
Abstract
Purpose
There is much interest in the development of a cellular therapy for the repair or regeneration of degenerate intervertebral discs (IVDs) utilising autologous cells, with some trials already underway. Clusters of cells are commonly found in degenerate IVDs and are formed via cell proliferation, possibly as a repair response. We investigated whether these clusters may be more suitable as a source of cells for biological repair than the single cells in the IVD.
Methods
Discs were obtained at surgery from 95 patients and used to assess the cell viability, growth kinetics and stem or progenitor cell markers in both the single and clustered cell populations.
Results
Sixty-nine percent (±15) of cells in disc tissue were viable. The clustered cell population consistently proliferated more slowly in monolayer than single cells, although this difference was only significant at P0–1 and P3–4. Both populations exhibited progenitor or notochordal cell markers [chondroitin sulphate epitopes (3B3(−), 7D4, 4C3 and 6C3), Notch-1, cytokeratin 8 and 19] via immunohistochemical examination; stem cell markers assessed with flow cytometry (CD73, 90 and 105 positivity) were similar to those seen on bone marrow-derived mesenchymal stem cells.
Conclusions
These results confirm those of previous studies indicating that progenitor or stem cells reside in adult human intervertebral discs. However, although the cell clusters have arisen via proliferation, there appear to be no greater incidence of these progenitor cells within clusters compared to single cells. Rather, since they proliferate more slowly in vitro than the single cell population, it may be beneficial to avoid the use of clustered cells when sourcing autologous cells for regenerative therapies.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Degeneration of the intervertebral disc (IVD) in humans is an important phenomenon due to its association with common clinical conditions including spinal stenosis, back pain [1] and disc herniation [2]. Current treatment regimens for these conditions are limited, often being targeted to patients when degeneration is extensive and involving interventional surgery. There is a great deal of interest in biological treatments, as these offer the promise of a less invasive and potentially more permanent solution. Autologous chondrocyte implantation (ACI) has been used successfully for two decades for treating damaged articular cartilage, either caused by trauma or degeneration [3, 4]. A similar technique to ACI using autologous disc cells has been used to a limited extent for treating herniated discs [5], with other trials ongoing [6]. Such autologous cell therapy usually relies on sourcing cells from within the patient’s IVDs.
There is a great deal of evidence in the literature that the viability of cells within human IVDs is low with up to 80 % of cells having a necrotic or apoptotic appearance, even in young individuals [7, 8]. Despite this, a feature of degenerate IVDs is the presence of clones or clusters of cells [9, 10] which have formed from increased proliferation [11]. These resemble cell clusters found in osteoarthritic cartilage [12]. In both articular cartilage and IVD, markers which are believed to represent progenitor or stem cells have been shown to be present in tissue sections [13–15], often occurring within clusters of cells. These include epitopes representing different sulphation patterns on chondroitin sulphate (CS) chains of glycosaminoglycans [14], molecules such as OCT3/4, indicating pluripotency, and Notch and its ligand, Jagged, which are involved in organ formation and morphogenesis [16, 17]. Cluster of differentiation (CD) markers of stem cells have also been reported [15].
We hypothesised that these cell clusters may have an enhanced population of stem cells in comparison to the remaining cells within the IVD matrix. Hence, using human surgical samples such as might be used as a source of autologous cells for biological therapies, we have firstly examined the viability of the cells and the presence of ‘progenitor markers’ in vivo, before determining the growth characteristics and stem cell immunoprofile of clustered compared to single cells in vitro.
Methods
Patient samples
Following local research ethical committee approval and informed consent, surgical samples of human IVD tissue were obtained from a total of 95 patients, aged 41 ± 10 years, having routine surgery for IVD disorders (Table 1). Bone marrow aspirates or iliac crest bone chips were also received from four patients undergoing spinal fusion procedures. Sub-groups of these surgical samples were used in the studies described here. Samples were dissected macroscopically and used to assess cell viability, snap-frozen for morphological and immunohistochemical studies or digested for cell isolation and subsequent monolayer culture.
Cellular viability
Freshly excised IVD tissue [containing both nucleus pulposus (NP) and annulus fibrosus (AF) tissue, when present], obtained from 40 patients, was incubated in 25 μm chloromethyl fluorescein diacetate (CMFDA; Life Technologies, UK) and 2 µm ethidium homodimer-1 (EH; Life Technologies) in phosphate buffered saline (PBS; Life Technologies) for 2 h at 37 °C, blotted dry and snap-frozen [18]. Cryosections were collected onto poly-l-lysine-coated slides and mounted under coverslips using Vectashield containing 4′,6-diamidino-2-phenylindole (DAPI; Vector Laboratories Ltd, UK) to stain the cell nuclei for cell count verification. Cell viability was quantified by counting green CMFDA-positive viable cells (due to cleavage of the fluorophore by active esterases in live cells) and red EH-positive dead cells (the EH entering through the damaged membrane of dead or dying cells and binding to the nuclear DNA) using a Leica DMLB fluorescent microscope. At least 200 cells were counted from a minimum of five fields of view using a 40× objective. Cryosections were also stained with haematoxylin and eosin (H&E) for assessing morphology.
Stem cell markers: immunohistochemistry
Immunohistochemistry for putative markers of progenitor cells [Notch-1 (Developmental Studies Hybridoma Bank, USA) and chondroitin sulphate (CS) epitopes, 3B3(−), 4C3, 6C3 and 7D4 [13, 19], as well as notochordal cell markers (cytokeratins 8 and 19; Acris Antibodies, Herford, Germany)] [20], was performed as previously described [21] using IVD samples (NP tissue) obtained from 37 patients. Cryosections were incubated overnight at 4 °C with the primary antibody (Table 2). For negative controls, the primary antibody was replaced with isotype-matched immunoglobulins (Dako UK Ltd, UK). After incubating with a biotinylated secondary antibody, endogenous peroxidase was blocked by incubation with 0.3 % (v/v) hydrogen peroxide (VWR International, UK) in methanol for 30 min. Labelling was amplified by incubating with an avidin biotin complex (Vectastain Elite kit, Vector Laboratories, UK) for 30 min and visualised with 0.06 % (w/v) diaminobenzidine tetrachloride (DAB, Sigma-Aldrich, UK). The intensity of DAB staining in cellular, pericellular, territorial matrix and inter-territorial matrices within the tissue sections (Fig. 1e) was evaluated semi-quantitatively by scoring each region as 0–7, ranging from none to very strong staining; in addition, the percentage of positively stained cells was assessed, as before.

Intervertebral disc cells in tissue sections and following isolation. a Clustered and single cell populations in a cryosection of IVD (42 year old, L4/5 IVD with protrusion) contained both live cells (green fluorochrome) and dead cells (red fluorochrome, white arrow). Sequential straining of enzyme-digested IVD tissue resulted in the isolation of three cell fractions: single cells <40 μm in size (b), small clusters 40–70 μm in size (c) and large clusters >70 μm in size (d). The cells (C), together with the pericellular matrix (PM, as seen in b, c) constitute a chondron. e Schematic demonstrating the different regions of IVD cryosections scored for immunohistochemical staining intensity: cellular (C), pericellular matrix (PM), territorial matrix (TM) and inter-territorial matrix (ITM); f Staining intensity often varied within the cryosections, particularly around the cell clusters (cryosection of 47 year old L4/5 IVD with spondylolisthesis). Cells are stained with Mayer’s haematoxylin and scale bars 20 μm
Cell culture
Samples of NP and AF tissue obtained from six patients (and NP alone from a further four patients) were finely minced and digested for 16 h at 37 °C with 0.8 mg/ml collagenase type XI (Sigma-Aldrich, UK) in DMEM/F12 (Life Technologies) before passing sequentially through cell strainers of pore sizes 70 and 40 μm (BD Biosciences, UK), forming three fractions of <40, 40–70 and >70 μm size. This digestion protocol was established as the optimum for differentially isolating single and clustered cell populations. Those cells existing within clusters following isolation could not be dissociated either with further enzymatic digestion (including with trypsin, collagenase XI and collagenase I), or with mechanical agitation (extensive vortex mixing or rapid pipetting). This suggests that the isolated single and clustered populations, following collagenase XI digestion, used routinely in this study, reflects how they would occur in vivo. Cytospins (Shandon, UK) of the freshly isolated cell populations were also generated to monitor the cell populations, via H&E staining. Cell size was determined using Nikon NIS-Elements image analysis software. For monolayer culture, cells were seeded at 5 × 103 cells/cm3. At 80 % confluence, cells were passaged, counted [using the trypan blue (Sigma) exclusion method] and re-seeded. Population doublings (PD) were calculated as previously reported [19]. Culture was continued to passage (P) 5 and maintained as previously described [22]. Bone marrow aspirates or iliac crest bone washouts were layered over Lymphoprep (Fresenius Kabi AG, Germany) and centrifuged at 900g for 20 min. A ‘buffy coat’ layer of mononuclear cells, separated by density gradient, was harvested and seeded at 2 × 107 cells per 75 cm2 culture flask and maintained as previously described [23].
Senescence
Cultured IVD cells from eight of the patient samples were assessed for senescence associated-β-galactosidase (SA-β-Gal) at the later passages, as previously described [24]. Senescent cells were determined as those containing positively stained lysosomes at pH 6, in comparison to pH 4 where all cell lysosomes should be positive. Percentage cell positivity was calculated as before, counting at least 200 cells.
Stem cell markers: flow cytometry
Flow cytometry was performed to determine the presence of CD markers, described as part of the minimal criteria for defining mesenchymal stem cells (MSCs) by the International Society for Cellular Therapy (ISCT) [25] [i.e. positive for CD73, CD90 and CD105 (BD Biosciences) and negative for CD14, CD19, CD31, CD34, CD45 and HLA-DR (Immunotools, Germany)]. Both clustered and single cell populations cultured to varying passages between 1 and 5 were examined and compared to bone marrow-derived MSCs, as previously reported [26]. Cell populations were isolated from NP tissue obtained from five patients and bone marrow MSCs from three of these patients for this analysis. Immunoreactivity was assessed using the BD FACScan flow cytometer and analysed with Cell Quest software (BD Biosciences), where staining was expressed as the percentage of the cells stained positively in comparison to the isotype-matched immunoglobulin negative control.
Statistical analysis
Data was analysed using Microsoft Excel and Analyse-It (version 2.21; Analyse-It Software Ltd, UK) add-on software. Where data were normally distributed, differences were calculated using a Student’s t test or one-way ANOVA with Bonferroni post hoc test. Non-normally distributed data was assessed using a Mann–Whitney test or Kruskal–Wallis. Significance was taken as p < 0.05.
Results
Cell viability and patient demographics
Cell viability in IVD tissue was determined as 68.9 ± 14.6 % (range 30.3–100 %). Whilst EH-bound cell nuclei were found in both single and clustered cells throughout the tissue sections (Fig. 1a), they were most frequent at the tissue periphery. More clustered cells were viable than single cells in the excised samples (73.5 ± 24.4 vs. 65.9 ± 16.9 %, respectively; Mann–Whitney test, p = 0.001), but patient age or gender had no effect on this property or any of the following studies. Analysis of H&E stained sections showed that 50 ± 20.6 % (range 5–95 %) of cells were within clusters and there were between 3 and 30 cells in each cluster (mean 6 ± 4.5). Both clustered and single cells could often be seen to sit within a clear region of pericellular matrix or capsule, forming a chondron (Fig. 1b, c).
Stem cell markers: immunohistochemistry
Immunostaining for the putative stem cell markers within the IVD tissue varied between patient samples, in both location and intensity of staining and between epitopes investigated (Fig. 2). Staining was generally widespread, but no individual regions or populations had a consistent staining pattern. The territorial matrices of cell clusters were often fainter in intensity than the surrounding inter-territorial matrices, suggesting a difference in matrix composition. With the exception of 3B3(−) and 4C3 which showed strong staining, the inter-territorial matrices showed little to no staining across the patient samples (Fig. 3). There was no significant difference between the percentage of positively stained cells in the clustered and single cell populations (Table 3). Within cell clusters, staining for the CS epitopes was often variable and both positive and negative cells could be seen within any one cluster. However, the staining seen for the Notch-1 and CK epitopes was less varied, with cells within a particular cluster generally being entirely positive or entirely negative (Fig. 2).

Immunohistochemical staining of surgical IVD samples, for chondroitin sulphate epitopes [3B3(−), 7D4, 4C3 and 6C3], Notch-1 and cytokeratins 8 and 19. Staining patterns varied throughout the inter-territorial matrices and also in and around cell clusters and single cells (black arrows). Frequently paler intensity staining was seen around the cell clusters (e.g. 4C3). Isotype-matched negative controls are also shown. Cell nuclei are counterstained with Mayer’s haematoxylin and scale bars 50 μm

Intensity and frequency of immunohistochemical staining of the inter-territorial matrix (ITM) for all antibodies used. Staining intensity was scored from none to very strong. The IVD tissue sections stained with the CS epitope, 4C3, all showed at least moderately strong staining in the ITM, whereas the other epitopes resulted in mostly faint ITM staining
Clustered and single cell culture
When grown in monolayer culture, the cells from within the clusters migrated out and formed a confluent monolayer by the end of P0, which appeared indistinguishable from the cultures of isolated single cell populations. As both the >70 μm and the 40–70 μm fractions contained clustered cells at isolation (as seen in the cytospins; Fig. 1b–d) and there was consistently no difference seen between the growth kinetics, the results of these two groups were combined. Therefore, analyses presented here refer to the <40 μm population (single cells) and the >40 μm population. At the point of initial isolation, the cells existing within clusters had a significantly smaller diameter than the single cells (14.7 ± 3.9 vs. 19.7 ± 7.1 µm diameter, respectively; Mann–Whitney test, p < 0.001). The most notable PDs were seen at P0–1, where the clustered cells underwent significantly more PDs than the single cells (3.3 and 1.1 PDs, respectively; Mann–Whitney test, p = 0.0002; Fig. 4a). Between passages 1 and 5, PDs were fairly consistent, regardless of cell population or passage number. For each passage, clustered cells consistently took a longer time to reach 80 % confluence than single cells. This difference was significant at P0–1 and P3–4 (Mann–Whitney test, p = 0.005; Fig. 4b). Time taken to reach confluence decreased with increasing passage number of the clustered cell population (Pearson’s correlation; r = −0.89, p = 0.045), but not for single cells. Lysosomal β-galactosidase staining within the cell cytoplasm (at pH 4) was seen in 97.7 ± 3.4 % of cells. At pH 6, SA-β-Gal staining was seen in both clustered and single cell populations. At passage 4, 15.4 ± 14.9 and 19.8 ± 7.8 % of clustered and single cells stained, respectively, whilst at passage 5, the number of cells staining increased to 20.4 ± 11.2 and 23.9 ± 11.4 of clustered and single cells, respectively. Although the percentage of SA-β-Gal positive cells was greater in the single than clustered cell population, this difference was not significant.

Growth kinetics of clustered and single cell populations in monolayer culture. a Freshly isolated cell clusters underwent a significantly greater number of population doublings than the single cells (Mann–Whitney test, p = 0.005). After the first passage, however, there was no difference in the number of population doublings for single or clustered cell populations at subsequent passages. b Cells isolated from clusters consistently took longer to reach confluence than cells existing singly; at P0–1 and P3–4, this difference was significant (p = 0.02 and 0.007, respectively; n = 44 cell populations). Significant differences are indicated as *p < 0.05, **p < 0.01 and ***p < 0.001
Stem cell markers: flow cytometry
Clustered and single IVD cell populations showed similar results for the cell surface markers suggested to indicate stem cells by the ISCT [25] (Fig. 5). However, the populations did not always fulfil the ≥95 and ≤2 % positivity or negativity, respectively, as specified but were generally >80 and <10 %, respectively, apart from CD14. The IVD cells showed unexpectedly high percentages of cells that were positive for CD14 (up to 66.5 %). There was no correlation between the passage number of the IVD cells and the percentage of expression of any of the cell surface markers analysed. Like the IVD cells, cultured MSCs fulfilled the general pattern of expression, except for cells demonstrating higher CD14 than expected (up to 50.9 % of the population).

Representative flow cytometry histograms of clustered and single IVD cell populations and bone marrow MSCs using the profile to define an MSC as stated by the ISCT (i.e. positive for CD73, 90 and 105 but negative for the others shown). The dot plot represents forward scatter (FSC) vs. side scatter (SSC), whilst the purple histograms represent the isotype controls
Discussion
It has long been known that IVD cells can exist within the matrix of the degenerate human NP either singly or as groups or clusters of cells [27]. However, to our knowledge, this study is the first time that they have been isolated and studied in vitro as two separate populations. Indeed, in some reports, the clustered cell population has been discarded and therefore entirely excluded from study [28–30].
We have shown that the clustered and single cell populations were remarkably similar, both in vitro (apart from the period initially following isolation) and in vivo. The more PDs and longer time taken to reach confluence during the first passage of clustered cells, in comparison to single cells could be due to the time taken for the cells to migrate out of the clusters. Not only may the cells have to overcome contact inhibition of neighbouring cells within the cluster but also have to break down the pericellular capsule if present (Fig. 1c).
Senescent (SA-β-Gal positive) cells were seen equally in both clustered and single cell populations in vitro. This is contradictory to the previous findings of cells within sections of human IVD, i.e. in vivo [10], where the cell clusters contained significantly more senescent cells than those existing singly. This may be due to the isolation and cell culture processes in the present study resulting in the selection of a proliferative cell population with the clustered and single cell populations becoming more similar following time in monolayer culture and therefore showing no difference in SA-β-Gal positivity. A further explanation would be if the senescent disc cells within the clusters isolated from the disc tissue behave as senescent osteoblasts have been reported to, being less adherent in culture with a lower attachment frequency [31].
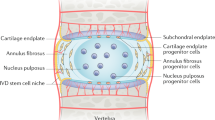
The environment of a stem cell niche acts to keep stem cells in a quiescent state until required for tissue regeneration or repair [32]. The presence of a stem cell niche in the IVD has been identified by the occurrence of a panel of markers including Stro-1, Jagged-1 and Notch-1 [33–35]. Notch-1 is transiently expressed by cells and is involved in both induction and inhibition of proliferation as well as cell fate in stem cell niches [11, 16]. It is also found in a region of known progenitor cells in articular cartilage [13]. Positive Notch-1 immunofluorescence staining in IVD cells, reported by Brisby et al. [15], in a small sub-population of the IVD cells, is in agreement with our findings in this study, suggesting a sub-population enriched in stem cells in both clustered and single cells.
Recently, a strong co-localisation of Notch-1 with the CS epitope, 7D4, has been reported in this IVD stem cell niche region [36]. 7D4, together with the other CS epitopes used in this study [3B3(−), 4C3 and 6C3], have been reported in both IVD tissue [14, 37] and developing, injured or repairing articular cartilage [38–40]. The presence of these epitopes in the degenerate IVD samples may be a repair response recapitulating developmental pathways, as is seen in repairing cartilage [39].
Our findings of positive staining for the cytokeratins, CK8 and CK19, which are commonly found in notochordal cells [41] [20, 42], within all or no cells of clusters are similar to those described by Sun et al. [43] who found all cells within clusters were uniformly either positive or negative. These authors suggest the positive clusters may be autophagosomes formed from notochordal cells. We found positive cytokeratin staining in samples from a wider age range than reported by Weiler et al. [42], who found no positive staining for CK8 and CK19 in surgical samples from patients older than 47 years. However, since microfilaments such as cytokeratins have been reported in normal chondrocytes [44], perhaps they are not unique markers for notochordal cells.
As well as immunohistochemistry, flow cytometry was used to assess the presence of stem cell markers. Due to the cell number required for flow cytometry, this study was only possible on the cultured populations of single and clustered cells rather than freshly isolated cells. The culturing process possibly resulted in an altered marker expression from that seen in vivo. Nonetheless, both clustered and single cell populations, as well as bone marrow-derived MSCs largely exhibited the pattern of expression specified by the ISCT [25]. However, both the IVD cells and the MSCs showed a greater expression of CD14 than expected. The presence of CD14, generally thought of as a monocyte or macrophage marker [45], may be indicative of an inflammatory environment due to its role as a co-receptor for lipopolysaccharide as an immune response (reviewed by [46]). The positive expression of CD14 by the cells may also be indicative of the expression of an epitope that cross-reacts with the CD14 antibody, as reported by Pilz et al. [47] who stated that MSCs express a similar epitope.
Following conflicting reports on the viability of cells within the degenerate IVD [7, 48], we have found that the majority of cells within surgical samples are viable, a promising finding when considering this tissue as a source of cells for regenerative therapies. As described in a previous study [49], we noted the presence of dead cells, particularly at the tissue periphery in these surgical samples. This is most likely due to cellular trauma inflicted during the surgical excision, which has been shown to cause cell death in articular cartilage [50].
Whilst this study is aimed at assessing the suitability of autologous IVD cell as a source for cellular therapy, there are still many obstacles that need to be overcome before a treatment of disc degeneration can be facilitated with cells (reviewed in [51]). For example, it is still not known whether the regeneration of the degenerate IVD would result in the reduction of pain, nor whether the conditions within the degenerate IVD would physically and nutritionally support a population of cells. Until these problems are addressed more fully, cellular therapy will remain a specialist treatment.
In conclusion, both the clustered and single cell populations examined in human IVD showed good viability in vivo and contain a similar proportion of cells with stem cell or progenitor cell properties. These cells may resemble those of resident stem cell niches found in other tissues, such as the satellite stem cells in adult skeletal muscle (making up 2–7 % of muscle cells), which remain quiescent in health but proliferate following injury to induce regeneration (reviewed by [52]). As clustered disc cells proliferate more slowly in vitro, show increased senescence and stress proteins in vivo [10, 53] and appear to have no greater enrichment of progenitor or stem cells over single disc cells, they appear to offer little or no advantage or benefit for biological therapies than single cells isolated from the disc.
References
Cheung KMC, Karppinen J, Chan D, Ho DWH, Song YQ, Sham P, Cheah KSE, Leong JCY, Luk KDK (2009) Prevalence and pattern of lumbar magnetic resonance imaging changes in a population study of one thousand forty-three individuals. Spine 34:934–940
Adams MA, Roughley PJ (2006) What is intervertebral disc degeneration, and what causes it? Spine 31:2151–2161
Bhosale A, Kuiper JH, Johnson WEB, Harrison P, Richardson JB (2009) Midterm to long-term longitudinal outcome of autologous chondrocyte implantation in the knee joint––a multilevel analysis. Am J Sports Med 20:1–8
Jungmann PM, Salzmann GM, Schmal H, Pestka JM, Sudkamp NP, Niemeyer P (2012) Autologous chondrocyte implantation for treatment of cartilage defects of the knee: what predicts the need for reintervention? Am J Sports Med 40:58–67
Meisel HJ, Ganey T, Hutton WC, Libera J, Minkus Y, Alasevic O (2006) Clinical experience in cell-based therapeutics: intervention and outcome. Euro Spine J 15(3):397–405
Kregar-Velikonja N, Urban J, Fröhlich M, Neidlinger-Wilke C, Kletsas D, Potocar U, Turner S, Roberts S (2013) Cell sources for nucleus pulposus regeneration. Eur Spine J. doi:10.1007/s00586-013-3106-9:
Trout JJ, Buckwalter JA, Moore KC (1982) Ultrastructure of the human intervertebral disc: II. Cells of the nucleus pulposus. Anat Rec 204:307–314
Sitte I, Kathrein A, Pfaller K, Pedross F, Roberts S (2009) Intervertebral disc cell death in the porcine and human injured cervical spine after trauma. Spine 34:131–140
Pritzker KPH (1977) Aging and degeneration in the lumbar intervertebral disc. Orthop Clin N Am 8:65–77
Roberts S, Evans EH, Kletsas D, Jaffray DC, Eisenstein SM (2006) Senescence in human intervertebral discs. Eur Spine J 15:S312–S316
Johnson WEB, Eisenstein SM, Roberts S (2001) Cell cluster formation in degenerate lumbar intervertebral disc is associated with increased disc cell proliferation. Connect Tissue Res 42:197–207
McGlashan SR, Cluett EC, Jensen CG, Poole CA (2008) Primary cilia in osteoarthritic chondrocytes: from chondrons to clusters. Dev Dyn 237:2013–2020
Hayes AJ, Tudor D, Nowell MA, Caterson B, Hughes CE (2007) Chondroitin sulfate sulfation motifs as putative biomarkers for isolation of articular cartilage progenitor cells. J Histochem Cytochem 62:125–138
Hayes AJ, Hughes CE, Ralphs JR, Caterson B (2011) Chondroitin sulphate sulphation motif expression in the ontogeny of the intervertebral disc. Eur Cells Mater 21:1–14
Brisby H, Papadimitriou N, Brantsing C, Bergh P, Lindahl A, Barreto HH (2013) The presence of local mesenchymal progenitor cells in human degenerated intervertebral discs and possibilities to influence these in vitro: a descriptive study in humans. Stem Cells Dev 22:804–814
Mead TJ, Yutzey KE (2009) Notch pathway regulation of chondrocyte differentiation and proliferation during appendicular and axial skeleton development. Proc Natl Acad Sci USA 106:14420–14425
Khan IM, Palmer EA, Archer CW (2010) Fibroblast growth factor-2 induced chondrocyte cluster formation in experimentally wounded articular cartilage is blocked by soluble Jagged-1. Osteoarthr Cartil 18:208–219
Gruber HE, Gordon B, Norton HJ, Kilburn J, Williams C, Zinchenko N, Heath J, Ingram J, Hanley EN Jr (2008) Analysis of cell death and vertebral end plate bone mineral density in the annulus of the aging sand rat. Spine J 8:475–481
Khan IM, Bishop JC, Gilbert S, Archer CW (2009) Clonal chondroprogenitors maintain telomerase activity and Sox9 expression during extended monolayer culture and retain chondrogenic potential. Osteoarthr Cartil 17:518–528
Gilson A, Dreger M, Urban JP (2010) Differential expression level of cytokeratin 8 in cells of the bovine nucleus pulposus complicates the search for specific intervertebral disc cell markers. Arthritis Res Ther 12:R24
Roberts S, Menage J (2004) Microscopic methods for the analysis of engineered tissues. In: Hollander AP, Hatton PV (eds) Methods in molecular biology, vol 238., Biopolymer methods in tissue engineeringHumana Press Inc., Totowa, pp 171–195
Johnson WEB, Sivan S, Wright KT, Eisenstein SM, Maroudas A, Roberts S (2006) Human intervertebral disc cells promote nerve growth over substrata of human intervertebral disc aggrecan. Spine 31:1187–1193
Wright KT, El Masri W, Osman A, Roberts S, Travedi J, Ashton BA, Johnson WEB (2008) The cell culture expansion of bone marrow stromal cells from humans with spinal cord injury: implications for future cell transplantation therapy. Spinal Cord 46:811–817
Dimri GP, Lee X, Basile G, Acosta M, Scott G, Roskelley C, Medrano EE, Linskens M, Rubelj I, Pereira-Smith O, Peacocke M, Campisi J (1995) A biomarker that identifies senescent human cells in culture and in aging skin in vivo. Proc Natl Acad Sci 92:9363–9367
Dominici M, Le Blanc K, Mueller I, Slaper-Cortenbach I, Marini FC, Krause DS, Deans RJ, Prockop DJ, Horwitz EM (2006) Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 8:315–317
Mennan C, Wright K, Bhattacharjee A, Balain B, Richardson J, Roberts S (2013) Isolation and characterisation of mesenchymal stem cells from different regions of the human umbilical cord. Biomed Res Int 2013:916136
Erskine L, McCaig CD (1997) Integrated interactions between chondroitin sulphate proteoglycans and weak dc electric fields regulate nerve growth cone guidance in vitro. J Cell Sci 110:1957–1966
Chelberg MK, Banks GM, Geiger DF, Oegema TR Jr (1995) Identification of heterogeneous cell populations in normal human intervertebral disc. J Anat 186(Pt 1):43–53
Kim JH, Deasy BM, Seo HY, Studer RK, Vo NV, Georgescu HI, Sowa GA, Kang JD (2009) Differentiation of intervertebral notochordal cells through live automated cell imaging system in vitro. Spine (Phila Pa 1976) 34:2486–2493
Hegewald AA, Endres M, Abbushi A, Cabraja M, Woiciechowsky C, Schmieder K, Kaps C, Thome C (2011) Adequacy of herniated disc tissue as a cell source for nucleus pulposus regeneration. J Neurosurg Spine 14:273–280
Kassem M, Ankersen L, Eriksen EF, Clark BF, Rattan SI (1997) Demonstration of cellular aging and senescence in serially passaged long-term cultures of human trabecular osteoblasts. Osteoporos Int 7:514–524
Mitsiadis TA, Barrandon O, Rochat A, Barrandon Y, De Bari C (2007) Stem cell niches in mammals. Exp Cell Res 313:3377–3385
Henriksson H, Thornemo M, Karlsson C, Hagg O, Junevik K, Lindahl A, Brisby H (2009) Identification of cell proliferation zones, progenitor cells and a potential stem cell niche in the intervertebral disc region: a study in four species. Spine (Phila Pa 1976) 34:2278–2287
Henriksson HB, Svala E, Skioldebrand E, Lindahl A, Brisby H (2012) Support of concept that migrating progenitor cells from stem cell niches contribute to normal regeneration of the adult mammal intervertebral disc: a descriptive study in the New Zealand white rabbit. Spine 37:722–732
Henriksson HB, Brisby H (2013) Development and regeneration potential of the mammalian intervertebral disc. Cells Tissues Organs 197:1–13
Shu C, Hughes C, Smith SM, Smith MM, Hayes A, Caterson B, Little CB, Melrose J (2013) The ovine newborn and human foetal intervertebral disc contain perlecan and aggrecan variably substituted with native 7D4 CS sulphation motif: spatiotemporal immunolocalisation and co-distribution with Notch-1 in the human foetal disc. Glycoconj J 30:717–725
Caterson B (2012) Fell-Muir Lecture: chondroitin sulphate glycosaminoglycans: fun for some and confusion for others. Int J Exp Pathol 93:1–10
Visco DM, Johnstone B, Hill MA, Jolly GA, Caterson B (1993) Immunohistochemical analysis of 3-B-3(−) and 7-D-4 epitope expression in canine osteoarthritis. Arthritis Rheum 36:1718–1725
Hazell PK, Dent C, Fairclough JA, Bayliss MT, Hardingham TE (1995) Changes in glycosaminoglycan epitope levels in knee joint fluid following injury. Arthritis Rheum 38:953–959
Melrose J, Isaacs MD, Smith SM, Hughes CE, Little CB, Caterson B, Hayes AJ (2012) Chondroitin sulphate and heparan sulphate sulphation motifs and their proteoglycans are involved in articular cartilage formation during human foetal knee joint development. Histochem Cell Biol 138(3):461–475
Naka T, Iwamoto Y, Shinohara N, Chuman H, Fukui M, Tsuneyoshi M (1997) Cytokeratin subtyping in chordomas and the fetal notochord: an immunohistochemical analysis of aberrant expression. Mod Pathol 10:545–551
Weiler C, Nerlich AG, Schaaf R, Bachmeier BE, Wuertz K, Boos N (2010) Immunohistochemical identification of notochordal markers in cells in the aging human lumbar intervertebral disc. Eur Spine J 19:1761–1770
Sun Z, Wang HQ, Liu ZH, Chang L, Chen YF, Zhang YZ, Zhang WL, Gao Y, Wan ZY, Che L, Liu X, Samartzis D, Luo ZJ (2013) Down-regulated CK8 expression in human intervertebral disc degeneration. Int J Med Sci 10:948–956
Ralphs JR, Benjamin M, Lewis A, Archer CW (1993) Cytokeratin expression in articular chondrocytes. Trans Orthop Res Soc 18:616
Krampera M, Galipeau J, Shi Y, Tarte K, Sensebe L (2013) Immunological characterization of multipotent mesenchymal stromal cells––the International Society for Cellular Therapy (ISCT) working proposal. Cytotherapy 15:1054–1061
Zanoni I, Granucci F (2013) Role of CD14 in host protection against infections and in metabolism regulation. Front Cell Infect Microbiol 3:32
Pilz GA, Braun J, Ulrich C, Felka T, Warstat K, Ruh M, Schewe B, Abele H, Larbi A, Aicher WK (2011) Human mesenchymal stromal cells express CD14 cross-reactive epitopes. Cytometry A 79:635–645
Meachim G, Cornah MS (1970) Fine structure of juvenile human nucleus pulposus. J Anat 107(Pt 2):337–350
Bibby SRS, Fairbank J, Urban MR, Urban JPG (2002) Cell viability in scoliotic discs in relation to disc deformity and nutrient levels. Spine 27:2220–2228
Tew SR, Kwan APL, Hann A, Thomson BM, Archer CW (2000) The reactions of articular cartilage to experimental wounding. Role of apoptosis. Arthritis Rheum 43:215–225
Tibiletti M, Kregar-Velikonja N, Urban JP, Fairbank JC (2014) Disc cell therapies: critical issues. Eur Spine J 23(3):375–384
Yin H, Price F, Rudnicki MA (2013) Satellite cells and the muscle stem cell niche. Physiol Rev 93:23–67
Sharp CA, Roberts S, Evans H, Brown SJ (2009) Disc cell clusters in pathological human intervertebral discs are associated with increased stress protein immunostaining. Eur Spine J 18:1587–1594
Sorrell JM, Lintala AM, Mahmoodian F, Caterson B (1988) Epitope-specific changes in chondroitin sulfate/dermatan sulfate proteoglycans as markers in the lymphopoietic and granulopoetic compartments of developing bursae of fabricius. J Immunol 140:4263–4270
Caterson B, Griffin J, Mahmoodian F, Sorrell JM (1990) Monoclonal antibodies against chondroitin sulphate isomers: their use as probes for investigating proteoglycan metabolism. Biochem Soc Trans 18:820–821
Acknowledgements
The Notch-1 (bTAN20) antibody developed by S Artavanis-Tsakonas was obtained from the Developmental Studies Hybridoma Bank developed under the auspices of the NICHD and maintained by The University of Iowa, Department of the Biology, Iowa City, IA 52242. We are grateful to the Institute of Orthopaedics Ltd, Oswestry and The Henry Smith Charity for funding, all the spinal surgeons at RJAH Orthopaedic Hospital and Mrs. Annie Kerr for assistance with obtaining patient samples.
Conflict of interest
None.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Supplementary material 1 (WMV 17184 kb)
Rights and permissions
About this article

Cite this article
Turner, S., Balain, B., Caterson, B. et al. Viability, growth kinetics and stem cell markers of single and clustered cells in human intervertebral discs: implications for regenerative therapies. Eur Spine J 23, 2462–2472 (2014). https://doi.org/10.1007/s00586-014-3500-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00586-014-3500-y