
Abstract
Ericoid mycorrhizas are associated with a number of host plants in the Ericaceae in high-elevation regions of Taiwan. The ability of these microorganisms to thrive in harsh environmental conditions in the regions implies their capability of decomposing plant organic matter (raw humus). The objective of this study was to investigate the decomposition characteristics of three ericoid mycorrhizal endophytes isolated from the roots of Formosan rhododendron (Rhododendron formosanum Hemsl.). Molecular analysis indicated that strains Rf9 and Rf32 belong to the genus Cryptosporiopsis while strain Rf28 is a member of the genus Phialocephala. Mycorrhizal synthesis experiment showed that the roots of synthesized seedlings produced hyphal coils, a characteristic of ericoid mycorrhiza. Decomposition ability analysis revealed that strains Rf28 and Rf32 had the highest rates of decomposition of organic matter (up to 10.4% after 70 days) while the value for strain Rf9 was about 6.8%. Consistently, these strains secreted extracellular oxidases when cultured on tannic acid medium. Enzyme assay revealed that strains Rf28 and Rf32 secreted peroxidase, laccase, tyrosinase, and cellulase, but strain Rf9 secreted mainly peroxidase and tyrosinase. Apparently, the differences in secreted hydrolytic enzymes among the three endophytes are related to their ability to decompose organic matter. In the mycorrhizal synthesis experiment, all inoculated seedlings survived in the organic matter substrate for 70 days and exhibited a stronger vigor than the control. This study demonstrated that these three isolated endophytes, Rf9, Rf28, and Rf32, are ericoid mycorrhizal fungi, capable of forming ericoid mycorrhiza with Formosan rhododendron. Meanwhile, all three endophytes can secrete hydrolytic enzymes to decompose organic matter for growth, presumably a prerequisite for the adaptation of Formosan rhododendron to the harsh environments of high elevation.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Ericoid mycorrhizal associations predominate the plant communities of the Ericaceae in high-elevation regions (Read 1991). The harsh environmental conditions of these regions hinder decomposition of dead plant tissues, resulting in soils that are acidic (low pH) and rich in organic matter (raw humus) but deficient in nitrogen and phosphorus (Straker and Mitchell 1986).
Functional groups of fungi in soils are usually separated into two groups, decomposers and fungal mutualists. The former can degrade organic matter and release nutrients while the latter can absorb ions from mineralization during the decomposition of organic matter. Pezoloma ericae (Read) Baral (Leotiaceae) (Baral and Krieglsteiner 2006) and Oidiodendron maius Barron (Couture et al. 1983; Dalpé 1986; Douglas et al. 1989) are ericoid mycorrhizal fungi (ERMFs), symbionts with ericaceous plants, and also can significantly degrade organic matter. ERMFs absorb nutrients from soils rich in phenolic compounds and organic matter and also transport the absorbed nutrients to host plants to support their growth (Read and Perez-Moreno 2003).
Thus, the ability to accelerate the decomposition of organic matter has been confirmed in ERMFs. ERMFs are capable of secreting two types of enzymes. The first type can degrade cellulose, hemicellulose, lignin, polyphenols, and tannic acid, which can accelerate the release of necessary nutrients. The second type promotes direct absorption of necessary nutrients (Cairney and Burke 1998; Read and Perez-Moreno 2003).
A variety of polyphenolic compounds can be found in classical soils of heathlands (Jalal and Read 1983). In the habitats of Calluna vulgaris L., where the surface layer is rich in organic matter, there are over 30% phenolic compounds by dry weight (Jalal et al. 1982). Other studies indicated that ERMFs use many phenolic compounds as a carbon source, but also release cellulase, hemicellulase, laccase, catechol oxidase, and pectinase to degrade cellulose, hemicellulose, lignin, and polyphenols (Cairney and Burke 1998; Read and Perez-Moreno 2003).
Formosan rhododendron (Rhododendron formosanum Hemsl.) is an endemic species of Taiwan, but its coriaceous foliage is difficult to decompose. The characteristics of its habitat are acidic soil, high humidity, low temperatures, and a shortage of available mineral nutrients such as nitrogen (N) and phosphorus (P). Major nutrients such as N and P are compounded by organic complexes, which causes restricted mineralization, and therefore, cannot be used by plants. However, ericoid mycorrhizas (ERMs) play important roles in combining and releasing nutrients in this habitat (Johansson 2001). Previous studies showed that ERMFs play a key role in nutrient cycles in heathlands (Cairney and Meharg 2003). In Formosan rhododendron forests, organic matter accumulates on the forest floor. However, the decomposition mechanism of organic matter in the habitats of Formosan rhododendron remains unknown. The purpose of this study was to examine the ability of ericoid mycorrhizal endophytes of Formosan rhododendron to decompose organic matter.
Materials and methods
Strains of mycorrhizas
Three endophytes were isolated previously from the roots of R. formosanum (120°47′31.35″ E, 23°38′9.85″ N), located in the Sanlinchi Recreational Park, Chushang Township, Nantou County, Taiwan (Lin et al. 2010a). Specimens of Rf9 (Lin et al. 2010b), Rf28 (Lin et al. 2010c), and Rf32 were deposited at the Forest Mycobiont Laboratory of National Chiayi University. These three endophytes also have been deposited in the Bioresource Collection and Research Centre (BCRC) (Rf9, Rf28, and Rf32 isolates: BCRC 34764, 34762, and 34763, respectively) and their internal transcribed spacer (ITS) genomic sequences were deposited in the GenBank (Rf9, Rf28, and Rf32 isolates: HQ260953, HQ260954, and HQ260955, respectively).
Preparation and analysis of organic matter
Organic matter was collected randomly from the O horizon of Formosan rhododendron forestland. Coarse debris, such as roots and twigs, was removed prior to drying the material, which was then homogenized in a blender for analysis after being dried (Piercey et al. 2002). Ashing (Moore and Chapman 1986) and dry combustion method (Chou 2003) were used to analyze the contents of K, Ca, Mg, Na, and total carbon and nitrogen.
Extraction of organic matter and determination of total phenolics
Preparation of organic matter and determination of total phenolic contents were based on the Folin–Ciocalteu method, as described previously (Huang et al. 2003; Kujala et al. 2000).
DNA extraction, sequencing, and phylogenetic analysis
The methods described by Sigler et al. (2005) were followed for ITS recombinant DNA (rDNA) analysis. Mycelium for DNA extraction, amplification, and sequencing was scraped from the surfaces of potato dextrose agar (PDA) cultures. Genomic DNA was extracted using a Puregene Proteinase K. Total fungal DNA was used as template for amplification with primers ITS1-F and TW13 (Sigler et al. 2005). PCR products were sequenced by Genomics BioSci and Tech Company. Sequences were assembled and related sequences were analyzed using BLAST searches.
The phylogenetic relationships were analyzed by molecular evolutionary genetics analysis. Bootstrapping was performed using neighbor-joining.
Resynthesis
Pure resynthesis was done by following the method of Dalpé (1986). After surface cleaning, the seeds of Formosan rhododendron were sterilized with 10% sodium hypochlorite solution for 15 min, rinsed three times with sterilized distilled water, and then incubated in test tubes containing 1% agar for germination. Germinated seedlings were transplanted to modified Mitchell and Read (MMR) medium (NH4Cl, 32 mg L−1; CaCl2‧7H2O, 43.5 mg L−1; MgSO4‧7H2O, 10 mg L−1; KCl, 5.5 mg L−1; FeCl3, 3.75 mg L−1; sucrose, 2 g L−1; KH2PO4, 210 mg L−1; pyridoxine, 100 μg L−1; thiamine, 100 μg L−1; agar, 10 g L−1). Seven days after germination, the aseptic seedlings were inoculated with Rf9, Rf28, and Rf32 fungal strains, respectively, and grown in a growth chamber (22°C, 65% RH and 16 h light with a maximum illumination of 5,000 lx).
Analysis of decomposition ability
The method for evaluation of organic matter decomposition by the mycorrhizas was modified from that of Piercey et al. (2002). The decomposition ability of the three endophytes was determined by placing a 0.5-mm3 plug of mycelium of each endophyte onto a 2-g dry weight organic matter moistened with 10 mL d-H2O in glass jar (95 × 73 mm). Four replicates were prepared for each endophyte and four uninoculated plates served as control. Plates were sealed with parafilm and incubated at 25°C in the dark for 70 days. To ensure that plates had remained uncontaminated during the incubation period, a small sample (<0.01 g) of organic matter taken at three time points from each jar was placed on PDA medium (39 g L−1) and incubated for 28 days. After 70 days of culture, the jars with substrate were autoclaved (liquid cycle at 121°C for 20 min), dried to a constant mass at 38°C, and weighed. Mass losses were expressed as percentages.
Observation of colonization in the roots system of seedlings
After 70 days of incubation, the seedlings were taken out, and the roots were stained with aniline blue for observation of hyphae (Upson et al. 2007).
Determination of extracellular enzymes
For detecting the activity of extracellular oxidase, small mycelial pieces (of 3 mm in diameter) of each endophyte cultured on 2% malt extract agar (MEA) medium (20 g L−1 malt extract agar and 15 g L−1 agar) were inoculated onto new MEA medium (containing 0.5% tannin acid) and maintained at 20°C in a growth chamber. Observations were made after 7 days of culture. If the medium turned brown, it was determined that the endophyte had secreted extracellular oxidase (Käärik 1965; Stalpers 1978; Taylor 1974).
For detecting the activity of peroxidase and tyrosinase, small mycelial pieces (3 mm in diameter) of each endophyte cultured on MEA media were inoculated onto new MEA media and maintained at 20°C in a growth chamber. After 7 days of incubation, different solutions were used to detect the activity of different hydrolytic enzymes after 3, 27, and 72 h (Käärik 1965; Stalpers 1978; Taylor 1974). For peroxidase, a mixture of 4% H2O2 and a 1% pyrogallol (1:1, v/v) solutions was added to the media. If the medium turned yellow, it suggested that the endophyte had secreted peroxidase. For tyrosinase, 1 M P-cresol solution was added to the media, and if the medium turned brownish orange, the endophyte had secreted tyrosinase.
For detecting the activity of laccase, the mycelial pieces were inoculated onto PDA medium (containing 100 ppm Congo red, Sigma, St. Louis, MO, USA) and maintained at 20°C in a growth chamber. Observations were made after 7 days of culture. If the medium turned transparent, it suggested that the endophyte had secreted laccase (Sai 2005).
For detecting the activity of cellulase, the small mycelial pieces from 2% MEA medium were inoculated onto cellulose media (KH2PO4, 1 g L−1; (NH4)2SO4, 1 g L−1; MgSO4‧7 H2O, 0.2 g L−1; CaCl2‧H2O, 0.1 g L−1; FeCl3‧H2O, 0.033 g L−1; carboxymethyl cellulose, 5 g L−1; peptone, 0.1 g L−1; yeast extract, 0.05 g L−1; agar, 18 g L−1) and maintained at 20 ± 1°C in a growth chamber. After 25 days, a 0.1% Congo red solution and a 5 N NaCl solution were added to the media, respectively, and if the medium had turned yellowish brown, it suggested that the endophyte had secreted cellulase (Wu 2004).
Statistical analyses
Statistical analysis was performed by using the software Statistical Package for the Social Science (SPSS 12.0) (IL, USA) for windows program. All data represent means of 4 separate experiments ± standard error (n = 4). Differences among endophytes were analyzed by Turkey’s multiple range test at p ≤ 0.05 significant level.
Results and discussion
Molecular phylogenetic analysis
Although numerous sterile fungi have been isolated from the roots of Ericaceae and Epacridaceae, they have not been identified to the species level due to their inability to form telemorph and conidium (Burgeff 1961; Peretto et al. 1990; Stoyke and Currah 1991; Hutton et al. 1994; Xiao and Berch 1996; Hambleton and Currah 1997). However, in recent years, ITS of rDNA has been successfully used to clarify the phylogenetic relationships and exhibit the genetic diversity of ERMF (Liu et al. 1998; Chambers et al. 1999; McLean et al. 1999; Monreal et al. 1999; Sharples et al. 2000; Usuki et al. 2003). Clearly, ITS analysis has contributed to the research of ERMF.
Taxonomic affinities are assigned to Rf9 and Rf32 based on BLAST sequence similarity analysis (http://blast.ncbi.nlm.nih.gov/Blast.cgi) including several most closely matched sequences. The ITS sequences of Rf9 and Rf32 can be matched to those of Ericoid mycorrhizal sp. ppo-7 (AY599245) and Cryptosporiopsis ericae (AY540126), and these species can be grouped (69% bootstrap) in the neighbor-joining analysis (Fig. 1). The ITS sequence of Rf28 has a close match to those of Phialocephala fortinii (AY394915) and Trimmatostroma salicis (EU019300); therefore, they are grouped in the neighbor-joining analysis (98% bootstrap), but Rf28 is closer to P. fortinii (AY394915) (Fig. 1). Our molecular analyses also included the nearest genera (Acremonium, Corynespora, Dermea, Hyphodiscus, Meliniomyces, Neofabraea, Oidiodendron, Pezoloma, Phialemonium, Rhizoscyphus, and Trimmatostroma) (http://www6.ufrgs.br/favet/imunovet/molecular_immunology/physiofungitaxonomy.html) because Cryptosporiopsis species are the anamorphs of Pezicula (Verkley et al. 2003). The neighbor-joining analysis (Fig. 1) suggests that endophytes of Rf9, Rf28, and Rf32 are not closely related to the species of uncultured Acremonium (GU055562), Rhizoscyphus ericae (AY394907), Phialemonium dimorphosporum (HM060271), Pezoloma sp. (AB506027), Pezizella amenti (AJ430398), Pezicula ocellata (AF141199), O. maius (FN662651), Neofabraea alba (AF141190), Meliniomyces variabilis (AY838789), Hyphodiscus hymeniophilus (DQ227258), Hymenoscyphus ericae (AY606310), Corynespora cassiicola (GU296145), and Dermea viburni (AF141163). The latter six species are clearly in a separate clade. In our previous observations of colony and hypha (Lin et al. 2010b, c), these endophytes showed different features than either C. ericae or P. fortinii. Thus, strain Rf28 can be considered a new species belonging to Phialocephala whereas strains Rf9 and Rf32 maybe new species of the genus Cryptosporiopsis.

Neighbor-joining phylogenetic tree based on rDNA ITS sequence data from endophytes of Rf9, Rf28, and Rf32 isolated from the root systems of Rhododendron formosanum with known ericoid endophytes and selected fungal species from GenBank with high sequence similarities. Numerical values above the branches indicate bootstrap percentiles from 1,000 replicates. Bootstrap numbers over 50% are indicated. Horizontal branch lengths are proportional to the scale of basepair substitutions
Features of the organic matter of the O horizon from the natural habitat
The important features of the organic matter from a Formosan rhododendron forest in the Sanlinchi area include pH 3.1, 49.6% total C, 1.08% total N, 32% total phenol, 1.31 cmol(+) kg−1 Ca, 0.72 cmol(+) kg−1 Mg, 0.89 cmol(+) kg−1 K, and 0.16 cmol(+) kg−1 Na. The results indicate that the organic matter of the habitat is extremely acidic with a high C/N ratio close to 50 and consisted of over 30% of the total phenolic compounds. All these features suggest that the organic matter in this habitat is difficult to be decomposed. Read et al. (2004) concluded that this type of organic matter has a very low mineralization ratio and barely any nitrification. Therefore, in order to survive in such a harsh environment, plants need the assistance of mycorrhiza for the acquisition of nutrients.
Morphology and colonization in resynthesized seedlings
After 70 days of incubation, all seedlings survived; however, the control seedlings showed chlorotic symptoms and their root growth was limited (Fig. 2). The resynthesized seedlings, cultured in the same organic matter, exhibited a strong vigor in growth than the control seedlings.

Morphology of Formosan rhododendron seedlings after culture on the MMR medium for 70 days. a Seedlings inoculated with Rf9. b Seedlings inoculated with Rf32. c Seedlings inoculated with Rf28. d Controls
A feature of hyphal coils of ERM was observed in the root systems of resynthesized seedlings (Fig. 3). On the contrary, this feature was not observed in the control seedlings. Therefore, these three endophytes could associate with Formosan rhododendron, and the root associations had clear features of ERMs. The colonization feature of these three endophytes further supports their classification to ERMFs.

Root structure of Formosan rhododendron seedlings after culture on the MMR medium for 70 days. a Seedling inoculated with Rf9. b Seedling inoculated with Rf32. c Seedling inoculated with Rf28. d Control
Ability to decompose organic matter
-
1.
Strains differences: As shown in Table 1 and Fig. 4, after incubation for 70 days, Rf28 and Rf32 caused the highest mass losses of organic matter, 10.38 ± 0.01% and 10.38 ± 0.01%, respectively, as compared to Rf9 (6.75 ± 0.01%). A small decrease in mass (4.91 ± 0.42%) was noted in the control plates, and contaminating organisms were not found. Mass losses in all treatments were significantly higher than that of the control (p < 0.005). The results showed that the Rf28 and Rf32 endophytes are capable of decomposing organic matter, with rates comparable to that of O. maius Barron at 11.65 ± 1.13% (Piercey et al. 2002).
Table 1 Mass loss percentages of organic matter in medium inoculated with the three endophytes and the control (without inoculation) following a 70-day incubation Fig. 4 
Mass loss of organic matter (percent) after culture for 70 days in substrate inoculated with three endophytes and the control. Values with the same letter are not significantly different (p = 0.05)
In our previous studies (Lin et al. 2010b, c), we have cultivated these three strains of endophytes in a MEA medium at 22°C for 12 days and found that the Rf9 strain grows at a faster rate (4.8 mm/day) than Rf28 (2.8 mm/day) and Rf32 (2.1 mm/day). However, when these strains were cultivated in an organic substrate of their habitat, Rf32 and Rf28 strains had the highest rates of organic matter loss, followed by Rf9 (Fig. 4, Table 1). This trend is in contrast to the case of strains living in a MEA medium, but does correspond to the enzyme activity results, which indicates that Rf32 and Rf28 strains can release multiple enzymes that can facilitate the decomposition of polyphenolic complex (see next section on extracellular enzymes). Hence, it is reasonable to expect them to exhibit the most significant organic matter losses. The rate of organic matter loss of the control group in this study (4.9%) is comparable to that of the control group of Piercey et al. (2002) (4.4%), which is presumably associated with the hydrolysis of water-soluble ingredients in the organic matter, as contamination has been excluded as a cause.
Clearly, these endophytes all have the ability to decompose organic matter. This ability could benefit the growth of both endophytes and their host plants. Read (1984) concluded that the reason for the communities dominated by Ericaceae is probably due to the strong acidity and rich contents of phenolic compounds. This study demonstrates that ERMFs play a key role in the decomposition of organic matter and nutrient cycling in the Formosan rhododendron forests, which may be essential for the adaptation of Formosan rhododendron to the harsh environments of high elevation.

-
2.
Resynthesis and control seedlings: After 70 days of incubation (Table 2, Fig. 5), seedlings inoculated with Rf9 and Rf32 caused the greatest mass losses of organic matter (10.98 ± 0.19% and 10.88 ± 0.17%, respectively), as compared with Rf28 (8.73 ± 0.83%). A small decrease in mass (4.55 ± 0.35%) was noted in control plates, but contaminating organisms were not found. Mass losses in all treatments were significantly higher than that of the control (p < 0.005). The results suggest that the association is critical for the host plants to acquire nutrients.
Table 2 Mass loss percentages of organic matter by resynthesis and control seedlings following a 70-day incubation Fig. 5 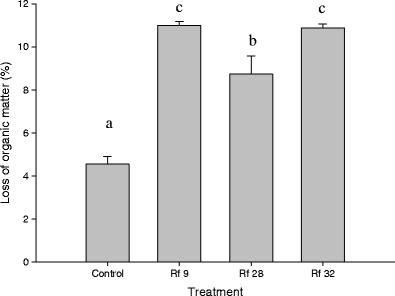
Mass loss of organic matter (percent) from substrate after culture with inoculated or control seedlings for 70 days. Values with the same letter are not significantly different (p = 0.05). Control: non-inoculated seedlings
In the strain comparison (Fig. 4), strains Rf28 and Rf32 performed better than Rf9 whereas in the resynthesis experiment (Fig. 5) seedlings inoculated with Rf9 and Rf32 grow more vigorously than seedlings inoculated with Rf28. The discrepancy may be explained by the differences in the affinity of the endophytes to the host plants. The resynthesis experiments were conducted by moving inoculated seedlings to sterile substrate, where it grew directly on the substrate, rather than by adding a fungus plug to the substrate for inoculation. Under such condition, the mass loss would be an indicative of the ability of the resynthesis, rather than the ability of the strain or the host plant.

Extracellular enzymes
The extracellular enzymes secreted from the three endophytes were analyzed by their ability to react with the substrates on the culture media after 25 (cellulase) or 7 (other enzymes) days of culture.
-
1.
Extracellular oxidase: When the three endophytes were cultured on MEA media containing tannic acid (Fig. 6), they turned the media dark-brown, which indicates that they are all capable of secreting extracellular oxidative enzymes into the medium (Käärik 1965; Stalpers 1978; Taylor 1974).
-
2.
Peroxidase: When a mixture of H2O2 and pyrogallol was sprayed over the 2% MEA media with the three endophytes, no reactions were detected after 3 h, but the media turned yellow after 24 h (data not shown), indicating the activity of peroxidase secreted by the three endophytes (Käärik 1965; Stalpers 1978; Taylor 1974).
-
3.
Tyrosinase: When the cultures on 2% MEA media were sprayed with P-cresol solution, only the media cultured with Rf28 had turned brownish orange after 3 h, but all the media had turned brownish orange after 72 h (data not shown). These results demonstrate that tyrosinase had been secreted by all three endophytes, but Rf28 is capable of secreting tyrosinase at an earlier stage (Käärik 1965; Stalpers 1978; Taylor 1974).
-
4.
Laccase: When the strains Rf28 and Rf32 were cultured on PDA media containing 100 ppm Congo red, the media had turned transparent after 7 days, but the media with Rf9 showed no signs of reaction (data not shown). This indicates that only Rf28 and Rf32 are capable of secreting laccase into the medium (Sai 2005).
-
5.
Cellulase: When Congo red and NaCl solutions were sequentially sprayed over the media to detect cellulase activity, the media with Rf28 and Rf32 turned yellowish brown, but the media with Rf9 showed no signs of reaction (Fig. 7). The results suggest that only Rf28 and Rf32 secreted cellulase (Wu 2004).

Representative MEA plates containing tannin acid after incubation of three endophytes for 21 days. The medium changing to a brown coloration indicated production of extracellular oxidase. a Rf9 strain. b Rf28 strain. c Rf32 strain

Representative MEA plates containing Congo red after incubation with three endophytes for 25 days. The medium changing to a yellowish-brown coloration indicated production of cellulase. a Rf9 strain. b Rf28 strain. c Rf32 strain
Taken together, these results suggest that Rf9 is capable of secreting peroxidase and tyrosinase while Rf28 and Rf32 are capable of secreting laccase and cellulose in addition to peroxidase and tyrosinase. Apparently, these enzymes promote the decomposition of organic matter and release available nutrients for the growth of both endophytes and seedlings of Formosan rhododendron. The resynthesized seedlings not only have the features of ERMs but also are capable of decomposing more organic matter than the control seedlings to provide nutrients needed for the growth of the endophytes themselves and the host plants as well. Our findings are consistent with the conclusion by Cairney and Meharg (2003) that mycorrhizal associations in their various forms are naturally ubiquitous and play important roles in plant nutrition and nutrient cycling.
Conclusions
The results of this study confirmed the association of three endophytes with the roots of Formosan rhododendron. Molecular analysis allows the classification of the three endophytes into two clusters: both Rf9 and Rf32 are classified into the genus Cryptosporiopsis while Rf28 is a member of the genus Phialacephala (Fig. 1). Furthermore, all these three endophytes belong to ERMFs (Fig. 3). The organic matter from the habitat of Formosan rhododendron in the Sanlinchi area is known to be very acidic (pH 3.1) and consisted of over 30% phenolic compounds. The ability of these endophytes to secrete many kinds of extracellular enzymes, such as peroxidase, tyrosinase, laccase, and cellulase (Fig. 7) for accelerated decomposition of the organic matter, allows the host plants to thrive in the harsh habitat. Among these three endophytes, strain Rf32 performs most significantly in the substrate of its natural habitat, regardless of self decomposition or symbiosis with R. formosanum.
References
Baral HO, Krieglsteiner L (2006) Hymenoscyphus subcarneus, a little known bryicolous discomycete found in the Bialowieza National Park. Acta Mycol 41:11–20
Burgeff H (1961) Mikrobiologie des Hochmoores. Fischer-Verlag, Stuttgart, p 197
Cairney JWG, Burke RM (1998) Extracellular enzyme activities of the ericoid mycorrhizal endophyte Hymenoscyphus ericae (Read) Korf & Kernan: their likely roles in decomposition of dead plant tissue in soil. Plant Soil 205:181–192
Cairney JWG, Meharg AA (2003) Ericoid mycorrhiza: a partnership that exploits harsh edaphic conditions. Eur J Soil 54:735–740
Chambers SM, Williams PG, Seppelt RD, Cairney JWG (1999) Molecular identification of a Hymenoscyphus spp. from rhizoids of the leafy liverwort Cephaloziella exiliflora in Australia and Antarctica. Mycol Res 103:286–288
Chou YJ (2003) Seasonal dynamics of litterfall, recycling amount of nutrients and decomposition rates in Hui-Sun Experimental Forest [Master’s thesis]. Taipei, Taiwan: Department of Forestry, National Chung-Hsing Univ [in Chinese]
Couture M, Fortin JA, Dalpé Y (1983) Oidiodendron griseurn Robak: an endophyte of ericoid mycorrhiza in Vaccirlium sp. New Phytol 95:375–380
Dalpé Y (1986) Axenic synthesis of ericoid mycorrhiza in Vaccinium angustifoliurn Ait. by Oidiodendron species. New Phytol 103:391–396
Douglas GC, Heslin MC, Reid C (1989) Isolation of Oidiodendron maius from Rhododendron and ultrastructural characterization of synthesized mycorrhizas. Can J Bot 67:2206–2212
Hambleton S, Currah RS (1997) Fungal endophytes from the roots of alpine and boreal Ericaceae. Can J Bot 75:1570–1581
Huang CY, Wu JH, Chang CS, Yeh YL, Chang ST (2003) Influence of manufacturing processes and harvesting seasons on the antioxidant activity of tea extracts. J Exp For Natl Taiwan Univ 17(4):231–237
Hutton BJ, Dixon KW, Sivasithamparam K (1994) Ericoid endophytes of Western Australian heaths (Epacridaceae). New Phytol 127:557–566
Jalal MAF, Read DJ (1983) The organic acid composition of Calluna heathland soil with special reference to phyto- and fungi-toxicity. I. Isolation and identification of organic acids. Plant Soil 70:257–272
Jalal MAF, Read DJ, Haslam E (1982) Phenolic composition and its seasonal variation in Calluna vulgaris. Phytochemistry 21:1397–1401
Johansson M (2001) Fungal associations of Danish Calluna vulgaris roots with special reference to ericoid mycorrhiza. Plant Soil 231:225–232
Käärik A (1965) The identification of the mycelia of wood-decay fungi by their oxidation reactions with phenolic compounds. Stud For Suec 31:1–80
Kujala TS, Loponen JM, Klika KD, Pihlaja K (2000) Phenolics and betacyanins in red beetroot (Beta vulgaris) root: distribution and effect of cold storage on the content of total phenolics and three individual compounds. J Agric Food Chem 48:5338–5342
Lin LC, Lee MJ, Wang YZ (2010a) Morphology and ultrastructure of root-fungus association of Rhododendron formosanum Hemsl. in Sanlinchi forest of central Taiwan. Taiwan J Biodivers 12(2):93–101
Lin LC, Lee MJ, Wang YZ (2010b) Rhododendron formosanum symbiosed with endophytic fungi Cryptosporiopsis sp. to form Ericoid mycorrhiza. Q J Chin Forest 43(1):171–179 [in Chinese]
Lin LC, Lee MJ, Chen JL (2010c) Axenic synthesis of ericoid mycorrhiza in Rhododendron formosanum with Phialocephala species. Taiwan J For Sci 25(3):211–218
Liu G, Chambers SM, Cairney JWG (1998) Molecular diversity of ericoid mycorrhizal endophytes isolated from Woollsia pungens. New Phytol 140:145–153
McLean CB, Cunnington JH, Lawrie AC (1999) Molecular diversity within and between ericoid endophytes from the Ericaceae and Epacridaceae. New Phytol 144:351–358
Monreal M, Berch SM, Berbee M (1999) Molecular diversity of ericoid mycorrhizal fungi. Can J Bot 77:1580–1594
Moore PD, Chapman SB (1986) Methods in plants ecology, 2nd edn. Blackwell, Oxford
Peretto R, Perotto S, Faccio A, Bonfante P (1990) Cell surface in Calluna vulgaris L. hair roots in situ localization of polysaccharide components. Protoplasma 155:1–18
Piercey MM, Thormann MN, Currah RS (2002) Saprobic characteristics of three fungal taxa from ericalean roots and their association with the roots of Rhododendron groenlandicum and Picea mariana in culture. Mycorrhiza 12:175–180
Read DJ (1984) Interaction between ericaceous plants and their competitors with special reference to soil toxicity. Asp Appl Biol 5:95–209
Read DJ (1991) Mycorrhizas in ecosystems. Experientia 47:376–391
Read DJ, Perez-Moreno J (2003) Mycorrhizas and nutrient cycling in ecosystems—a journey towards relevance? New Phytol 157:475–492
Read DJ, Leake JR, Perez-Moreno J (2004) Mycorrhizal fungi as drivers of ecosystem processes in heathland and boreal forest biomes. Can J Bot 82:1243–1263
Sai IH (2005) Optimization of growth condition and laccase production by white rot fungi [Master’s thesis]. Taipei, Taiwan: Department of Microbiology, Soochow Univ [in Chinese]
Sharples JM, Chambers SM, Meharg AA, Cairney JWG (2000) Genetic diversity of root-associated fungal endophytes from Calluna vulgaris at contrasting field sites. New Phytol 148:153–162
Sigler L, Allan T, Lim SR, Berch S, Berbee M (2005) Two new Cryptosporiopsis species from roots of ericaceous hosts in western North America. Stud Mycol 53:53–62
Stalpers JA (1978) Identification of wood-inhabiting Aphyllophorales in pure culture. Centraalbureau Voor Schimmelcultures, Baarn. Stud Mycol 16:1–248
Stoyke G, Currah RS (1991) Endophytic fungi from the mycorrhizae of alpine ericoid plants. Can J Bot 69:347–352
Straker CJ, Mitchell DT (1986) The activity and characterization of acid phosphatase in endomycorrhizal fungi of the Ericaceae. New Phytol 104:243–256
Taylor JR (1974) Biochemical test for identification of mycelial culture of Basidiomycetes. Ann Appl Biol 78:113–123
Upson R, Read DJ, Newsham KK (2007) Microscopy analyses of field-collected Cephaloziella varians. New Phytol 176:460–471
Usuki F, Abe PJ, Kakishima M (2003) Diversity of ericoid mycorrhizal fungi isolated from hair roots of Rhododendron obtusum var. kaempferi in a Japanese red pine forest. Mycoscience 44:97–102
Verkley GJM, Zijlstra JD, Summerbell RC, Berendse F (2003) Phylogeny and taxonomy of root-inhabiting Cryptosporiopsis species, and C. rhizophila sp. nov., a fungus inhabiting roots of several Ericaceae. Mycol Res 107:689–698
Wu HT (2004) Screening, identification and basic properties of multienzyme producing microorganism [Master’s thesis]. Taichung, Taiwan: Department of Applied Chemistry, Chaoyang Univ of Technology [in Chinese]
Xiao G, Berch SM (1996) Diversity and abundance of ericoid mycorrhizal fungi of Gaultheria shallon on forest clearcuts. Can J Bot 74:337–346
Acknowledgment
The authors gratefully acknowledge the financial support from the National Chiayi University.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Lin, LC., Lee, MJ. & Chen, JL. Decomposition of organic matter by the ericoid mycorrhizal endophytes of Formosan rhododendron (Rhododendron formosanum Hemsl.). Mycorrhiza 21, 331–339 (2011). https://doi.org/10.1007/s00572-010-0342-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00572-010-0342-2