
Abstract
The rabbit fish is an economically valuable teleost species which lives in shallow coastal waters amongst aquatic plants. Two species of rabbit fish, Siganus sutore and Siganus javus, have been identified in the sea south of Iran. In this study, in order to investigate the histology of the outer layer of the S. javus eye, the eyes of 12 healthy specimens of S. javus were extracted and histologic sections were prepared. The sections were studied under a light microscope after staining with hematoxylin and eosin and Masson's trichrome stains. The outer layer was composed of the cornea cranially and the sclera caudally. The sclera contained an episclera zone and two cartilaginous segments with connective tissue correlation. The cornea included dermal components (stratified cuboidal epithelium, Bowman's membrane, and dermal stroma with occasional flattened cells); the scleral part consisted of two main layers (irregular fibers of connective tissue overlaying the second scleral stroma layer). The fibers of the scleral stroma were the only component present at the posterior part of the cornea. These results reveal that the eye of S. javus does not possess Descemet's membrane or endothelium in the cornea.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The wall of the eyeball is composed of three layers: the outer layer or fibrous tunic, the middle or vascular layer, and the inner or nervous layer. The outer fibrous layer comprises the sclera, covering most of the eye posteriorly and the cornea anteriorly (Banks 1993).
The sclera is often made up of collagenous fibers and may possess ossified cartilage in some teleost species (Kunz 2004). Scleral ossicles or scleral cartilages exhibit a large degree of variation in presence or absence, number, and morphology within the teleost eye (Franz-Odendaal and Hall 2006; Franz-Odendaal 2008). Two distinct scleral ossicle morphologies have recently been identified, and the number of these elements decline from the ancestral arrangement of four scleral ossicles per eye to the present arrangement of two or less scleral ossicles per eye (Franz-Odendaal 2008).
The cornea is a transparent window which mainly consists of an epithelium overlying a basement membrane (Collin and Collin 2001). The density of its surface cells in bony fish ranges from 2,300 to 40,800 cells/ mm2 (Harding et al. 1974). The stroma with sutures and occasional cells (Collin and Collin 1998a) has a modified anterior zone called Bowman's layer, although it is not well developed in some species and it does not exist in all bony fish (Collin and Collin 1998a, 2001, 1997), and an endothelium with a basement membrane known as Descemet's membrane separating it from the stroma (Collin and Collin 2001). Although the histology of fish eyes from numerous species has been undertaken, no study has investigated the histology of the Siganus javus eye. This fish is an economically valuable teleost species which lives in shallow coastal waters with aquatic plants. Two species of rabbit fish have been identified in the sea south of Iran as Siganus sutore and S. javus.
An accurate recognition of fish physiologic conditions and normal morphology is an appropriate way to determine fish culture needs. Information on the morphological, macroscopic, and microscopic features of fish tissues can help develop cellular biology and provide pathologic evaluation of diseases and lesions. Histological research often plays an important role in obtaining these results. Therefore, in this study, the outer layer of the eye of S. javus was investigated.
Materials and methods
Twelve male rabbit fish with an approximate weight of 365 ± 100 g were chosen from a marine fish propagation and growing center in the Bushehr province in southern Iran. All were healthy and in good condition, without any external deformities or lesions. The fish were decapitated and their heads were kept in 10 % formalin solution for 7 days and then the eyes were removed. Routine histological techniques were undertaken, and 6-μm-thick sections were cut. Sections were stained with standard hematoxylin and eosin (H&E) and Masson's trichrome. Histological study was performed using a light microscope, and photographs were taken for detailed illustration of the results.
Results
The sclera was divisible into an episclera zone and a sclera proper zone. The outer layer of the episclera was composed of connective fibers loosely attached to the sclera proper. The inner layer of the sclera consisted of dense connective tissues with two cartilaginous parts continuing over the ora serrata. These two parts were joined together by means of connective fibrils at the posterior of the globe and changed into the corneal connective tissue at the anterior end. The transition from sclera proper to scleral cornea which occurred at the corneoscleral junction or limbus included vessel extension.
The cornea of S. javus had both dermal and scleral components, and the two distinct parts were separated by a distinct region. The outer dermal portion was composed of three layers: (1) an anterior stratified cuboidal epithelium; (2) Bowman's layer, made up of connective bundles; and (3) dermal stroma, which consisted of layers of fibers parallel with each other and the corneal surface. There were transverse connections between adjacent fibers. Flattened cells were scattered among the lamellae identified by Masson's trichrome staining.
The inner scleral stroma included two parts: irregular fibers of connective tissue situated over the scleral stroma consisted of regular dense connective tissue with no cells. The posterior cornea was covered with fibers of scleral stroma. This was the only component present here, and Descemet's membrane and endothelium were not seen (Figs. 1, 2, 3, 4, 5, and 6).

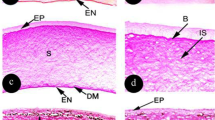
Sagittal section of rabbit fish eye. a, scleral cartilage; b, connective tissue between two cartilages; c, ora serrata; d, limbus; e, cornea. H&E, ×20

Sagittal section of rabbit fish eye. a, scleral cartilage; b, connective tissue between two cartilage areas; c, limbus; d, ora serrata; e, cornea. Masson's trichrome, ×20

Limbus (arrows) in transverse section. H&E, ×400

Sagittal section of the cornea. a, epithelium; b, Bowman's layer; c, dermal stroma; d, separator layer; e, iridescent layer; f, scleral stroma. Masson's trichrome, ×100

Sagittal section of the cornea. a, epithelium; b, Bowman's layer; c, dermal stroma; d, separator layer; e, scleral stroma. H&E, ×400

The cornea. a, Bowman's layer; b, flattened cells. Masson's trichrome, ×400
Discussion
The sclera
Two cartilaginous segments with connective tissue correlation were seen in the sclera of the S. javus eye. The first extensive investigation on the distribution of scleral ossicles in the living teleosts was done by Franz-Odendaal (2008) and suggested that having no ossicles is more common among teleosts. Chondrostei (bichirs, sturgeons, etc.) and Neopterygii (bowfins, gars, etc.) do not have scleral ossicles (Franz-Odendaal and Hall 2006). The oldest actinopterygian fossil has four ossicles; within Chondrostei, the same trend is seen, while modern Chondrostei have none (Franz-Odendaal 2008). Fish without scleral ossicles have a continuous cartilage element within the sclera (Franz-Odendaal et al. 2007). When two parts of the ossicles are present, these two elements are joined to one another by means of a cartilaginous bridge (Franz-Odendaal and Hall 2006).
There is a strong correlation between fish habitats, activity level, and presence/absence of scleral ossicles. Accordingly, fish that inhabit deep sea environments are the most likely to lack scleral ossicles, and 100 % of those that are very active have two ossicles per eye (Franz-Odendaal 2008). Tuna and swordfish, which are fast swimmers, have large bony rings in their eyes (Nakamura and Yamaguchi 1991).
The cornea
The stratified cuboidal epithelium of the dermal cornea in this fish was continuous with the conjunctiva. A corneal epithelium continuous with the conjunctiva and the skin appears to be present in all aquatic vertebrates (Collin and Collin 2001).
A Bowman's layer has been identified in only a few species of bony fish. In spite of the claim by that introduced the Bowman's layer as a normal component of the teleost cornea, it was not present in the species examined by Collin and Collin (1995, 1996, 1998a, b). It is an unorganized membrane in some trout species (Edelhauser and Siegesmund 1968), but it is seen in the cornea of S. javus, at the anterior stroma adjacent to the epithelial cells. This layer is also easily recognizable in the lamprey Petromyzon marinus (Van Horn et al. 1969; Pedersen et al. 1971) and in cartilaginous fish (Keller and Pouliquen 1988).
The two separated parts of the dermal stroma present in the rabbit fish cornea are in accordance with the investigation of Collin and Collin (1995, 1996) on the pipefish Corythoichthyes paxtoni and the salamander fish Lepidogalaxias salamandroides . Aquatic vertebrate corneas possess one stroma, but up to three stromata are present in some deep sea species (Collin and Collin 1998a). The results of this study confirmed the presence of numerous cells in the dermal stroma as is demonstrated in the Florida garfish Lepisosteus platyrhincus (Collin and Collin 1993).
The region separating the two stromata of the S. javus eye occasionally had connective tissue fibrils. This region may be filled with a granular material as in the pipefish (Collin and Collin 1995) or mucoid tissue as in the salamander fish L. salamandroides (Collin and Collin 1994) and the eel Anguilla anguilla (Walls 1963). In the deep-sea gadiforme, Coryphaenoides armatus, which has one layer of dermal stroma and two layers of scleral stroma, a mucoid layer separates the two main parts: dermal and scleral.
In some species, an autochthonous layer which thickens towards the periphery and terminates at the scleral margin has been reported (Collin and Collin 2001). Munk (1968) described the same layer in the cornea of Amia calva in a light microscopy study. Electron microscopy studies demonstrated that the autochthonous layer may be an iridescent layer (Collin and Collin 2001). Corneal iridescence has been observed in some species examined (Collin and Collin 1994, 1988, 1995, 1998a, b). Light microscopic investigation of the S. javus cornea confirmed the presence of a connective tissue layer between the separating layer and the scleral stroma. This finding, when compared with reported data, suggests that this layer may represent an iridescent layer.
Among bony fish, Descemet's membrane as the basement membrane of the corneal endothelium is present in most species (Collin and Collin 2001), but it was not seen in the eye of S. javus. It is also absent in the sea lamprey P. marinus (Van Horn et al. 1969; Pedersen et al. 1971).
Although there is a single layer of endothelial cells in the posterior cornea in bony fish examined by Collin and Collin (1994, 1998a, b, 1988, 1993, 1995, 2000a, b, c), there was no cellular layer in the posterior cornea of the eye of the rabbit fish, and also a complete absence of the endothelium is seen in some cartilaginous fish.
References
Banks WJ (1993) Applied veterinary histology. Mosby, St. Louis, pp 465–468
Collin SP, Collin HB (1988) The cornea of the sandlance, Limnichthyes fasciatus (Creediidae). Cornea 7:190–203
Collin SP, Collin HB (1993) The visual system of the Florida garfish, Lepisosteus platyrhincus (Ginglymodi). II. Cornea and lens. Brain Behav Evol 42:98–115
Collin SP, Collin HB (1994) The fine structure of the cornea of the salamanderfish, Lepidogalaxias salamandroides (Lepidogalaxiidae, Teleostei). Cornea 15:414–426
Collin SP, Collin HB (1995) Ultrastructure and organization of the cornea, lens and iris in the pipefish, Corythoichthyes paxtoni (Syngnathidae, Teleostei). Histol Histopathol 10:313–323
Collin, Collin (1997) The head and eye of the sand lance Limnichthyes fasciatus—a filled emission scanning electron microscopy study. Clin Exp Optom 80:133–138
Collin SP, Collin HB (1998a) A comparative study of the corneal endothelium in vertebrates. Clin Exp Optom 81:245–254
Collin SP, Collin HB (1998b) The deep-sea teleost cornea: a comparative study of gadiform fishes. Histol Histopathol 13:325–336
Collin SP, Collin HB (2000a) A comparative SEM study of the vertebrate corneal epithelium. Cornea 19:218–230
Collin SP, Collin HB (2000b) The corneal endothelium in the blowfish (Torquigener pleurogramma). Cornea 19:231–235
Collin SP, Collin HB (2000c) The corneal surface of aquatic vertebrates: microstructures with optical and nutritional function? Philos Trans R Soc Lond B Biol Sci 355:1171–6
Collin SP, Collin HB (2001) The fish cornea: Adaptation for different aquatic environments. In: Kapoor BG, Hara TG (eds) Sensory biology of jawed fishes: New insights, 1st edn. Science, Enfield, pp 57–96
Edelhauser HF, Siegesmund KA (1968) Ultrastructure of trout cornea. J Fish Res Board Can 25:863–866
Franz-Odendaal TA (2008) Scleral ossicle of teleostei: evolutionary and developmental trends. Anat Rec 291:161–168
Franz-Odendaal TA, Hall BK (2006) Skeletal elements within teleost eye and a discussion of their homology. J Morphol 267:1326–1337
Harding CV, Bagchi M, Weinsieder A, Peters V (1974) A comparative study of the corneal epithelial cell surfaces utilizing the scanning electron microscope. Invest Ophthalmol Vis Sci 13:906–912
Kunz YW (2004) Developmental biology of teleost fishes, 1st edn. Springer, Dordrecht, pp 303–330
Munk O (1968) The eye of Amia and Lepisosteus (Pisces, Holostei) compared with the brachiopterygian and teleostean eyes. Vidensk Meddr Dansk naturh Foren 131:109–127
Pedersen HJ, Van Horn DL, Edelhauser HF (1971) Ultrastructural changes associated with the loss of transparency in the primary spectacle and cornea of spawning sea lamprey. Exp Eye Res 12:147–150
Van Horn DL, Edelhauser HF, Schultz RO (1969) Ultrastructure of the primary spectacle and the cornea of the sea lamprey. J Ultrastruct Res 26:454–464
Walls GL (1963) The vertebrate eye and its adaptive radiation. Hafner, New York, pp 563–588
Acknowledgments
This project was funded by the Shahid Bahonar University of Kerman. The authors are thankful to Mr. Saeed Hasanzadeh for his assistance in the sampling process and his technical support.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
sadat Mansoori, F., Sattari, A., Kheirandish, R. et al. A histological study of the outer layer of rabbit fish (Siganus javus) eye. Comp Clin Pathol 23, 125–128 (2014). https://doi.org/10.1007/s00580-012-1582-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00580-012-1582-7