al. 2009), Ukraine (Klimenko 2004), Slovakia (Brindza et al. 2009), Czech Republic (Rop et al. 2010) and, recently, in Serbia (Ninić Todorović et al. 2005; Mratinić et al. 2008; Ognjanov et al. 2009; Bijelic et al. 2011).
Serbia is a well known by its rich gene pool of cornelian cherry genotypes adapted to different local conditions in different regions of the country. The largest number of natural populations can be found on mountains Stara Planina, Kopaonik, Zlatibor, Zlatar and Fruška Gora, where it grows mainly as a companion tree in hornbeam and oak forests (Bijelic et al. 2011). Moreover, it is also found on hills, slopes, in forest clearings and in thickets. On the contrary, some trees grow on farm yards or can often be seen growing on the grasslands and nearby paths as solitary trees.
The gene pool of cornelian cherry is threatened by genetic erosion. During the last decades, the native and semi-wild cornelian cherry plants have been disappearing rapidly. To determine the existing biodiversity of cornelian cherry trees, collecting and conservation will be necessary. According to Zamani et al. (2013), traditional methods for genotype characterization and identification are based on phenotypic observations. Morphological traits are useful for preliminary evaluation because they facilitate fast and simple evaluation and can be used as a general approach for assessing genetic diversity among morphologically distinguishable accessions. Since the information generated from morphological characterization is derived from a large data set consisting of qualitative and quantitative traits, the use of multivariate analysis is particularly well adapted to this type of situation (Iezzoni and Pritts 1991). Among the multivariate techniques, principal component analysis and cluster analysis are the most important strategies for the characterization, evaluation and classification of germplasm collections (Odong et al. 2011).
The aim of the current study was to characterize and quantify the variability of morphological traits within the set of 150 wild cornelian cherry accessions from the ‘Stara Planina’ Mountain, Serbia, to identify the most useful variables for discrimination among the accessions and to detect relationships between the accessions.
Materials and methods
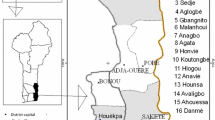
In this study, we included the set of 150 Cornus mas L. accessions selected from five geographically separated locations, with 30 accessions per each site. The investigation area was limited to the southeast Serbia, at Stara Planina Mountain. The altitude in the investigation area is rising from 513 up to 1723 m (Table 1). All accessions were selected as phenotypically mature trees from a mixed deciduous forest. Within the study plots, cornelian cherry accessions were scattered as single trees or in small groups and its collection may be considered as a completely random sample. The trees were chosen as representative of the phenotypic variability observed in the field.
Accessions have been evaluated using a set of 31 traits (Table 2). The phenotypic traits were measured in three consecutive years (2007–2009). Fruiting twigs were removed from the tree and examined during tree dormancy. Flowers used in this study were randomly selected, and all measurements were done during full bloom (when 80 % of flowers were open). For fruits and stones, all observations were made on ripe fruit and sampled randomly from the periphery of the trees.
The sphericity (φ) and surface area (S) of the fruit or stone were calculated using the following equations (Mohsenin 1978).
where L is the length; W is the width; and T is the thickness of fruit or stone.
The tree vigor was determined by five-score scale (very weak, weak, medium, strong and very strong) and tree habit by a four-score scale (upright, semi-upright, spreading and drooping) both adapted to UPOV descriptor for sour and duke cherry (2007).
Data analysis
Statistical analyses were performed for the mean values of each trait, calculated on the basis of 3-year study. First, descriptive analysis was performed to characterize the variation existing on the five locations and between all 150 cornelian cherry accessions. For qualitative characteristics, variation was assessed by distribution of frequency and expressed in percent. For quantitative traits, following parameters were evaluated: mean, minimum and maximum value, maximum/minimum value ratio and coefficient of variation (CV %).
The collected data were also analyzed by multivariate analysis of variance using principal component analysis (PCA) and clustering analysis (CA). PCA was used to study patterns of variation in a set of interrelated traits through the identification of subsets of these traits, called factors, which are substantially correlated with each other, simultaneously affecting these traits to a large extent. As a criterion to extract the main principal components, eigenvalue greater than 1 was taken, and to determine which of the PC scores accounted for the greatest amount of variation, the eigenvalues of these components were compared for each trait. Also, results of PCA were used to construct two-dimensional scatter plots for graphical overview of the relationships among accessions. CA was applied to classify the accessions into homogenous groups (Crossa and Franco 2004). The UPGA method as agglomeration rule and the Euclidean distance as a measure of dissimilarity were carried out. Statistical analyses were performed using Statistica for Windows, version 5.0 (StatSoft Inc., Tulsa, OK) statistical package.
Results
Morphological description
The first group of characteristics that was used to distinguish Cornus mas accessions consisted of five tree traits which variability is shown in Fig. 1. Different number of categories and different accessions frequencies within selected categories were observed in five populations. Considering all 150 accessions, arbor was stronger (56 %) than bush (44 %), while most of the accessions herein had medium vigor. Here as well, the main part (40 %) of the evaluated Cornus mas accessions showed spreading canopy and slightly less (35 %) semi-upright. In relation to bark color, ‘brown’, ‘reddish’ and ‘greenish’ (33, 33 and 27 %, respectively) showed a frequent tendency, sometimes tending more toward ‘gray’ or ‘blackish’ (4 and 3 %, respectively). The majority of accessions (43 %) had very low yields, while only 9 % had very high yields.

Frequency distribution for five tree traits in five locations (I-Zlot; II-Radičevac; III-Tresibaba; IV-Tupižnica; V-Babin Zub) and all 150 cornelian cherry accessions selected from natural populations in the Stara Planina Mountain, Serbia
The descriptive statistical analysis values for each of 26 quantitative traits are reported in Table 3. The highest mean value for vegetative bud length was determined in Zlot (7.22 mm) and for vegetative bud width in Tupižnica (2.72 mm). The greatest variation among populations was observed regarding the average length of fruiting twigs (LFT) and the number of flower buds per fruiting twig (NFB) as one of the most important yield components. So, LFT varied from 16.1 cm (Zlot) to 52.1 cm (Babin Zub), while NFB ranged from 7.0 (Zlot) to 14.8 (Tresibaba). Regarding locations, differences related to the average number of flowers per inflorescence (from 15.1 to 19.5) and for peduncle length (5.1–6.1 mm) were detected. In the majority of the locations, the mean value for number of pistils per flower was 1.0. The exceptions are the accessions from Babin Zub, where the number of pistils varied from 0.8 to 2.5. Mean value for flower diameter ranged from 0.89 mm (Zlot) to 2.97 mm (Tupižnica). In studied agro-ecological zone, accessions were averagely harvested between 23 August and 18 October. As regards to fruit and stone dimension, the lowest mean values were observed in Tresibaba while the highest in the Tupižnica. The length/width fruit ratio and length/width stone ratio showed similar mean values in all populations. Fruit weight had a mean value of 1.58 g, with a variation among accession’s location that ranged from 1.25 (Tresibaba) to 1.91 g (Tupižnica). Soluble solid content, as one of the most important quality parameters for many fruits, ranged from 17.2 % (Babin Zub) to 24.7 % (Zlot).
In particular, the highest maximum/minimum value ratio variation was observed in NFB, ranging from 6.9 in Babin Zub to 27.3 in Tupižnica. In contrast, the lowest maximum/minimum ratio was displayed by NPF in Zlot, Radičevac, Tresibaba and Babin Zub (0.7, 1.0, 1.0 and 2.5, respectively) and SW and ST (5.4 each) in Tupižnica. The highest coefficient of variation (CV %) was established for NFB varied from 48.0 % (Babin Zub) to 72.6 % (Tupižnica), with the lowest CV being evident in traits involving flowers and stones. None of these characters reached CV values of higher than 20 %.
Principal component analysis
Principal component analysis produced nine principal components with eigenvalues greater than 1, explaining 74.7 % of the total variability observed. A plot of the variance extracted by the first nine PC shows that the variance explained by a single PC decreased strongly between PC1 and PC6, but from PC6 onward, the decrease became very slow and several PCs with almost the same variance could then be found (Fig. 2). Also, on the basis of correlation between the original variables and PCs and using an absolute value greater than 0.50 as a criterion, it was found that these values are present only in the first five PCs. From these reasons, only the first five PCs were taken into further consideration. Traits with higher scores on PC1 are related to fruit and stone dimension, fruit and stone surface as well as fruit weight indicates 22.9 % of variance (Table 4). In this regard, this component could be called Fruit and stone size. The second factor (PC2) correlated strongly with stone sphericity and length/width stone ratio (r = 0.797 and −0.892, respectively) and moderately with tree vigor (r = 0.505) and contributed 11.7 % of total variance. Therefore, PC2 represents the second most important factor, called Stone shape and tree vigor. PC3 accounted 10.1 % variability were mainly associated with fruit sphericity and length/width fruit ratio expresses another important factor, called Fruit shape. PC4 with 7.7 % of total variance included can be consisted as a factor Flower size because this component was moderate positively correlated with petal length (r = 0.520) and was moderate negatively correlated with flower diameter (r = −0.529). Finally, PC5 indicating 6.3 % of variance had high loading for vegetative bud length and width as well as length of fruiting twigs and could be called Size of vegetative bud and fruiting twigs.

Percent of variance explained by the first nine principal components (PCs)
Scatter plot
Figure 3 shows the projection of cornelian cherry accessions on the PC1/PC2 plane, based on the regression factor score values. In general, starting from negative to positive values of PC1, the cornelian cherry accessions indicated increase in the fruit and stone size. Starting from the negative toward the positive values of PC2, the accessions were characterized by flatter stone shape and larger tree vigor. The majority of accessions are located in the central part of the scatter plot in the inner part of the ellipse (Fig. 3). This group mixed almost all accessions from locality Babin Zub, but also a significant number of accessions originating from the remaining four sites. The majority of the remaining accessions can be seen in the outer ring of the ellipse where its grouping according to their locality was noticed. Only one gap (outside of ellipse) was formed composed of six accessions selected from the location Zlot. It suggests that a great number of phenotypically similar accessions exist regardless the location, together with numerous accessions with specific characteristics originated from each studied site.

Scatter plot of PC1/PC2 plane showing the relationships between 150 cornelian cheery accessions selected from natural populations in the Stara Planina Mountain, Serbia, on five locations (closed diamond Zlot, open diamond Radičevac, open square Tresibaba, open triangle Tupižnica, closed square-Babin Zub)
Cluster analysis
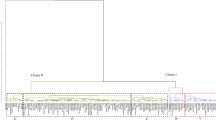
The dendrogram obtained by the cluster procedure shows numerous hierarchical levels of cornelian cherry accessions separated in three main clusters (Fig. 4). The first cluster included only one accession from location Tužnica. The second cluster consisted of the largest number of cornelian cherry accessions (108) and was split off into two distinct sub-groups, defined as cluster IA and IB. Sub-group IIA consisted of 24, while 84 accessions formed sub-group IIB. The most common location in sub-cluster IIA was Tupižnica, with nine representatives. In sub-cluster IIB, all locations were almost equally represented (around 20), except for Tresibaba location that had only ten members. Cluster III had 41 cornelian cherry accessions and was also divided into sub-groups IIIA and IIIB, with 23 and 18 accessions, respectively. Sub-cluster IIIA consisted predominantly of the accessions originated from location Tresibaba, while in IIIB, the highest number of representatives was from Radičevac location.

Dendrogram obtained from cluster analysis based on all traits studied (quantitative and qualitative) of 150 cornelian cherry accessions
Discussion
Evaluation of cornelian cherry germplasm, even in a small area points such as Stara Planina Mountain, revealed considerable phenotypic diversity among accessions for all the characters studied. Similar level of variability and similar intervals of variation for flower, fruit and stone traits were found with different cornelian cherry collections by Karadeniz (2002), Demir and Kalyoncu (2003), Pirlak et al. (2003), Klimenko (2004), Turhan et al. (2007), Prokaj et al. (2009), Ognjanov et al. (2009), Brindza et al. (2009), Bijelić et al. (2010), Mert (2012) Rad and Vand (2012). On the contrary, Bijelić et al. (2011) reported much higher values for fruit dimensions of the cornelian cherry accessions from north Serbia, while Ercisli (2004), Ercisli et al. (2006, 2011) and Hassanpour et al. (2012) established much lower soluble solid content that the values obtained in this study. This could be the result of different environmental conditions and genotype types (Güleryüz et al. 1998; Demir and Kalyoncu 2003).
Based on the CV, values in our paper can be noticed that the majority of the examined properties showed similar values in each five locations tested together with the entire population (all 150 accessions). This trend was also observed in relation to the average values, intervals of variation and maximum/minimum ratio for most traits. This phenomenon can be interpreted with a high degree of heterozygosity and therefore accessions genetic variability in each location as a result of open pollination. Also, since there is a little evidence that the different sites distinguished by the altitude show no major discrepancies between the accessions, it goes in favor that cornelian cherry can be successfully grown at high altitudes where conditions for other fruit species are unfavorable.
Principal component analysis identified that the cumulative variation explained by first five components was only 58.9 %, and according to Reim et al. (2012), this result revealed a great morphological variation, indicating a high genetic diversity between the cornelian cherry accessions, and suggests that the evaluation of different morphological characteristics remains necessary for a meaningful characterization.
The variables related to fruit and stone size and shape have high discriminating power and could be sufficient for reliable characterization of different cornelian cherry germplasm collections. The traits moderately correlated with first five PCs, representing tree vigor, flower size, vegetative bud size and fruiting twigs length may have a discriminatory effect too, but in a much smaller extent. Our results correspond to those of Yilmaz et al. (2009) and Ercisli et al. (2011), who found that the variables corresponding to fruit and stone size highly correlated with PC1 or PC2 in cornelian cherry collections studied.
Besides being indicate discriminating power of traits, according to Zeinali et al. (2009), principal components revealed intra-correlations among traits and estimate how the characters affect each other. A study of the relationships among traits could be beneficial to the breeders in their breeding programs. In this study, PC1 showed that some fruit (length, width, thickness, surface and weight) and stone traits (length, width, thickness and surface) had positive intra-correlations and can be used to predict each other. High correlation between stone sphericity and length/width stone ratio and PC2, as well as between fruit sphericity and length/width fruit ratio indicates that one of these two parameters can be used as good indicators of their shape. On the other hand, the absence of correlation between groups of traits related to the size of three, fruit, flower, twigs and buds within any of the five PCs suggests that this trait was genetically affected by independent genes not showing pleiotropic effects.
The scatter plot shows geometrical distances among accessions reflecting phenotypic and in this regard presumable genetic dissimilarity between them. The accessions had continuous variation, no apparent patterns were detected among them and it is difficult to assign accessions into main groups. Dispersed accessions disposition has been established at the level of individual locations. Opposite to the results of Aliyoun Nazari et al. (2012) who cited that the diversity of Prunus incana accessions from different geographic regions was greater than that of the accessions from a particular region, in this study, many cornelian cherry accessions originating from different locations overlap each other, which means that the same genotypes can be found in various sites. The majority of accessions are distributed in the central part of the graph. Coart et al. (2003) interpret this with the appearance of intermediate phenotypes at each location, which is the result of spontaneous hybridization between the accessions for a long time. In addition, some accessions characterized by a unique combination of properties can be also found and those are arranged outside the inner ellipse.
UPGMA clustering of the accessions into clusters and sub-clusters consistently demonstrated that the divergence between accessions originated from different altitude does not exist, showed no association between accessions from the same locality and was not in a function of an altitude. CA confirmed that the morphological variability of the cornelian cherry accessions is of a continuous nature and revealed strong overlaps between locations. Also, it was not possible to extract traits clearly responsible for cornelian cherry accessions grouping. Overall, the greatest impact on the separation of the clusters had length of fruiting twigs and number of flower buds per fruiting twig while sub-clustering was done according to some fruit and stone properties. This is in agreement with Petruccelli et al. (2013) who cited that characteristics with high degree of variations are more discriminating and can be reliable markers for the classification.
All of the results of multivariate statistical analysis in the present study indicate that observed accessions may be recognized as representative gene pool of natural population of cornelian cherry on Stara Planina Mountain and should be treated as a single population. For further collecting, it is sufficient to take accessions with different phenotypes regardless of the location of their origin.
As conclusion, natural populations of cornelian cherry from Stara Planina Mountain, Serbia, were diverse suggesting it is a high genetic potential, which could be used from the agronomic point of view to find several valuable well-adapted genotype suitable for substantial improvement of the production in our country. Furthermore, cornelian cherry genotypes could become a new source for the production of a functional food or value-added ingredients in our dietary system and enrich the range of fruit commodities on a global scale.
References
Aliyoun Nazari S, Zamani Z, Fatahi MR, Shiekh Sofla H (2012) Morphological characterization of Prunus incana Pall. by multivariate analysis. Plant Syst Evol 298:1805–1814. doi:10.1007/s00606-012-0681-x
Bijelić S, Gološin B, Ninić Todorović J, Cerovic S (2011) Morphological characteristics of best cornelian cherry (Cornus mas L.) genotypes selected in Serbia. Genet Resour Crop Evol 58:689–695. doi:10.1007/s10722-010-9612-2
Brindza P, Brindza J, Tóth D, Klimenko SV, Grigorieva O (2009) Biological and commercial characteristics of cornelian cherry (Cornus mas L.) population in the Gemer region of Slovakia. Acta Hortic 818:85–94
Coart E, Vekemans X, Smudlers MJM, Wagner I, van Huylenbroeck J, van Bockstaele E, Roldán-Ruiz I (2003) Genetic variation in the endangered wild apple (Malus sylvestris (L.) Mill.) in Belgium as revealed by amplified fragment length polymorphism and microsatellite markers. Mol Ecol 12:845–857
Crossa J, Franco J (2004) Statistical methods for classifying genotypes. Euphytica 137:19–37
Demir F, Kalyoncu IH (2003) Some nutritional, pomological and physical properties of cornelian cherry (Cornus mas L.). J Food Eng 60:335–341. doi:10.1016/S0260-8774(03)00056-6
Ercisli S (2004) Cornelian cherry germplasm resources of Turkey. J Fruit Ornam Plant Res Spec 12:87–92
Ercisli S, Orhan E, Esitken A (2006) Genetic diversity in fruit quality traits in cornelian cherry (Cornus mas L). Asian J Chem 18(1):650–654
Ercisli S, Yilmaz SO, Gadze J, Dzubur A, Hadziabulic S, Aliman J (2011) Some fruit characteristics of Cornelian Cherries (Cornus mas L.). Not Bot Hort Agrobot Cluj 39:255–259
Güleryüz M, Bolat I, Pırlak L (1998) Selection of table cornelian cherry (Cornus mas L.) types in Coruh Valley. Turkish J Agr Forest 22:357–364
Hassanpour H, Hamidoghli Y, Hajilo J, Adlipour M (2011) Antioxidant capacity and phytochemical properties of cornelian cherry (Cornus mas L.) genotypes in Iran. Sci Hortic 129:459–463. doi:10.1016/j.scienta.2011.04.017
Hassanpour H, Hamidoghli Y, Samizadeh H (2012) Some fruit characteristics of Iranian Cornelian Cherries (Cornus mas L.). Not Bot Hort Agrobot 40:247–252
Iezzoni AF, Pritts MP (1991) Application of principal component analysis to horticultural research. HortScience 26:334–338
Karadeniz T (2002) Selection of native Cornelian Cherries grown in Turkey. J Amer Pomolog Soc 56:164–167
Klimenko S (2004) The cornelian cherry (Cornus mas L.) collection, preservation and utilization of genetic resources. J Fruit Ornam Plant Res (Spec Ed) 12:93–98
Mamedov N, Craker LE (2002) Cornelian cherry: a prospective source for phytomedicine. Acta Hort 629:83–86
Mert C (2012) Studies on the structure of flowers and inflorescences of cornelian cherry (Cornus mas L.). Not Bot Horti Agrobo 40:53–57
Mohsenin NN (1978) Physical properties of plant and animal materials. Gordon and Breach Science Publisher, New York
Mratinić E, Miletić R, Fotirić M, Žikić M (2008) Biological diversity of dogwood (Cornus mas L.) population in the Stara Planina region [Serbia]. J Sci Agr Res 69:43–53
Murrell Z (1993) Phylogenetic relationships in Cornus (Cornaceae). Syst Bot 18:469–495
Ninić-Todorović J, Cerović S, Gološin B, Bijelić S, Miletić R (2005) Pomological biochemical characteristics of selected cornelian cherry genotypes. J Pomol 39:395–399
Odong TI, van Heerwaarden J, Jansen J, van Hintum TJL, van Ecuwijk FA (2011) Determination of genetic structure of germplasm collections: are traditional hierarchical clustering methods appropriate for molecular marker data? Theor Appl Genet 123:195–205. doi:10.1007/s00122-011-1576-x
Ognjanov V, Cerović S, Todorović J, Jaćimović V, Gološin B, Bijelić S, Vračević B (2009) Selection and utilization of table cornelian cherry (Cornus mas L.). Acta Hort 814:121–124
Petruccelli R, Ganino T, Ciaccheric L, Masellid F, Mariottia P (2013) Phenotypic diversity of traditional cherry accessions present in the Tuscan region. Sci Hortic 150:334–347. doi:10.1016/j.scienta.2012.11.034
Pirlak L, Guleryuz M, Bolat I (2003) Promising Cornelian Cherries (Cornus mas L.) from the Northeastern Anatolia region of Turkey. J Am Pomol Soc 57:14–18
Prokaj E, Medve A, Koczka N, Ombodi A, Dimeny J (2009) Examination of cornel (Cornus mas L.) fruits in Brsod-Abaujzemplen county (Hungary). Agronom Vēstis 12:87–93
Rad ZS, Vand SH (2012) Identify the main characteristics of some cornelian cherry germplasms. Indian J Innovations Dev 1:400–405
Reim S, Proft A, Heinz S, Hofer M (2012) Diversity of the European indigenous wild apple Malus sylvestris (L.) Mill. in the East Ore Mountains (Osterzgebirge), Germany: I. Morphological characterization. Genet Resour Crop Evol 59:1101–1114. doi:10.1007/s10722-011-9746-x
Rop O, Mlcek J, Kramarova D, Jurikova T (2010) Selected cultivars of cornelian cherry (Cornus mas L.) as a new food source for human nutrition. Afr J Biotechnol 9:1205–1210
Seeram N, Schutzki R, Chandra R, Nair MG (2002) Characterization, quantification, and bioactivities of anthocyanins in Cornus species. J Agr Food Chem 50:2519–2523. doi:10.1021/jf0115903
Turhan K, Ilkay T, Yunus P (2007) Drying characteristics and heat energy require of cornelian cherry fruits (Cornus mas L.). J Food Eng 78:735–739. doi:10.1016/j.jfoodeng.2005.09.035
UPOV. 2007. Guidelines for the Conduct of Tests for Distinctness, Uniformity and Stability. Sour cherry (Prunus cerasus L.) and duke cherry (Prunus × gondouinii (Poit. & Turpin) Rehder). TG/230/1 Corr., Geneva, Swiss, pp 1–35
Yilmaz KU, Zengin Y, Ercisli S, Orhan E, Yalcinkaya E, Taner O, Erdogan A (2009) Biodiversity, ex-situ conservation and characterization of cornelian cherry (Cornus mas l.) genotypes in Turkey. Biotechnol Biotechnol Eq 23:1143–1149
Zamani Z, Adabi M, Khadivi-Khub A (2013) Comparative analysis of genetic structure and variability in wild and cultivated pomegranates as revealed by morphological variables and molecular markers. Plant Syst Evol. doi:10.1007/s00606-013-0851-5
Zeinali H, Tabaei-Aghdaei SR, Arzani A (2009) A study of morphological variations and their relationship with flower yield and yield components in Rosa damascene. J Agric Sci Technol 11:439–448
Acknowledgments
This study was supported by the FP7 Project AREA 316004 and Serbian Ministry of Education, Science and Technology Development Project (TR 31063).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Mratinić, E., Akšić, M.F., Rakonjac, V. et al. Morphological diversity of cornelian cherry (Cornus mas L.) populations in the Stara Planina Mountain, Serbia. Plant Syst Evol 301, 365–374 (2015). https://doi.org/10.1007/s00606-014-1079-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00606-014-1079-8