
Abstract
We investigated the species identity of mat-forming ectomycorrhizal (EM) fungi associated with old- and second-growth Douglas-fir stands. Using molecular analyses of rhizomorphs and EM root tips, we characterized 28 unique internal transcribed spacer sequences and considered them proxies for mat-forming EM species. In both stand age classes, one Athelioid species in the genus Piloderma dominated our sample of the mat-forming fungal community. In second-growth stands, the second most frequently encountered mat-forming EM species belonged to the genus Hysterangium. In old-growth stands, several Ramaria species were associated with a frequently encountered mat morphology but no species dominated the community. After using rarefaction analysis to standardize sampling effort, the total species richness did not differ statistically between old- and second-growth habitats. Both an abundance of infrequently encountered species and incomplete sampling of the mat-forming EM community may have limited our ability to detect potential differences in species richness. Several frequently encountered Piloderma species appear to have broad (holarctic) distributions and diverse host associations and their potential importance in forest ecosystems warrants further study.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Ectomycorrhizal (EM) fungi play an influential role in forest community dynamics because they form beneficial symbioses with many dominate trees and augment nutrient uptake by their host plants (Smith and Read 1997). Prominent within several forest ecosystems are subsets of EM fungi that colonize soil and litter layers with dense, perennial mats (Griffiths et al. 1991a; Agerer 2001). Research within small forest plots (e.g., 25 m2 × 10 cm depth) has shown that individual EM species can colonize up to 16% of the local soil volume (Hintikka and Naykki 1967; Cromack et al. 1979; Griffiths et al. 1996) with hyphal and rhizomorphic mats that constitute up to 50% of the dry weight of associated soils (Ingham et al. 1991). Because of their ability to dominate soils locally, mat-forming EM fungi are hypothesized to be important participants in forest soil nutrient cycles wherever they are abundant.
Mats formed by EM fungi aggregate soil and alter its appearance in a variety of ways. The resulting mat-associated soil morphologies are thought to signify different soil exploration strategies (Agerer 2001) and have been linked to various soil processes (Griffiths et al. 1996). For example, soil morphologies commonly produced by mats in Pacific Northwest (PNW) Douglas-fir (Pseudotsuga menziesii) forests include pale, hydrophobic, powdery soils associated with EM fungi that dominate mineral horizons and organic soil horizons and litter tightly aggregated by rhizomorphs of EM species (Griffiths et al. 1991a, 1996). Past research has demonstrated that these morphologies are associated with physical and chemical variation in soil properties (Cromack et al. 1979; Griffiths et al. 1990, 1991b, 1994) that have significant impacts on soil microbial diversity (Cromack et al. 1988). Fungi that form mats are physiologically capable of using protein bound nitrogen and polyphenol-complexed protein as nitrogen sources (Griffiths and Caldwell 1992) allowing access to otherwise recalcitrant nitrogen pools (Perez-Moreno and Read 2000) and providing nitrogen to host trees without the need of microbial mineralization (Näsholm et al. 1998). Fungal mats also produce high concentrations of oxalic acid (Griffiths et al. 1994), leading to increased soil and rock weathering (Blum et al. 2002). The abundance, discrete composition, easy detection, and biological properties of mats make them attractive subjects for experimental research on the roles EM fungi play in soil ecosystems (Griffiths and Caldwell 1992; Griffiths et al. 1996). Identifying the species that compose this interesting assemblage of fungi is fundamental to the design and interpretation of research into their ecological functions.
Genera known to contain mat-forming EM species include Bankera, Boletopsis, Gautieria, Geastrum, Gomphus, Hydnellum, Hysterangium, Phellodon, Piloderma, Ramaria, and Sarcodon (Agerer 2001 and references therein). In the PNW, mat-forming EM fungi have commonly been associated with two genera, Gautieria (Griffiths et al. 1991a) and Hysterangium (Cromack et al. 1979) that represent two independent lineages within the Phallales/Gomphales clade (Castellano 1988; Hibbett et al. 1997; Humpert et al. 2001). Past identification of PNW mat-forming EM species was based on mat morphology coupled with associated hypogeous fruit bodies. However, Griffiths et al. (1996) observed that mat morphologies commonly associated with the genus Hysterangium likely are produced by a more diverse set of EM taxa. Similarly, Nouhra et al. (2005) have observed dense, hydrophobic mats similar to those attributed to Gautieria species beneath the fruit bodies of several Ramaria species. Phylogenetic studies have shown that Gautieria is derived from within the genus Ramaria (Humpert et al. 2001) and these similarities in mat morphologies may be a function of their shared evolutionary history. Overall, these observations imply that the phylogenetic diversity of mat-forming EM species in Douglas-fir forests is likely higher than previously documented and that mat morphology and fruit body associations alone are insufficient to characterize the taxa involved.
Recent advances in molecular methods and the use of DNA sequence databases now allow efficient and accurate identification of fungi from soil (Horton and Bruns 2001; O’Brien et al. 2005). The primary objective of this study was to use sequence data from the nrDNA internal transcribed spacer (ITS) to characterize the richness and encounter rates of EM species that form mats in forests dominated by Douglas-fir. Past watershed-scale studies based both on fruit body collections of EM species (Luoma et al. 1991; Hunt and Trappe 1987) and mat sampling (Griffiths et al. 1996) have shown that the distributions of mat-forming EM fungi can be influenced by stand age. Because of this, a second objective was to compare the mat-forming EM species assemblages in forest stands that represent early and late seral time points typical of Douglas-fir forest succession in the PNW. Characterizing the diversity of mat-forming EM fungi will facilitate sound design of future research into their impacts on soil nutrient cycles and microbial communities.
Materials and methods
Study area
This study was conducted at the HJ Andrews Experimental Forest (HJA), which is part of the National Science Foundation Long-Term Ecological Research (LTER) network. The HJA (lat. 44°15′N, long. 122°10′W) is a 6,400 ha watershed administered as an LTER site on the Willamette National Forest since 1948 (Franklin et al. 1990). About 90% of the area’s annual precipitation falls from October to April with the wettest period in December and peak drought conditions occurring in July. Elevations within the HJA range from 420 to 1,630 m, and the area is typical of the central portion of the western Cascade Mountains in Oregon (Franklin et al. 1990).
Site selection
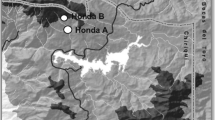
Within the HJA, we sampled 17 old-growth sites (350+ years) and 14 second-growth sites (clear cut in 1950–1960) selected from a larger set of 183 sites previously characterized during an NSF-REU research project on soil characteristics across the HJA (Griffiths, unpublished data). To improve our sampling of the EM mat morphologic diversity previously described from the PNW (Cromack et al. 1979; Griffiths et al. 1991a), we intentionally selected sites characterized as having high EM mat encounter rates (Griffiths, unpublished data). We also sampled one additional second-growth site in the Mill Creek area that lies along the southeast boundary of the HJA. The stands included in this study ranged from 420 to 1,192 m in elevation, had aspects representing all cardinal directions, and ranged from 20% to 35% in average slope (Fig. 1). Because most of our potential study sites were dominated by Douglas-fir, we narrowed the focus of our diversity estimations by sampling only stands dominated by Douglas-fir and western hemlock (Tsuga heterophylla) and avoiding other EM hosts (e.g., Pacific silver fir, Abies amabilis) that typically occur at higher elevations. All stands fall within the western hemlock community types of Franklin (1979) with understory vascular plants including Polystichum munitum/Oxalis oregana in wet sites, Berberis nervosa/Rhododendron macrophyllum in mesic sites, Berberis nervosa/Gautheria shallon in drier sites, and Linnaea borealis in cooler, drier sites.

A relief map showing the locations and relative distribution of old-growth and second-growth plots sampled in this study with an inset showing a schematic of plot sampling methodology. The solid line represents the boarder of the HJ Andrews experimental forest
Site sampling
Collecting was initiated 2 weeks after the first substantial rain (October) of the 2004 and 2005 fall seasons and continued through mid-November of each year. Within each stand a 900 m2 (30 × 30 m) study plot was delineated (Fig. 1). To minimize variability in disturbance due to edge effects, plots were centered on a randomly selected point such that the plot boundary was at least 40 m from any road or other abrupt edge. Substantial patches of fungal mycelia are visible to the naked eye throughout many PNW forest soils, but not all exhibit the rhizomorph densities and morphologies associated with mat formation (Agerer 2001). EM mats as recognized in other studies, and defined here, are dense profusions of rhizomorphs associated with obvious EM root tips that aggregate soil and alter its appearance and are uniform in structure and appearance for an area at least 0.5 m in diameter. This definition characterizes the specific structural characteristics we targeted during field sampling of mat-forming EM fungi. We developed this sampling concept with the guidance of others (Cromack and Caldwell, personal communication) who have substantial experience working on EM mats in the PNW (Cromack et al. 1979, 1988; Griffiths and Caldwell 1992; Griffiths et al. 1990, 1991a, b; Ingham et al. 1991).
During the 2004 field season, all plots were searched for mats by gently removing the upper layers of moss and litter and using a four-pronged rake to explore both organic and mineral soil horizons. Each 900-m2 plot was systematically searched by 2 individuals until 30 mats were located and flagged or the plot had been searched for 30 min, whichever came first. Individual soil explorations (i.e., searches for individual mats) covered areas up to 1 m in diameter and included examination of both organic and mineral horizons up to total depths of 20 cm (depending on depths of individual horizons). Our primary interest was in mat-forming EM fungi that colonize and aggregate forest soils, so large patches of coarse wood (which is often buried) were avoided during sampling.
From the population of flagged mats on each plot, three to five mats were selected randomly for further excavation and closer visual examination. We were primarily interested in characterizing individual mat-forming EM fungi capable of locally dominating forest soils over areas >0.5 m in diameter (see mat definition given above). As a result, mats that lacked obvious ectomycorrhizae, mats <0.5 m diameter or mats composed of several visually distinct rhizomorph and root tip morphologies were excluded from further consideration. When, based on these visual criteria, a mat was eliminated, a new mat was randomly selected as a replacement. Mats that met these visual criteria were sampled by removing a 10-cm3 soil sample from a central location within the mat perimeter. This evaluation and sampling procedure was repeated until three to five mats were selected and sampled for molecular identification of the dominant EM fungus.
All plots were sampled during the 2004 field season and additional, independent mats were sampled from a subset of plots in 2005 to finalize a more complete sampling. In the PNW, many mats formed by EM fungi are believed to be perennial (Griffiths et al. 1991a, 1994) and the persistence of individual mats is likely longer than 2 years (Dunham, personal observation). As a result, data from 2004 and 2005 were pooled for old-growth/second-growth species assemblage comparisons.
Molecular characterization
Using a dissection scope, EM root tip and rhizomorph structures that formed the mat colonizing each soil sample were visually evaluated for uniformity of color, size, texture, and ramification pattern. Entire soil samples were dissected and several EM root tip and rhizomorph structures from the visually dominant EM fungus were removed from several locations throughout the soil core. Selected root tips and rhizomorphs were cleaned by repeated washes with distilled water and approximately one to three EM root tips and three to six rhizomorphs were combined and stored in 300 μl filter sterilized, 2X cetyltrimethylammonium (CTAB) buffer (100 mM Tris–HCl (pH 8.0), 1.4 M NaCl, 20 mM EDTA, 2% CTAB) at −20°C before DNA extraction. Most mats yielded a single collection of EM root tips and rhizomorphs with highly similar (identical) morphologies. When this was not the case, distinct morphologies were prepared separately for DNA extraction.
Genomic DNA was extracted using a modified CTAB method. Tissue was homogenized in 300 μl 2X CTAB and incubated for 1.5 h at 65°C. Following the addition of one volume (300 μl) chloroform, each sample was briefly vortexed then centrifuged for 15 min at room temperature. Finally, DNA was purified from the upper aqueous phase of each sample using the GENECLEAN® extraction kit (Qbiogene, Irvine, CA). DNA samples were diluted 10- to 1,000-fold before use, and the polymerase chain reaction (PCR) was attempted on successive tenfold dilutions until successful amplification of the internally transcribed spacer of the nuclear rDNA repeat unit (ITS) was achieved. The fungal specific primer ITS1-F (Gardes and Bruns 1993) and general primer ITS4 (White et al. 1990) were used in all PCRs. PCR volumes were 30 μl, contained 3 μl of diluted template, and consisted of the following PCR mix components: 1X assay buffer A (Fisher Scientific); 200 μM dNTPs each; 0.2 μM of each primer; 0.75 U Taq DNA polymerase (Fisher Scientific). Cycling conditions included an initial denaturation at 95°C for 3 min followed by 35 PCR cycles (95°C, 45 s; 52°C, 55 s; 72°C, 90 s), and a 10-min 72°C extension applied in a PTC-100 Programmable Thermal Controller (MJ Research).
Before sequencing, PCR products were purified and diluted twofold using QIAquick PCR purification kit (Qiagen, Chatsworth, CA). Following standard direct dye-terminator automated florescence methods (Applied Biosystems), the ITS region was directly sequenced from both primers employed in PCR reactions. Using the program Bioedit (v. 7.0.5; Hall 1999), sequences were edited and grouped manually into alignments of sequences types with 90–100% sequence similarity. When we identified a group of sequences with similarity values that fell within the range where species boundaries were in question (97–99%), we explored the distinctiveness of sequence types within these groups using phylogenetic analyses (see description of Piloderma analysis below). All differentiated sequence types, considered proxies for EM species, were identified to genus or species level by querying the GenBank database using the nucleotide–nucleotide (blastn) BLAST search option available through NCBI (Altschul et al. 1997). Sequences that did not yield conclusive BLAST results in GenBank were similarly compared to a personal database of ITS sequences generated from Ramaria and Hysterangium fruit body collections deposited in the mycological collection of the Oregon State University herbarium (OSC). This sequence database is deposited in GenBank under accession numbers DQ365559–DQ365631. Names were assigned to taxa according to the best BLAST matches. Remaining unidentified sequences were compared to the collection of ITS sequences held by the mycology group in the department of Plant and Environmental Sciences, Göteborg University, Sweden. GenBank accession numbers for sequences that had high similarity to those from EM mats are reported in this study (Table 1).
Data analysis
The composition of EM species assemblages were qualitatively compared using frequency histogram plots. For species richness comparisons, the frequency of mat-forming EM species was defined as the number of plots from which a sequence type (species) was encountered. Plots, rather than individual mats, were treated as the replicated unit because genet sizes have not been characterized for any mat-forming EM species. Consequently, sample independence within plots cannot be assured unless the mats are formed by different taxa. This incidence (presence/absence) data and the program EstimateS (v. 7.5; Colwell 2005) were used to generate expected species accumulation curves (Colwell et al. 2004) and associated 95% confidence intervals for three sample sets: all sites combined, old-growth sites, and second-growth sites. Species accumulation curves (defined as sample-based rarefaction; Gotelli and Colwell 2001) model the expected number of species detected in 1, 2, ... up to the total number of sites. To compare species richness at similar levels of sampling effort, the number of expected species was plotted against the accumulated number of individuals (Gotelli and Colwell 2001).
More detailed characterization of several Piloderma taxa was achieved by phylogenetic analyses including ITS sequences from our EM mats, sequences generated from Piloderma fruit bodies, and highly similar environmental sequences identified in the NCBI sequence database. The program Bioedit (v. 7.0.5; Hall 1999) was used to create the alignment and convert it to the nexus format for analysis in PAUP* (v. 4.0b10; Swofford 2002). The sequence alignment minimized the number of inferred gaps and was recoded from the DNA format to the symbols format to use extended gaps as single informative characters (sensu Bruns et al. 1992). Phylogenetic trees were constructed using the maximum parsimony method implemented in PAUP*. Gaps were treated as a fifth character. Heuristic searches used 100 random taxon addition replicates and TBR branch swapping. Due to dense sampling of terminal clades with many nearly identical sequences (i.e., ITS terminals), finding all most parsimonious trees was not possible; therefore, a maximum of 100 trees of shortest length were retained for each replicate. Relative robustness of nodes was estimated through nonparametric bootstrapping. Settings used were 200 bootstrap replicates with 5 random addition sequences per replicate, TBR branch swapping and MulTrees off.
Results
Richness and encounter frequency of mat-forming EM fungi
Rhizomorph and EM root tip material were collected from 122 mats during the 2004 and 2005 field seasons. Of these samples, 117 (70 from 17 old-growth stands; 47 from 14 second-growth stands) yielded sufficient sequence data for molecular identification of the dominant EM fungus. ITS sequence similarity generally was greater than 99.5% within groups and less than 98% among groups considered unique. Phylogenetic analyses were employed to further explore the distinctiveness of only one variable sequence group (Piloderma, see below). Overall, 28 unique sequence types (EM species) were identified, 24 of which were encountered in old-growth and 13 in second-growth (Fig. 2a and b). Representative sequences for each ITS type were deposited in GenBank under accession numbers DQ365632–DQ365684.

Frequency histograms showing the species composition and relative encounter rates of mat-forming EM fungi in old-growth and second-growth Douglas-fir dominated forests. The names along the x-axes represent the best matches found by BLAST searches of a representative sequence from the ITS type against either personal or published data bases. The qualifier ‘cf.’ is used to denote uncertainty in the specific name due either to equally good matches to identical sequences associated with more than one taxon name or the matches to sequence groups representing multiple taxa deposited under the same name. a A plot of relative species encounter rates across all plots sampled with no information on within plot abundance (i.e., presence/absence only). This figure represents the data used to construct rarefaction curves. b A histogram showing relative species encounter rates across all mats sampled, which includes within plot abundance data
The 28 mat-forming EM species we detected belonged to 10 genera within 6 major homobasidiomycete lineages (Hibbett et al. 1997; Binder and Hibbett 2002; Larsson et al. 2004; Binder et al. 2005) including the Athelioid clade (Piloderma, 4 species), Gomphoid–Phalloid clade(Ramaria, 11 species; Gautieria, 3 species; Gomphus, 1 species; Hysterangium, 2 species), the Euagarics clade (Hebeloma, 1 species; Cortinarius, 3 species), the Trechisporoid clade (Trechispora, 1 species), the Cantharelloid clade (Sistotrema, 1 species), and the Bolete clade (Suillus, 1 species). Unfortunately, many BLAST searches did not yield exact matches to well-described taxa (e.g., Piloderma sequences) and in some cases searches yielded exact matches with two or more identical sequences deposited under different taxon names (e.g., Ramaria sequences). Hence, the specific epithets applied to the mat-forming EM species identified in this study are tentative (Fig. 2a and b) and should be interpreted carefully.
Of the 28 mat-forming EM species, 16 were encountered in only 1 plot (Fig. 2a; 12 in old-growth; 4 in second-growth). The abundance of infrequently encountered species resulted in expected species accumulation curves (Fig. 3) that did not plateau. After using rarefaction to standardize sampling effort (Gotelli and Colwell 2001), consistently fewer species were accumulated across second-growth plots compared to old-growth plots (Fig. 3). Although visually striking, these differences are not statistically significant as the 95% confidence intervals (not shown to improve figure clarity) bracketing the old-growth and second-growth species accumulation estimates overlap substantially at all comparable levels of sampling effort. These broad confidence intervals are likely due to both the large number of species, particularly in old-growth, that were encountered only once and our incomplete sampling of the community of mat-forming EM fungi (Fig. 3). These results do not change when the presence/absence data used to generate expected accumulation curves (data shown in Fig. 2a) is converted to abundance data (data shown in Fig. 2b; accumulation curves not shown).

Expected species accumulation curves for old-growth, second-growth, and all plots combined sample sets. Visual differences in expected species richness between old-growth and second-growth curves are not statistically different. The 95% confidence intervals (not shown for figure clarity) bracketing each curve overlap for all relevant levels of sampling effort. The degree of 95% confidence interval overlap is on the order of 3–4 species for each level of sampling effort
Nine species were shared between old-growth and second-growth habitats leaving 15 species (62.5%) unique to old-growth and 4 (30.8%) unique to second-growth (Fig. 2a and b). In both habitats, the most frequently encountered species was a taxon in the genus Piloderma (Fig. 2a; Table 1; Fig. 4) followed by three Ramaria species in old-growth and two Hysterangium species in second-growth. Mats formed by Piloderma cf. fallax sp. 1 (Fig. 4) were found on 76.5% of old-growth and 84.6% of second-growth plots (Fig. 2a), and constituted 34.3% and 40.4% of the total mats sampled (Fig. 2b) in each respective habitat. In old-growth plots, mats formed by Ramaria species were frequently encountered but the number of associated ITS sequence types was large compared to all other genera detected and no single species dominated. In second-growth, Hysterangium setchellii and H. crassirhachis each formed mats in 53.8% of the plots (Fig. 2a) and represented 17.0% and 19.1% of the respective mats sampled (Fig. 2b).

A maximum parsimony tree generated from ITS sequence generated from EM mats formed by Piloderma species, fruit body collections, and environmental samples deposited in the NCBI GenBank database. All accession numbers preceded by an asterisk are from this study and were collected from stands of pure Douglas-fir and western hemlock. Accession numbers shown in red represent sequences generated from Piloderma fruit bodies
Piloderma diversity
The four distinct but closely related ITS sequence types identified from Piloderma mats in our study area permitted exploration of the species boundaries and diversity within this genus. We aligned 32 mat-associated ITS sequences with 14 Piloderma fruit body sequences and 22 environmental sequences from GenBank. We also included one sequence from a fifth Piloderma species found at the HJA that produces bright yellow rhizomorphs and colonizes coarse wood (Table 1). This sequence set exhibited little length variation but proper alignment required the introduction of gaps 1 to 6 nucleotides long resulting in a total of 545 characters that were included in the maximum parsimony analysis. Six positions were excluded from the analyses. Of the remaining characters, 385 were constant, 71 were variable but parsimony uninformative and 83 were parsimony informative. The analysis yielded 200 trees 238 steps long (CI = 0.8235, RI = 0.9574).
Phylogenetic analysis of this alignment identified five clades with moderate to strong bootstrap support (clades 2–6; Fig. 4) and one unsupported clade (clade 1; Fig. 4). While many sequence types fell into well-supported clades, the relationships among these clades was not well-resolved in this analysis. Sequences associated with the EM mats sampled in this study were included in clades 1–4 (Table 1). Levels of sequence divergence ranged from 0.0% to 0.30% within clades and averaged 5.00% across all in-group comparisons. The lowest between-clade divergence value was 2.3%, (clades 1 and 2; Fig. 4), which is within the range of ITS sequence differentiation observed for biological species EM fungi (e.g., Dunham et al. 2003) and phylogenetic species (e.g., Horton 2002 and references therein). Between-clade divergences ranged up to 8.6% between clades 4 and 5 and 8.4% between clades 3 and 5. Given the magnitude of between-clade sequence divergences, we infer that the well-supported clades identified in this analysis represent Piloderma species or closely related species complexes.
This analysis (Fig. 4) showed that two mat-forming species (clades 1 and 2) were similar to but not conspecific with Piloderma fallax and P. olivaceum. A third clade (clade 3) appears to be conspecific with P. byssinum, while the fourth (clade 4) was allied to but not conspecific with P. byssinum. Clade 4 (Fig. 4) contained a sequence generated from a Piloderma fruit body deposited in the OSC under the name P. fallax highlighting the taxonomic difficulties with this group of fungi. Because fruit bodies sequences were members of only two clades (3 and 4) and one of these (clade 4) contains a sequence from fruit body that perhaps was misidentified, and because we were not able to sample well-characterized type collections for Piloderma taxa, the specific identity of three mat-forming taxa that occur at the HJA remains unclear. Thus, at the HJA experimental forest the genus Piloderma is likely characterized by species that either fruit rarely or have gone uncollected, and may represent undescribed species.
Most of the clades containing mat-forming Piloderma species (except clade 4; Fig. 4) also included environmental sequences deposited in the NCBI GenBank database that come from a diverse set of localities and are associated with several EM host trees (Fig. 4; Table 1). This indicates that at least three of the four Piloderma species detected in this study have broad geographic distributions within North America and/or the Holarctic and show broad EM host specificity within the Pinaceae.
Discussion
The hyphal and rhizomorphic mats formed by some EM fungi perform several ecological functions in PNW forests. Central among these are organic matter decomposition and mineral soil weathering hypothesized to increase nutrient availability to host trees and increase survival of Douglas-fir seedlings under closed canopies (Griffiths et al. 1991a, b, 1994). The presence of mats also creates spatial heterogeneity in the chemical and physical properties of forest soils, which increases the biodiversity of soil-associated organisms (Cromack et al. 1988; Griffiths et al. 1994). This study represents the first application of molecular tools in the identification of mat-forming EM fungi in forests dominated by Douglas-fir.
Richness and encounter frequency of mat-forming EM fungi
The richness of EM species involved in mat formation was much higher than previously documented in PNW Douglas-fir forests (Cromack et al. 1979; Griffiths et al. 1991a, 1996). With the exception of a few mats formed by Hebeloma, Cortinarius, Trechispora, Sistotrema, and Suillus species, the EM species detected in this study belong to genera already known or thought to contain mat-forming species (Griffiths et al. 1996; Agerer 2001; Nouhra et al. 2005). It is interesting to note that we did not encounter any mats formed by EM species in the Thelephoroid clade even though several genera from this clade have been found to contain mat-forming species (Agerer 2001) and EM root tips formed by Thelephoroid species were previously detected within the study area (e.g., Horton et al. 2005).
Expected species accumulation curves (Fig. 3) indicated that our sampling effort was not sufficient to fully document the total richness that exists within this specialized group of fungi. Many belowground studies of EM fungal communities have documented high variability in species frequency of occurrence (Horton and Bruns 2001), which can limit species richness comparisons across treatments. Our study was not an exception as the abundance of infrequently encountered species likely influenced the width 95% confidence intervals around species accumulation estimates making differences between old-growth and second-growth habitats (if they exist) difficult to quantify. ITS sequence divergence values across the entire data set were similar to what we observed within (<0.5%) and between (>2.0%) Piloderma ITS clades (Fig. 4), suggesting that the number of species detected was not artificially inflated by our sequence divergence criteria. In fact, because our sampling was focused primarily on larger (>0.5 m diameter), homogeneous, soil-associated mats that could be visually detected, our species richness estimates likely are conservative. Mat-forming EM species that are restricted to coarse wood comprise another source of diversity that was not sampled in this study. It also is possible that soil-associated EM fungi with darkly pigmented hyphae and rhizomorphs form mats that would easily be missed with the raking methods we employed in our sampling.
These results suggest that within EM genera, the species exhibiting mat soil exploration morphologies (e.g., mat-formers) are more diverse than currently documented (Agerer 2001 and references therein). Several of the new mat-forming taxa identified here are from large, species-diverse genera (e.g., Cortinarius) that exhibit considerable morphological and phylogenetic diversity; therefore encountering new ecological traits (mat-forming) is not unexpected. For some EM genera (e.g., Hysterangium), mat formation likely is a common life-history strategy, but even within these groups not all species form mats. In other EM genera, mat formation is likely an infrequent life-history strategy, thus mat formation by EM genera likely is not an all-or-nothing phenomenon at higher taxonomic levels, i.e., genera. This would partially account for our characterization of this EM mat-forming community as containing many infrequently encountered species and only a few dominant mat-formers. This community structure (few dominants; many infrequently encountered species) is almost universal in belowground studies of EM fungi (Horton and Bruns 2001), thus it was not surprising to find such a pattern in a sampling of a specialized group, such as mat-formers.
In terms of community composition, the mat-forming EM species detected in second-growth may be a subset of those found in old-growth (Fig. 2a and b), suggesting that as with other EM fungi (Dighton and Mason 1985; Kranabetter et al. 2005), the diversity of mat-formers increases as habitat characteristics associated with old-growth develop. Only four EM species from three genera were encountered in more than five plots with the remaining species detected in five or fewer plots (Fig. 2a). One of the four mat-forming Piloderma species was, by far, the most frequently encountered species in both old-growth and second-growth habitats. It is possible that this taxon has not been formally described, but appears to be closely related to P. fallax (Lib.) Stalp. (clade 6; Fig. 4), an organism thought to be old-growth-associated and that colonizes coarse wood with characteristically bright yellow mycelia and rhizomorphs (Smith et al. 2000; Tedersoo et al. 2003). P. fallax (clade 6, Fig. 4) is common on the HJA but did not form any of the soil-associated mats that we sampled. With respect to stand age, the four Piloderma species we found forming mats appear more broadly distributed than P. fallax possibly because they are not associated with coarse wood, which is more common in old-growth forests (Smith et al. 2000).
In second-growth stands, Hysterangium setchellii and H. crassirhachis were encountered with equally high frequency. This was not surprising given that past research on mats has been heavily focused on these species because of their abundance (Cromack et al. 1979, 1988; Griffiths and Caldwell 1992; Griffiths et al. 1994, 1996). Contrary to our expectations, mats formed by Gautieria species constituted only a minor component of the overall richness. Instead, many of the mats exhibiting the powdery, hydrophobic morphology in upper mineral soils thought to be characteristic of Gautieria species (e.g., Griffiths et al. 1995, 1996) were formed by several Ramaria species. In retrospect, given the close phylogenetic relationship between Gautieria and Ramaria (Humpert et al. 2001) commonalities in mat morphologies between these two taxa are consistent with their phylogeny. In total, 11 of the species encountered (39%) belonged to Ramaria and 7 of these were encountered only once. Thus, Ramaria species strongly influenced the high species richness we detected, particularly in old-growth. Of these, R. cf. celerivirescens was the most frequently encountered. The association between increased mat area and encounter frequency for ‘Gautieria-like’ (i.e., Ramaria) mats and old-growth habitats has been quantified previously (Griffiths et al. 1996) and is consistent with the documented strong association between Ramaria species richness and old-growth habitats (US Department of Agriculture and US Department of the Interior 1994; Epsy and Babbitt 1994).
Ectomycorriza formation in Sistotrema and Trechispora
Sistotrema is a polyphyletic genus composed primarily of species that form corticioid fruiting bodies (Larsson et al. 2004; Moncalvo et al. 2006). The genus belongs to the cantharelloid clade where several members are known to be EM (Hibbett et al. 2000). The single species detected in this study is most similar to S. muscicola (Eriksson et al. 1984). EM formation for S. muscicola has recently been demonstrated (Nilsson et al. 2006). Trechispora, another genus with predominantly corticioid members, falls within a homobasidiomycete lineage referred to as the trechisporoid clade (Larsson et al. 2004). To date, EM formation has not been reported for any fungi known from this clade. However, Trechispora species show several traits that usually characterize EM species including occurrence on strongly decayed wood or other debris on the forest floor, loosely attached basidiomata and rhizomorphs, and difficulty germinating spores on common malt agar.
Diversity and dominance of Piloderma
Piloderma species dominated the community of mat-forming fungi in our study area both in terms of the number of plots where they were detected (Fig. 2a) and the high encounter rate of Piloderma mats within plots (Fig. 2b). Our study is not the first to document a number of frequent and probably undescribed Piloderma species in the EM fungal communities associated with coniferous forests (Lilleskov et al. 2002, 2004; Landeweert et al. 2003; Rosling et al. 2003; Douglas et al. 2005). Results from these recent studies indicate that this genus likely houses many undescribed and little-known species of potential ecological importance. We attempted to better characterize the sequence diversity from our study by expanding the context to include the set of environmental Piloderma sequences currently available in GenBank. The taxonomy of this genus is difficult and the names we apply in this study are provisional and should be interpreted with caution. Our primary goal is to document the diversity and interesting biogeographic distribution demonstrated in these data.
We detected five Piloderma species (clades 1–4 and 6; Fig. 4) within our study plots on the HJA, four of which formed mats (clades 1–4). The total diversity of Piloderma in Douglas-fir forests may be even higher as some species have been shown to colonize deeper soil horizons not frequently sampled in EM community studies (Landeweert et al. 2003; Rosling et al. 2003). When compared to fruit body sequences, one environmental sequence (not mat-associated) from the HJA (clade 6; DQ371933) is associated with P. fallax and two of our mat-forming clades appear closely related to P. fallax (clades 1 and 2). However, no fruit body sequences fall into clades 1 and 2 so they remain identified to genus only. Several P. byssinum fruit body sequences were embedded within clade 3 indicating that the EM mats falling within this clade were formed by this species. Clade 4 comprised two mat-associated sequences and one fruit body sequence (DQ371931; Table 1) that was deposited in OSC under P. fallax. This fruit body perhaps was misidentified and exemplifies the taxonomic difficulty associated with fruit body identification for this genus. Given that the ITS sequences within clades 3 and 4 are strongly differentiated from each other (3.4% sequence divergence), clade 4 likely represents a distinct Piloderma species closely related to P. byssinum.
Comparison of the Piloderma sequences from recent studies (Table 1) with those we detected at the HJA demonstrates that several species (e.g., clades 1, 3, 5, and 6) have holarctic distributions, whereas others (clades 2 and 4) are at a minimum broadly distributed in the PNW (Table 1). Our results also indicate that the distribution of Piloderma is likely not restricted by EM host tree specificities. The recent studies from which we drew sequence data documented EM associations with up to nine host tree species that, with one exception, were conifers (Table 1). Because the aim of past research has primarily been to characterize various aspects of EM fungal community structure using root tip data, the soil colonization patterns of individual fungi were not recorded. Consequently, we do not know if the Piloderma species detected on the HJA form mats in the diverse set of forest types they inhabit (Table 1).
The dominance of Piloderma species detected in this study is comparable to the characterization of this genus in many forest ecosystems (Erland and Taylor 1999; Table 1). Given the high encounter frequency of Piloderma mats on the HJA, the species in this genus potentially play a critical role in nutrient availability to Douglas-fir and other EM host trees. Lilleskov et al. (2002) found that Piloderma species dominated low nitrogen (N) soils along an N-gradient and declined in frequency as atmospheric N inputs increased. Kårén (1997) found similar results for P. croceum in a fertilized stand of Scotch pine (Pinus sylvestris) in northern Sweden. In addition, soils colonized by the hyphae of Piloderma croceum on Norway spruce seedlings have been shown to have lower \( {\text{NH}}^{ + }_{4} \) concentrations than immediately surrounding soils (Arocena et al. 2004). Piloderma species are typically associated with late seral forests (Visser 1995; Smith et al. 2000, 2002; Horton et al. 2005) with low availability of readily accessible N (Van Cleve and Vierech 1981). While the actual functions of these mat-forming EM fungi are unknown, in vitro studies of Piloderma species have shown the ability to use protein (Hutchinson 1990; Eaton and Ayres 2002) and the presence of N-acetylhexosaminidase genes (Lindahl and Taylor 2004), suggesting the capacity to use two of the major forms of organic N that occur in mature forests, protein, and chitin. Nutrient uptake studies (Mahmood et al. 2003) and direct measurements of element concentrations within rhizomorphs (Hagerberg et al. 2005) suggest that Piloderma species may also play a role in potassium and calcium transport to EM host trees. Taken together, these observations imply that Piloderma species may be involved in N transport to EM host trees. Our results show that at least some species are also important components of young Douglas-fir forests, indicating a possible role for this genus in forest succession (Cline et al. 2005; Smith et al. 2005). Given their broad geographic distribution, diverse EM host tree associations and dominance, species in this genus are likely critical components of forest ecosystems and warrant further study.
References
Agerer R (2001) Exploration types of ectomycorrhizae: a proposal to classify ectomycorrhizal mycelial systems according to their patterns of differentiation and putative ecological importance. Mycorrhiza 11:107–114
Altschul SF, Madden TL, Schaffer AA et al (1997) Gapped BLAST and PSI-BLAST—a new generation of protein database search programs. Nucleic Acids Res 25:3389–3402
Arocena JM, Göttlein A, Raidl S (2004) Spatial changes of soil solution and mineral composition in the rhizosphere of Norway-spruce seedlings colonized by Piloderma croceum. Journal of Plant Nutrition and Soil Science 167:479–486
Binder M, Hibbett DS (2002) Higher-level phylogenetic relationships of Homobasidiomycetes (mushroom forming fungi) inferred from four rDNA regions. Mol Phylogenet Evol 22:76–90
Binder M, Hibbett DS, Larsson K-H et al (2005) The phylogenetic distribution of resupinate forms across the major clades of mushroom forming fungi (Homobasidiomycetes). Syst Biodiv 3:113–157
Blum JD, Klaue A, Nezat CA et al (2002) Mycorrhizal weathering of apatite as an important calcium source in base-poor forest ecosystems. Nature 417:729–731
Bruns TD, Vilgalys R, Barns SM et al (1992) Evolutionary relationships within the fungi: analyses of nuclear small subunit rRNA sequences. Mol Phylogenet Evol 1:231–241
Castellano MA (1988) The taxonomy of the genus Hysteragium (Basidiomycotina, Hysterangiaceae) with notes on its ecology. Ph.D. thesis, Oregon State University
Cline ET, Ammirati JF, Edmonds RL (2005) Does proximity to mature trees influence ectomycorrhizal fungus communities of Douglas-fir seedlings? New Phytol 166:993–1009
Colwell RK (2005) EstimateS: Statistical estimation of species richness and shared species from samples. Version 7.5. User’s Guide and application published at: http://purl.oclc.org/estimates
Colwell RK, Mao CX, Chang J (2004) Interpolating, extrapolating, and comparing incidence-based species accumulation curves. Ecology 85:2717–2727
Cromack K Jr, Sollins P, Graustein WC et al (1979) Calcium oxalate accumulation and soil weathering in mats of the ectomycorrhizal fungus Hysterangium crassum (Tul. and Tul.) Fischer. Soil Biol Biochem 11:463–468
Cromack K Jr, Fichter BL, Moldenke AM, Entry JA, Ingham ER (1988) Interactions between soil animals and ectomycorrhizal fungal mats. Agric Ecosyst Environ 24:161–168
Dighton J, Mason PA (1985) Mycorrhizal dynamics during forest tree development. In: Moore D, Casselton LA, Wood DA, Frankland JC (eds) Developmental ecology of higher fungi. Cambridge University Press, Cambridge, Great Britain, pp 117–139
Douglas RB, Parker VT, Cullings KW (2005) Belowground ectomycorrhizal community structure of mature lodgepole pine and mixed conifer stands in Yellowstone National Park. For Ecol Manag 208:303–317
Dunham SM, O’Dell TE, Molina R (2003) Analysis of nrDNA sequences and microsatellite allele frequencies reveals a cryptic chanterelle species Cantharellus cascadensis sp. nov. from the American Pacific Northwest. Mycol Res 107:1163–1177
Eaton GK, Ayres MP (2002) Plasticity and constraint in growth and protein mineralization of ectomycorrhizal fungi under stimulated nitrogen deposition. Mycologia 94:921–932
Epsy M, Babbitt B (1994) Record of decision for amendments to forest service and bureau of land management planning documents within the range of the northern spotted owl. US Department of Agriculture, Forest Service and US Department of the Interior, Bureau of Land Management, Washington, DC
Eriksson J, Hjortstam K, Ryvarden L (1984) The Corticiaceae of North Europe, vol 7. Fungiflora, Oslo, Norway
Erland S, Taylor AFS (1999) Resupinate ectomycorrhizal fungal genera. In: Cairney JWG, Chambers SM (eds) Ectomycorrhizal fungi. Key genera in profile. Springer, Berlin, pp 347–358
Franklin JF (1979) Vegetation of the Douglas-fir region. In: Heilman PE, Anderson HW, Baumgartner DM (eds) Forest soils of the Douglas-fir region. Washington State University, Cooperative Extension, Pullman, Washington, pp 93–112
Franklin JF, Bledsoe CS, Callahan JT (1990) Contributions of the long-term ecological research program. Bioscience 40:509–523
Gardes M, Bruns TD (1993) ITS primers with enhanced specificity for basidiomycetes—applications to the identification of mycorrhizae and rusts. Mol Ecol 2:113–118
Gotelli NJ, Colwell RK (2001) Quantifying biodiversity: procedures and pitfalls in the measurement and comparison of species richness. Ecol Lett 4:379–391
Griffiths RP, Caldwell BA (1992) Mycorrhizal mat communities in forest soils. In: Read DJ, Lewis DH, Fitter AH, Alexander IJ (eds) Mycorrhizas in ecosystems. Cambridge University Press, Cambridge, pp 98–105
Griffiths RP, Caldwell BA, Cromack K Jr, Morita RY (1990) Douglas-fir forest soils colonized by ectomycorrhizal mats. I. Seasonal variation in nitrogen chemistry and nitrogen cycle transformation rates. Can J For Res 20:211–218
Griffiths RP, Castellano MA, Caldwell BA (1991a) Hyphal mats formed by two ectomycorrhizal fungi and their association with Douglas-fir seedlings: a case study. Plant Soil 134:255–259
Griffiths RP, Caldwell BA, Ingham ER, Castellano MA, Cromack K Jr (1991b) Comparison of microbial activity in ectomycorrhizal mat communities in Oregon and California. Biol Fertil Soils 11:196–202
Griffiths RP, Baham JE, Caldwell BA (1994) Soil solution chemistry of ectomycorrhizal mats in forest soil. Soil Biol Biochem 26:331–337
Griffiths RP, Chadwick AC, Robatzek M, Schauer K, Schaffroth K (1995) Associations of ectomycorrhizal mats with Pacific yew and other understory trees in coniferous forests. Plant Soil 173:343–347
Griffiths RP, Bradshaw GA, Marks B, Lienkaemper GW (1996) Spatial distribution of ectomycorrhizal mats in coniferous forests of the Pacific Northwest, USA. Plant Soil 180:147–158
Hagerberg D, Pallon J, Wallander H (2005) The elemental content in the mycelium of the ectomycorrhizal fungus Piloderma sp. during colonization of hardened wood ash. Mycorrhiza 15:387–392
Hall TA (1999) Bioedit: a user friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp Ser 41:95–98
Hibbett DS, Pine EM, Langer E, Langer G, Donoghue MJ (1997) Evolution of gilled mushrooms and puffballs inferred from ribosomal DNA sequences. Proc Natl Acad Sci USA 94:12002–12006
Hibbett DS, Gilbert L-B, Donoghue MJ (2000) Evolutionary instability of ectomycorrhizal symbioses in basidiomycetes. Nature 407:506–508
Hintikka V, Naykki O (1967) Notes on the effects of the fungus Hydnellum ferrugineum (Fr.) Karst. on forest soil and vegetation. Commun Inst For Fenn 62:1–22
Horton TR (2002) Molecular approaches to ectomycorrhizal diversity studies: variation in ITS at a local scale. Plant Soil 244:29–39
Horton TR, Bruns TD (2001) The molecular revolution in ectomycorrhizal ecology: peeking into the black box. Mol Ecol 10:1855–1871
Horton TR, Molina R, Hood K (2005) Douglas-fir ectomycorrhizae in 40 and 400 year-old stands: mycobiont availability to late successional western hemlock. Mycorrhiza 15:393–403
Humpert AJ, Muench EL, Giachini AJ, Castellano MA, Spatafora JW (2001) Molecular phylogenetics of Ramaria and related genera: evidence from nuclear large subunit and mitochondrial small subunit rDNA sequences. Mycologia 93:465–477
Hunt GA, Trappe JM (1987) Seasonal hypogeous sporocarp production in a western Oregon Douglas-fir stand. Can J Bot 65:438–445
Hutchinson LJ (1990) Studies on the systematics of ectomycorrhizal fungi in axenic culture. II. The enzymatic degradation of selected carbon and nitrogen compounds. Can J Bot 68:1522–1530
Ingham ER, Griffiths RP, Cromack K, Entry JA (1991) Comparison of direct versus fumigation-flush microbial biomass estimates from ectomycorrhizal mat and non-mat soils. Soil Biol Biochem 23:465–472
Kårén O (1997) Effects of air pollution and forest regeneration methods on the community structure of ectomycorrhizal fungi. Ph.D. thesis, Swedish University of Agricultural Sciences
Kranabetter JM, Friesen J, Gamiet S, Kroeger P (2005) Ectomycorrhizal mushroom distribution by stand age in western hemlock–lodgepole pine forests of northwestern British Columbia. Can J For Res 35:1527–1539
Landeweert R, Leeflang P, Kuyper TW et al (2003) Molecular identification of ectomycorrhizal mycelium in soil horizons. Appl Environ Microbiol 69:327–333
Larsson K-H, Larsson E, Kõljalg U (2004) High phylogenetic diversity among corticioid homobasidiomycetes. Mycol Res 108:983–1002
Lilleskov EA, Fahey TJ, Horton TR, Lovett GM (2002) Belowground ectomycorrhizal fungal community change over a nitrogen deposition gradient in Alaska. Ecology 83:104–115
Lilleskov EA, Bruns TD, Horton TR, Taylor DL, Grogan P (2004) Detection of forest stand-level spatial structure in ectomycorrhizal communities. FEMS Microbiol Ecol 49:319–332
Lindahl BD, Taylor AFS (2004) Occurrence of N-acetylhexosaminidase-encoding genes in ectomycorrhizal basidiomycetes. New Phytol 164:193–199
Luoma DL, Frenkel RE, Trappe JM (1991) Fruiting of hypogeous fungi in Oregon Douglas-fir forests: seasonal and habitat variation. Mycologia 83:335–353
Mahmood S, Finlay RD, Fransson A-M, Wallander H (2003) Effects of hardened wood ash on microbial activity, plant growth and nutrient uptake by ectomycorrhizal spruce seedlings. FEMS Microbiol Ecol 43:121–131
Moncalvo J-M, Nilsson RH, Koster B et al (2006) The cantharelloid clade: dealing with incongruent gene trees and phylogenetic reconstruction methods. Mycologia 98:937–948
Näsholm T, Ekbald A, Nordin A, Giesler R, Högberg M, Högberg P (1998) Boreal plants take up organic nitrogen. Nature 392:914–916
Nilsson RH, Larsson K-H, Larsson E, Kõljalg U (2006) Fruiting body-guided molecular identification of root-tip mantle mycelia provides strong indications of ectomycorrhizal associations in two species of Sistotrema (Basidiomycota). Mycol Res 110:1426–1432
Nouhra ER, Horton TR, Cazares E, Castellano M (2005) Morphological and molecular characterization of selected Ramaria mycorrhizae. Mycorrhiza 15:55–59
O’Brien HE, Parrent JL, Jackson JA, Moncalvo JM, Vilgalys R (2005) Fungal community analysis by large-scale sequencing of environmental samples. Appl Environ Microbiol 71:5544–5550
Perez-Moreno J, Read DJ (2000) Mobilization and transfer of nutrients from litter to tree seedlings via the vegetative mycelium of ectomycorrhizal plants. New Phytol 145:301–309
Rosling A, Landeweert R, Lindahl B et al (2003) Vertical distribution of ectomycorrhizal fungal taxa in a podzol soil profile. New Phytol 159:775–783
Schubert R, Raidl S, Funk R et al (2003) Quantitative detection of agar-cultivated and rhizotron-grown Piloderma croceum Erikss. & Hjortst. by ITS1-based fluorescent PCR. Mycorrhiza 13:159–165
Smith SE, Read DJ (1997) Mycorrhizal symbiosis, 2nd edn. Academic, New York
Smith JE, Molina R, Huso MMP, Larsen MJ (2000) Occurrence of Piloderma fallax in young, rotation-age, and old-growth stands of Douglas-fir (Pseudotsuga menziesii) in the Cascade Range of Oregon, USA. Can J Bot 78:995–1001
Smith JE, Molina R, Huso MMP et al (2002) Species richness, abundance, and composition of hypogeous and epigeous ectomycorrhizal fungal sporocarps in young, rotation-age, and old-growth stands of Douglas fir (Pseudotsuga menziesii) in the Cascade Range of Oregon, USA. Can J Bot 80:186–204
Smith JE, McKay D, Brenner G, McIvers J, Spatafora JW (2005) Early impacts of forest restoration treatments on the ectomycorrhizal fungal community and fine root biomass in a mixed conifer forest. J Appl Ecol 42:526–535
Swofford DL (2002) PAUP* 4.10b: phylogenetic analysis using parsimony (*and other methods). Sinauer, Sunderland, MA, USA
Tedersoo L, Koljalg U, Hallenberg N, Larsson K-H (2003) Fine scale distribution of ectomycorrhizal fungi and roots across substrate layers including coarse woody debris in a mixed forest. New Phytol 159:153–165
US Department of Agriculture and US Department of the Interior (1994) Final supplemental environmental impact statement on management of habitat for late-successional and old-growth forest related species within the range of the northern spotted owl, vol 1. Forest Service, Bureau of Land Management, Portland, Oregon
Van Cleve K, Vierech LA (1981) Forest succession in relation to nutrient cycling in the boreal forest of Alaska. In: West DC, Shugart HH, Botkin DB (eds) Forest succession: concepts and applications. Springer, New York, NY, pp 185–211
Visser S (1995) Ectomycorrhizal fungal succession in jack pine stands following wildfire. New Phytol 129:389–401
White TJ, Bruns T, Lee S, Taylor J (1990) Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In: Innis MA, Gelfand DH, Sninsky JJ, White TJ (eds) PCR protocols, a guide to methods and applications. Academic, New York, NY, pp 315–322
Acknowledgements
This work is a contribution from the HJ Andrews Ecosystem Research Group. Our plots were selected from a sampling network designed and installed by Bob Griffiths who generously supplied data useful in plot selection. Kentaro Hosaka provided invaluable field assistance and intellectual input during the early stages of this project. Additional field assistance was provided by Katie Tinnesand and Laurel Kluber. Significant laboratory assistance was provided by Tri Tran, Alicia Leytem, and Maribeth Latvis. We thank Conrad Schoch for the assistance with the phylogenetic analyses of Piloderma taxa and Jane Smith and Doni McKay for laboratory resources. This work was funded by a National Science Foundation Microbial Observatory grant MCB-0348689 to David Myrold, Peter Bottomley, Joseph Spatafora, Kermit Cromack Jr., and Bruce Caldwell at Oregon State University. Any opinions, findings, and conclusions or recommendations expressed in this material are those of the authors and do not necessarily reflect the views of the National Science Foundation.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Dunham, S.M., Larsson, KH. & Spatafora, J.W. Species richness and community composition of mat-forming ectomycorrhizal fungi in old- and second-growth Douglas-fir forests of the HJ Andrews Experimental Forest, Oregon, USA. Mycorrhiza 17, 633–645 (2007). https://doi.org/10.1007/s00572-007-0141-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00572-007-0141-6