
Abstract
In this research, morphological characterization of wild Prunus scoparia species, naturally growing in Iran, was investigated. Twenty-one variables along flowering and ripening date were recorded in 150 accessions of this species to detect their phenotypic variation. All studied characteristics were showing a high degree of variability, but it was highly pronounced for secondary shoot number, leaf area and shape, growth habit, fruit exocarp color, nut shape, pubescence on fruit, canopy size and trunk diameter. The majority of significant correlation coefficients were found in the characteristics representing tree and nut sizes with leaf size. Principal component analysis was performed for phenotypical diversity determination and grouping of species. Characters with high discriminating values were those related to tree height, growth habit, canopy size and kernel weight. Based on the UPGMA cluster analysis, accessions were placed in two main clusters. The first main cluster was divided into five subclusters with high variability within and between populations, while the second cluster contained 11 accessions of Estahban region. Current findings supported these opportunities since the phenotypic variability in Iranian wild P. scoparia species has been found to be very high, suggesting an extensive genetic diversity available to almond cultivar and rootstock development programs. The wide adaptation of this species indicates its potential as resources for resistance to abiotic and biotic stresses such as drought and spring frost. The conservation of the highly diverse native populations of Iranian wild P. scoparia species is recommended.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Iran represents a significant resource of germplasm of wild and cultivated fruit species, especially for those from the genus Prunus (Mozafarian 1996). Prunus L. (Rosaceae) is a morphologically diverse genus of about 200 species of small trees or shrubs with pentamerous flowers and monocarpellate drupes (Robertson 1974). Many species of this genus are economically important, especially those that are used for ornamentals and food (plums/prunes, peaches, apricots, cherries, and almonds). The most widely accepted classification of Prunus is that of Rehder (1940). He divided Prunus into five subgenera (Prunophora, Amygdalus, Cerasus, Padus, and Laurocerasus).
Subgenus Amygdalus is a small group within Prunus, containing approximately 24 species. Out of them, Almonds [P. dulcis (Mill.) D. A. Webb and relatives] and peaches [P. persica (L.) Batsch and relatives] are among the most economically important fruit crops in subtropical and temperate regions (Watkins 1976). The majority of species in subg. Amygdalus are distributed in Iran and eastern Turkey with a few in southeastern Europe, the Mediterranean region, and east into Mongolia and western China, a distribution pattern referred to as the Irano-Turanian distribution (Browicz 1969). Members of subg. Amygdalus are deciduous shrubs or small trees that are important components of northern temperate forests in Asia and Europe, mostly occurring in mountainous areas, often between 1,000 and 2,500 m in elevation. They occur in a variety of environments, but mainly in relatively dry habitats, and sometimes in deserts [e.g., P. arabica (Oliv.) Meikle]. A few species, such as P. persica, occur in more humid regions. Subgenus Amygdalus receives a great deal of attention because of its agricultural value, especially from plant breeders and horticulturists, usually to assess the potential for their use in cultivar improvement (Socias i Company 1992; Gradziel et al. 2001). Some wild species of subg. Amygdalus include Prunus spartioides, Prunus spinosissima, Prunus scoparia, Prunus elaeagnifolia, Prunus eriocalda and Prunus horrida. Among them, Prunus scoparia is distributed in different natural regions of Iran and is called Jargeh locally. It is wild deciduous shrub or small tree, usually living on dry calcareous soils at rocky mountain and has been used as a grafting rootstock for domesticated almonds to provide drought resistance (Rechinger 1969; Mozafarian 1996). Also, it is well adapted to diverse conditions of its growing region, providing an extensive germplasm resource for domestication and improvement and can have the potential of being a dwarfing rootstock for almond, and also improving other Prunus species as a gene donor (Nikoumanesh et al. 2011).
The importance of genetic diversity in breeding is essential. Therefore, the recognition and measurement of such diversity and its nature and magnitude are crucial to a breeding program. The availability and informative value of plant germplasm are becoming more and more important for the future preservation and sustainable use of genetic resources (Khadivi-Khub et al. 2012). Also, investigations of germplasm should include morphological and agronomical characterization of traits having breeding value. Characterization of plant genetic resources (PGR) usually contains a wide range of data, on both qualitative and quantitative traits. Such data are generally large and multivariate with a considerable number of descriptors measured on each of many accessions. The most widely used multidimensional analysis methods in PGR characterization are principal component analysis (PCA) and cluster analysis. The advantages of PCA in horticultural studies have been widely discussed (e.g., Iezzoni and Pritts 1991). PCA allows to evaluate multicollinear data and to determine the traits most suitable for classification. Also, cluster analysis allows to analyze simultaneously both quantitative and qualitative traits, and each entry is treated as an individual entity of equal weight (Peeters and Martinelli 1989; Iezzoni and Pritts 1991).
Evaluation and characterization as well as estimation of morphological diversity have been performed for various subg. Amygdalus collections (e.g., Sorkheh et al. 2009; Nikoumanesh et al. 2011; Zeinalabedini et al. 2012). However, despite of many reports about Prunus species, we still do not have comprehensive information about the phenotypic and genetic diversity of wild P. scoparia species. Thus, the objective of this work was to characterize phenotypic diversity of wild P. scoparia genetic resources from different natural regions of Iran using multivariate statistical analysis and to evaluate their potential in rootstock breeding. The best accessions for breeding and PGR maintenance were selected.
Materials and methods
Plant material
A total of 150 wild accessions of P. scoparia were studied from four natural regions of Iran. These regions are important producers of almond fruit in Iran and growers use P. scoparia as rootstock for cultivated almonds because of having characters such as vigor and drought tolerance. Thus, regions were selected based on potential diversity and use of this species as rootstock for almond. Ecological and geographical data of the localities are shown in Table 1. The distance between sampled accessions was at least 300 m. The sampled stands were chosen to provide maximum representation of the ecological conditions of the area. The number of accessions sampled per region ranged from 20 to 50, depending on habitat diversity.
Characterization and evaluation
Survey was made using 21 variables of tree, leaf and fruit traits along flowering and ripening date based on almond descriptors developed by the International Plant Genetic Resources Institute (IPGRI) (Gulcan 1985) for two successive years. Some of morphological characters were measured at the sampling location, and then the samples were transported to the laboratory for further study evaluation. Measurements were scored for 30 fruits and 30 leaves per accession. Quantitative traits were measured by suitable equipment such as digital caliper and precision weighing balance, while qualitative characters were considered based on rating, scoring and coding according to almond descriptor (Gulcan 1985). Pictures from tree, flower, fruit, nut and kernel of wild P. scoparia species are shown in Fig. 1.

Pictures from different organs of wild Prunus scoparia species
Statistical analysis
Analysis of variance was performed for morphological variables using SAS software (SAS Institute Inc., Cary, NC, USA). Coefficients of variation (CV %) were determined as indicators of variability. The simple correlation coefficient was calculated to determine the relationships between the studied morphological variables using the Pearson correlation coefficient by SPSS® version 17 (SPSS, Chicago, IL, USA). Relationships among the accessions were investigated by principal component analysis (PCA). PCA was performed using SPSS statistics software. Mean values were used to create a correlation matrix from which standardized principal component (PC) scores were extracted. The morphological similarity coefficients according to Euclidian method were calculated using the SIMINT program of the numerical taxonomy multivariate analysis system NTSYS-pc version 2.10 (Rohlf 2000) and the dendrograms were constructed through SAHN clustering program using the unweighted pair group method with arithmetic means (UPGMA). Also, scatter plot was created according to the PC1 and PC2 using PAST statistics software (Hammer et al. 2001).
Results and discussion
Morphological characteristics of accessions
The analysis of variance showed that most parameters were significant (P ≤ 0.01) between accessions. Mean, minimum, maximum and coefficient of variation (CV) values for studied characters in studied germplasm are given in Table 2. Among the variables studied, the largest variability corresponded to nut shape, secondary shoot number, leaf shape, growth habit, fruit exocarp color, pubescence on fruit, canopy size and trunk diameter. Coefficient of variation was the lowest for kernel taste (CV = 7.74 %), while it was the highest for fruit shape (CV = 89.95 %). Nikoumanesh et al. (2011) reported that leaf area had the highest coefficient of variation in several wild species of Amygdalus subgen. In this study, coefficient of variation for trunk diameter was 42.36 % and near to result of Nikoumanesh et al. (2011). Also, Zeinalabedini et al. (2012) reported range of 5.85–76.06 % for coefficient of variation for morphological traits in almond germplasm. Our studied germplasm has shown good candidates for clones according to our objectives: extension of the blooming season, moderate vigor and high resistance to drought. These important characteristics showed significant diversity among accessions.
The tree size and growth habit are affected by genetic and ecological factors. They are two of the main adaptation indicators in selection of rootstock for fruit crops (Browicz and Zohary 1996). In the studied P. scoparia accessions, very diverse growth habits were observed, from recumbent, spreading, erect and upright, but the dominant habit of the trees was spreading. Predominance of accessions with intermediate vigor was observed. This could be explained by the fact that the size of the plant was probably one of the first parameters to be considered as a goal of the rootstock selection by growers and breeders who attempted to achieve balance between vegetative and reproductive growth. The highest values for the tree height (3.68 m) were determined in the Estahban49 accession and the lowest (0.69 m) in the Sardasht36 that it had recumbent growth habit. The lowest trunk diameters were observed in Sardasht4, and the highest in Estahban35. Also, canopy size varied from 23 to 256 cm.
Leaf length encompassed an extensive range between the largest accession (Sardasht6, 55.20 mm) and the smallest one (Arak2, 15.10 mm), while, leaf width varied from 1.60 (in Arak2) to 9.20 mm (in Estahban2). Scored leaf traits included eight leaf shapes (lanceolate, oblanceolate, oblong-lanceolate, ovate-lanceolate, oblong, oval, elliptical and obovate), two leaf margin shapes (entire and crenate), four leaf apex shapes (acute, obtuse, acuteobtuse, notched) and presence or absence of spines. Leaf trait differences between accessions were found to be associated with environment since populations located in drier area (Estahban) had mean smallest leaf sizes than those located in semi-humid or humid regions (Arak, Mehalt and Sardasht). Accessions from the locations that receive greater annual rainfall always had greater leaf dimensions, so that accessions from Arak and Sardasht, which receives the highest annual rainfall (320.20 mm/year), had greater average leaf length and leaf width, while Estahban with the lowest annual rainfall (186.81 mm/year), had accessions with the lowest leaf length and leaf width. Totally, P. scoparia had low leaf area which can indicate better adaptation to drought conditions. This result agreed with findings of Sorkheh et al. (2009) and Khadivi-Khub et al. (2012) in wild species of Amygdalus and Cerasus subgen., respectively, who reported that decreased leaf area is an early adaptive response to water deficit and drought stress (Taiz and Zeiger 2002). Also, leaves of P. scoparia drop in natural conditions after few weeks, while the green shoots continue photosynthesis, possibly being a mechanism for tolerance against drought stress that agreed with findings of Sorkheh et al. (2009).
Nut weight varied widely between 1.50 and 2.50 g, while kernel weight ranged between 0.30 and 0.88 g. Nut length varied from 12.70 (in Arak7) to 25.50 mm (in Sardasht33), while nut width ranged between 7.90 (Mehalat6) and 20.90 mm (Arak30). Shell thickness ranged between 0.20 and 0.87 mm. The small kernel size common in this species is undesirable in breeding programs, where high kernel weight (approximately 1.00 g) is desired (Gradziel and Kester 1998; Ledbetter and Shonnard 1992), although for local production, some of the accessions showed relatively high kernel weight that resembled or even exceeded kernel weights of local commercial cultivars (Etemadi and Asadi 1999; Moradi 2005).
Unlike leaf traits, fruit dimensions did not appear associated with either rainfall or elevation, suggesting the high degree of homoplasy described in Prunus by Bortiri et al. (2006). Homoplasy occurs when characters are similar but are not derived from a common ancestor. In addition, most accessions predominantly have slightly bitter kernel because of high levels of the glucoside amygdaline, which hydrolizes to benzaldehyde and cyanide when exposed to the enzyme emulsin (Conn 1980). This trait has adaptive value by discouraging seed predation by birds and mammals including human. Individuals producing sweet kernels probably appear to have originated as mutations with subsequent seedling segregation within various Prunus species, including almond, peach and apricot (Bailey and Hough 1975).
In the studied accessions, very diverse nut shapes were observed, so that scored nut shape included 13 shapes (such as oval, flat, globular, longish and their combination), but the dominant shape of nut was oval. Also, fruit exocarp in different accessions showed ten colors from light green to reddish. Furthermore, some accessions had no or low pubescence on fruit exocarp surfaces, while others had medium or high pubescence on fruit exocarp surfaces.
Results showed that only one of the studied accessions (Sardasht35) had 100 % susceptibility to doubled fruit phenomenon, while remaining accessions had no any doubled fruit. The occurrence of double pistils is markedly increased when the trees are exposed to high temperatures throughout the period of flower differentiation (Beppu et al. 2001), but because of the same environmental conditions for Sardasht accessions, the occurrence of this phenomenon in one accession can be more related to genetic factors (Beppu et al. 2001).
Agronomic traits evaluated included flowering date (evaluated from January to March as relatively early, middle, late and very late within this period) and ripening date (evaluated from July to October as early, middle, late and very late within this period). Flowering and ripening dates were variable (from early to very late), indicating differences in the chilling requirements for flowering and differences in the growth cycle resultant (Alburquerque et al. 2008). Most accessions of Estahban regions were the earliest to bloom that indicated they had the lowest chilling requirements. On the contrary, the latest accessions to bloom were most of Arak and Sardasht accessions, indicating more chilling requirements for them. Blooming of accessions of Mehalat was middle to late. Spring frosts can be a limiting factor for almond production (Gradziel et al. 2001), so one plant breeding approach to minimize the potential of spring freeze injury to almond flowers is to develop late-blooming cultivars. Therefore, these late-blooming accessions can be used as donor parents in breeding program to develop late-blooming almond cultivars (Gradziel et al. 2001). Also, there are large variations in ripening date between studied accessions and mentioned ranges were true for ripening date.
Correlation among traits
Simple correlation coefficient analysis showed the existence of significant positive and negative correlations among characteristics (Table 3). It was found that growth habit positively correlated with tree height, trunk diameter and negatively correlated with secondary shoot number, in accordance with result of Nikoumanesh et al. (2011). There was a high positive correlation between canopy size and trunk diameter, agreeing with the findings of others (Talhouk et al. 2000; Nikoumanesh et al. 2011). Significant correlations were found between leaf variables so that, leaf length, width and shape were positively correlated with each other and leaf length was positively correlated with canopy size. These results agreed with finding of wild species of Amygdalus subgen. (Sorkheh et al. 2009; Nikoumanesh et al. 2011). The existence of close positive correlations among leaf length, leaf width, and leaf area and also positive correlation between leaf length and canopy size indicate that more leaf expansion leads to stronger aerial growth. This correlation could be considered as a suitable relationship for improving vigorous rootstocks suitable for dry environments where a fast and strong growth is needed in the beginning of the seasonal life cycle to induce and maintain appropriate vigor in scion and also for reaching to an appropriate size for budding and/or grafting as soon as possible in nurseries (Nikoumanesh et al. 2011). Also, tree height, canopy size and trunk diameter had negative correlations with leaf width. Negative correlation between leaf width and traits related to vigor is an important relationship for improving drought tolerant rootstocks in a different point of view. As in water deficiency conditions plants reduce leaf expansion and aerial growth by some strategies like reducing leaf area, leaf abscission and rolling of leaves (Scott 2008; Hopkins and Huner 2009), having thinner leaves (high leaf length and low leaf width) accompanied by less aerial growth, may lead to more root penetration. This could be considered as a possible potential for drought tolerance, particularly for the purpose of establishing orchards on slopes and also under non-irrigated conditions where a good and quick root penetration is necessary. Furthermore, leaves of P. scoparia drop in natural ecosystems after a few weeks of growth and the green shoots continue photosynthesis (as a mechanism for drought tolerance). In a similar study, Lansari et al. (1994) concluded that Moroccan almond clones have smaller leaves than European and American cultivars (a probable property in Moroccan almond germplasm to flourish in dry conditions).
In the present study, nut length, nut width and kernel weight were correlated positively with leaf width and length, indicating the role of leaf in increasing nut and kernel size (Roper and Loescher 1987). A positive correlation between nut and kernel sizes with leaf traits was determined by Sorkheh et al. (2009) and Zeinalabedini et al. (2012) in subgen. Amygdalus. In addition, nut width was highly correlated with nut length and also kernel weight was significantly correlated with nut dimensions, in accordance with results of others (Sorkheh et al. 2009; Zeinalabedini et al. 2012). A close relationship between traits could facilitate or hinder gene introgression since strong selection for a desirable trait, could favor the presence of another desirable trait from this population (Dicenta and Garcia 1992).
Principal component analysis
The aim of principal component analysis is to determine the number of main factors for reducing the number of effective parameters to discriminate genotypes. In addition, associations between traits emphasized by this method may correspond to genetic linkage between loci controlling traits or a pleiotropic effect (Iezzoni and Pritts 1991). Principal component analysis (PCA), one of the multivariate statistical procedures, was used to identify the most significant traits in the data set. Previously, PCA had been used to establish genetic relationships among cultivars and genotypes, to study correlations among tree traits and to evaluate germplasm of different Prunus species (Ruiz and Egea 2008; Horvath et al. 2008; Sorkheh et al. 2009; Nikolic et al. 2010; Rakonjac et al. 2010; Khadivi-Khub et al. 2012, 2013).
For each factor, a principal component loading of more than 0.54 was considered as being significant that indicated eight components with explaining 68.03 % of the total variance (Table 4). Principal component analysis revealed that tree and leaf traits were prevalent in the first three components and contributed to most of total variation, indicated that these attributes have the highest variation between the studied accessions and had the greatest impact on separation of the genotypes (Iezzoni and Pritts 1991). The first three components explained 34.16 % of the total variation. The first component contributed 14.22 % of the variation where tree height, growth habit, canopy size and kernel weight had the highest loadings. The second component accounted for 10.78 % of the total variation and featured secondary shoot number, main trunk presence, trunk diameter, while the third component accounted for 9.17 % of the variation featuring leaf shape, leaf length and leaf width, are thus useful for almond germplasm characterization. These results in some cases were in agreement with the results reported by others (Talhouk et al. 2000; Zeinalabedini et al. 2008; Sorkheh et al. 2009; Nikoumanesh et al. 2011). Nikoumanesh et al. (2011) studied several wild species of Amygdalus subgen. and found that the discriminating characters for their germplasm were related to leaf and tree. Zeinalabedini et al. (2008) studied nut, kernel and leaf traits in P. scoparia, P. eleagnifolia, P. hauskenchtii, and P. lycioides and reported that these traits showed significant differences among these species and indicated the possibility of the use of them in distinguishing Prunus germplasm. Also, Lansari et al. (1994) and Sorkheh et al. (2009) used a similar analysis to morphological traits among Moroccan almond clones and wild almond species, respectively, and found that the variables contributing to kernel, leaf and tree traits are important in explaining the variation among the selections. Thus, leaf, kernel and tree traits variables were consistently presented in the main components and therefore contributed high variability.
Cluster analysis and scatter plot
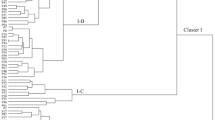
UPGMA cluster analysis allowed the assessment of similarity or dissimilarity and clarified some of the relationships among 150 accessions of P. scoparia species. Initially, the dendrogram identified two major clusters (not shown), so that the first cluster consists of most studied accessions and was divided into five subclusters with high diversity. In this cluster, some inter and intra-regional dissimilarities were observed, indicating high morphological variation between studied accessions. Accessions of Arak, Sardasht and Mehalat regions were distributed in subclusters of one to four, while subcluster five contained 12 accessions of Estahban region. Furthermore, the second cluster included 11 accessions of Estahban region which were characterized by upright or spread growth habit, higher tree height, higher fruit set, larger canopy size, higher trunk diameter and smaller leaf area. These traits were used previously for study of wild species of subg. Amygdalus (Nikoumanesh et al. 2011). The highest intra-regional similarity was observed between accessions of Estahban, while the lowest was among Sardasht accessions. Also, scatter plot was prepared according to the PC1 and PC2 that reflected relationship among accessions in terms of phenotypic resemblance and morphological characteristics. Results of scatter plot supported the results of cluster analysis and accessions were distributed in four sides of plot, so that accessions of Estahban showed high differences with others (Fig. 2). For more knowledge about inter-regional relationships, a population analysis was performed. Dendrogram of population analysis distinguished relationships between regions (Fig. 3) and a good correlation was observed between morphological variation and geographical distribution, in which high differences were revealed between Estahban population and others because of high geographical distance and different climate conditions (subtropical and arid). Also, the highest inter-regional similarity was observed between Arak and Sardasht, because of low geographical distance and similarity in climate condition. Low geographical distance can help to gene flow (Sefc et al. 2000) and the same climate condition has equal effect on morphological traits.

Scatter plot for the first two principle components (PCs) for studied Prunus scoparia accessions based on morphological characters (for explanation of accession numbers, see Table 1)

UPGMA dendrogram of population analysis for studied regions of Prunus scoparia
Results indicated a high morphological diversity of studied accessions of P. scoparia in Iran. Seed propagation, cross-incompatibility, natural hybridization, gene flow and human selection may have contributed to this variation (Sefc et al. 2000). Furthermore, this considerable variability may be mainly due to recombination (resulting from out-crossing) combined with sexual propagation for long-term and uncontrolled spread of plant material (Sefc et al. 2000). Many cultivated almonds and its related wild species such as P. scoparia express gametophytic self-incompatibility. Gametophytic incompatibility prevents self-fertilization (Socias i Company 1992), favors crosspollination (Weinbaum 1985), and maintains genetic variability within seedling populations (Arulsekar et al. 1989). This trait, although is a negative trait from the agronomic point (Socias i Company 1992), would have contributed to the phenotypic and genotypic variability that likely insured the wide distribution and adaptation of these species.
Observed phenotypic variability in our germplasm was higher than previous reports in P. scoparia based on morphological traits (Sorkheh et al. 2009; Nikoumanesh et al. 2011) and molecular markers (Zeinalabedini et al. 2008). Sorkheh et al. (2009) and Nikoumanesh et al. (2011) studied 18 and 7 accessions of P. scoparia based on morphological variables and Zeinalabedini et al. (2008) investigated genetic variation of four accessions of P. scoparia using molecular markers, while our study contained 150 accessions from different natural regions. Thus, higher morphological variation in the present study can be due to higher sampling size, different natural ecosystems and gene flow (Sefc et al. 2000). Also, this result was expected since most species of subg. Amygdalus are outbreeder and spontaneous hybridization is known to occur among species (Serafinov 1971; Denisov 1988).
Wild studied P. scoparia accessions in Iran showed high phenotypic variability and consequently could be considered as a potential resource of germplasm to be exploited in almond improvement. For instance, many accessions had very small leaves, a probable adaptation to the xerophytic conditions (<300 mm/year of rainfall). Lansari et al. (1994) found that phenotypes collected from “native” seedling populations in Morocco tended to have smaller leaves than introduced cultivars. P. scoparia has very small leaves on long, slender shoots. These leaves tend to abscise early in the season, with the green stems continuing to photosynthesize, a characteristic that is transmitted to the progeny. In this sense, Gentry (1956) reported that P. spinosissima and P. spartioides growing wild in Iran have been used by growers in very arid conditions. Development of drought-resistant almonds production systems, possibly utilizing native germplasm such as P. scoparia, would allow a more sustainable production, particularly in more marginal areas. Since the leaves of P. scoparia and its close relatives typically drop in early summer, these species may be better suited as rootstock material. Other more cultivated almond-like species such as P. arabica (Denisov 1988) or P. fenzliana (Browicz and Zohary 1996) may also contribute drought resistance without major changes in plant development patterns.
Also, frost resistance is a major breeding goal in many production areas owing to cultivated almond’s very early flowering time during late winter and early spring. The possibility of use of related species with a very late flowering date (high chilling requirements), as studied in the germplasm of P. scoparia to develop new cultivars with delayed flowering, would reduce not only frost damage, but also disease damage if flowering were delayed beyond the rainy season, and would allow more efficient use of increasingly scarce insect pollinators (Rickter 1972). The use of P. elaeagnifolia to impart late blooming has been also reported in almond breeding programs (Sorkheh et al. 2009).
Furthermore, hard shells with well-sealed sutures are reported to be more resistant to insect and fungal infestation while open sutures are highly susceptible to insect and fungal damages (Ledbetter and Shonnard 1992; Gradziel and Martinez-Gomez 2002). The use as a resource of insect and fungal resistance of related wild species with very well-sealed shell offers breeding opportunities not readily available in more traditional germplasm. In this sense, we have to note the absence of resistance to fungus as Aspergillus flavus in the cultivated almond varieties (Dicenta et al. 2003). Studied P. scoparia germpalsm can also be used as a resource of less immediate breeding needs such as more compact growth habit form or early crop maturity.
The existing gene pool in cultivated almond restricts its introduction to new regions and also limits its cultivation due to always changing environmental conditions. These problems have led to an inevitable need for genetic solutions and utilization of genetic diversity (Gradziel et al. 2001). Related almond species demonstrate a greater resistance to abiotic and biotic stresses and so represent valuable germplasm resources for breeding (Gradziel et al. 2001). Crosses between almond and related species have been successful (Felipe 1975; Socias i Company 1992; Kester and Gradziel 1996; Gradziel and Kester 1998) and numerous spontaneous interspecific hybrids have been reported (Serafinov 1971; Denisov 1988; Browicz and Zohary 1996). Interspecific hybrids between these related species including peach [P. persica (L. Batsch] × almond, and P. webbii Spach × peach have been previously used long time for almond rootstock breeding in France (Bernhard 1949), USA (Kester and Hansen 1966), Spain (Felipe 1975) or Yugoslavia (Vlasic 1977).
In conclusion, current findings supported these opportunities since the phenotypic variability in Iranian wild P. scoparia species has been found to be very high, suggesting an extensive genetic diversity available to almond cultivar and rootstock development programs. The wide adaptation of this species indicates its potential as resources for resistance to abiotic and biotic stresses. Finally, this species can be considered as characterizing the large gene pool that contributed to the domestication process of almond. Also, the conservation of the highly diverse native populations of Iranian wild P. scoparia species is recommended.
References
Alburquerque N, García-Montiel F, Carrillo A, Burgos L (2008) Chilling and heat requirements of sweet cherry cultivars and the relationship between altitude and the probability of satisfying the chill requirements. Environ Exp Bot 64:162–170
Arulsekar S, Parfitt DE, Kester DE (1989) Comparison of isozyme variability in peach and almond cultivars. J Hered 77:272–274
Bailey CH, Hough LF (1975) Apricots. In: Janick J, Moore JN (eds) Advances in fruit breeding. Purdue University Press, West Lafayette, pp 367–385
Beppu K, Ikeda T, Kataoka I (2001) Effect of high temperature exposure time during flower bud formation on the occurrence of double pistils in ‘Satohnishiki’ sweet cherry. Sci Hortic 87:77–84
Bernhard R (1949) The peach-almond and its utilization. Rev Horticole 121:97–101
Bortiri PE, Heuvel BV, Potter D (2006) Phylogenetic analysis of morphology in Prunus reveals extensive homoplasy. Plant Syst Evol 259:53–71
Browicz K (1969) Amygdalus L. In: Rechinger KH (ed) Flora Iranica, vol 66., pp 166–168
Browicz K, Zohary D (1996) The genus Amygdalus L. (Rosaceae): species relationships, distribution and evolution under domestication. Genet Resour Crop Evol 43:229–247
Conn EE (1980) Cyanogenic compounds. Annu Rev Plant Physiol 31:433–451
Denisov VP (1988) Almond genetic resources in the USSR and their use in production and breeding. Acta Hortic 224:299–306
Dicenta F, Garcia JE (1992) Phenotypical correlations among some traits in almond. J Genet Breed 46:241–246
Dicenta F, Martinez-Gomez P, Martinez-Pato E, Gradziel TM (2003) Screening for Asperguillus flavus resistance in almond. HortScience 38:266–268
Etemadi N, Asadi E (1999) Study the wild species of almond and their distribution in Chaharmahal Bakhtiari province. In: Proceedings first national conference on Almond, Shahrekord (IRAN), 24–27 August
Felipe AJ (1975) F1 hybrids of peach and almond trees as a model for both species. Agricultura 44:661–663
Gentry HS (1956) Almond culture in southern Iran. Almond Facts 21:6–7
Gradziel TM, Kester DE (1998) Breeding for self-fertility in California almond cultivars. Acta Hortic 470:109–117
Gradziel TM, Martinez-Gomez P (2002) Shell seal breakdown in almond is associated with the site of secondary ovule abortion. J Am Soc Hortic Sci 127:69–74
Gradziel TM, Martinez-Gomez P, Dicenta F, Kester DE (2001) The utilization of Prunus species for almond variety improvement. J Am Pomol Soc 55:100–108
Gulcan R (ed) (1985) Descriptor list for almond (Prunus amygdalus). International Board for Plant Genetic Resources, Rome
Hammer Ø, Harper DAT, Ryan PD (2001) PAST: paleontological statistics software package for education and data analysis. Palaeontologia Electronica 4(1):9. http://palaeoelectronica.org/2001_1/past/issue1_01.htm
Hopkins WG, Huner NPA (2009) Acclimation to environmental stress. In: Witman K, Vargas VA (eds) Introduction to plant physiology. Wiley, USA, pp 241–260
Horvath A, Christmann H, Laigret F (2008) Genetic diversity and relationships among Prunus cerasifera (cherry plum) clones. Botany 86:1311–1318
Iezzoni AF, Pritts MP (1991) Applications of principal components analysis to horticultural research. HortScience 26:334–338
Kester DE, Gradziel TM (1996) Almonds. In: Janick J, Moore JN (eds) Fruit breeding. Wiley, New York, pp 1–97
Kester DE, Hansen CJ (1966) Rootstock potentialities of F1 hybrids between peach (Prunus persica L.) and almond (Prunus amygdalus Batsch.). J Am Soc Hortic Sci 89:100–109
Khadivi-Khub A, Zamani Z, Fatahi MR (2012) Multivariate analysis of Prunus subgen. Cerasus germplasm in Iran using morphological ariables. Genet Resour Crop Evol 59:909–926
Khadivi-Khub A, Jafari HR, Zamani Z (2013) Phenotypic and genotypic variation in Iranian sour and duke cherries. Trees 27:1455–1466
Lansari A, Lezzoni AF, Kester DE (1994) Morphological variation within collections of Moroccan almond clones and Mediterranean and North American cultivars. Euphytica 78:27–41
Ledbetter CA, Shonnard CB (1992) Evaluation of selected almond (Prunus dulcis (Miller) D.A. Webb) germplasm for several shell and kernel characteristics. Fruit Var J 46:79–82
Moradi M (2005) Identification and collection of almond species and germplasm in the Chaharmahal va Bakhtiari province. In: Proceeding of IV International Symposium on Pistachios and almonds. Tehran (IRAN), 22–25 May
Mozafarian V (1996) Lexicon of Iranian plant names. Publishing contemporary vocabulary. Tehran press, Tehran, pp 234–251 (In Farsi)
Nikolic D, Rakonjac V, Milatovic D, Fotiric M (2010) Multivariate analysis of vineyard peach [Prunus persica (L.) Batsch.] germplasm collection. Euphytica 171:227–234
Nikoumanesh K, Ebadi A, Zeinalabedini M, Gogorcena Y (2011) Morphological and molecular variability in some Iranian almond genotypes and related Prunus species and their potentials for rootstock breeding. Sci Hortic 129:108–118
Peeters JP, Martinelli JA (1989) Hierarchical cluster analysis as a tool to manage variation in germplasm collections. Theor Appl Genet 78:42–48
Rakonjac V, Fotiric Aksic M, Nikolic D, Milatovic D, Colic S (2010) Morphological characterization of ‘Oblacinska’ sour cherry by multivariate analysis. Sci Hort 125:679–684
Rechinger KH (1969) Rosaceae. In: Rechinger KH (ed) Flora Iranica, vol 66., pp 187–203
Rehder A (1940) Manual of cultivated trees and shrubs hardy in North America exclusive for subtropical and warmer temperate regions, 2nd edn. Macmillan, New York
Rickter AA (1972) L’amandier. Academie Sciences Agricoles, Jardin Botanique de, Nikits
Robertson KR (1974) The genera of Rosaceae in the southeastern United States the Arnold. Arboretum 55:654–662
Rohlf FJ (2000) NTSYS-pc numerical taxonomy and multivariate analysis system. Version 2. 1. Exeter Software, Setauket
Roper TR, Loescher WH (1987) Relationship between leaf area per fruit and fruit quality in ‘Bing’ sweet cherry. HortScience 22:1273–1276
Ruiz D, Egea J (2008) Phenotypic diversity and relationships of fruit quality traits in apricot (Prunus armeniaca L.) germplasm. Euphytica 163:143–158
Scott P (2008) Plants and stress. In: Wade R (ed) Physiology and behavior of plants. Wiley, England, pp 175–191
Sefc KM, Lopez MS, Lefort F, Botta R (2000) Microsatellites variability in grapevine cultivars from different European regions and evaluation of assignment testing to assess the geographic origin of cultivars. Theor Appl Genet 100:498–505
Serafinov S (1971) A spontaneous hybrid of Amygdalus kuramica Korsh. and A. spinosissima Bge. from Afghanistan. Compt Rend Acad Sci Agric Bulgaricae 4:349–351
Socias i Company R (1992) Breeding self-fertile almonds. Plant Breeding Rev 8:313–338
Sorkheh K, Shiran B, Rouhi V, Asadi E, Jahanbazi H, Moradi H, Gradziel TM, Martinez-Gómez P (2009) Phenotypic diversity within native Iranian almond (Prunus spp.) species and their breeding potential. Genet Resour Crop Evol 56:947–961
Taiz L, Zeiger E (2002) Plant physiology, 3rd edn. Sinauer Associates, Sunderland
Talhouk SN, Lubani RT, Baalbaki R, Zurayk R, Alkhatib A, Parmaksizian L, Jaradat AA (2000) Phenotypic diversity and morphological characterization of Amygdalus L. species in Lebanon. Genet Resour Crop Evol 47:93–104
Vlasic A (1977) L’amygdalus webbi Spach ed I suolsi ibridi col pesco com portaninnesto del mandorlo. 3d Colloque GREMPA, Bari, pp 80–81
Watkins R (1976) Cherry, plum, peach, apricot and almond. In: Simmonds NW (ed) Evolution of crop plants. Longman, London, pp 242–247
Weinbaum SA (1985) Role of natural self-pollination in self-fruitfulness in almond. Sci Hortic 27:295–302
Zeinalabedini M, Majourhat K, Khayam-Nekoui M, Grigorian V, Torchi M, Dicenta F, Martinez-Gomez P (2008) Comparison of the use of morphological, protein and DNA markers in the genetic characterization of Iranian wild Prunus species. Sci Hortic (Amsterdam) 116:80–88
Zeinalabedini M, Sohrabi S, Nikoumanesh K, Imani A, Mardi M (2012) Phenotypic and molecular variability and genetic structure of Iranian almond cultivars. Plant Syst Evol 298:1917–1929
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Khadivi-Khub, A., Anjam, K. Morphological characterization of Prunus scoparia using multivariate analysis. Plant Syst Evol 300, 1361–1372 (2014). https://doi.org/10.1007/s00606-013-0967-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00606-013-0967-7