
Abstract
The Mississippian (Early Carboniferous) is generally a period of scarce carbonate buildups in South China. This study documents outcrops of stromatolite mounds at Mengcun and Helv villages, in Laibin City, Guangxi Province, South China. The stromatolite mounds contain various stromatolite morphologies including laminar, wavy-laminar, domal or hemispheroidal, bulbous, and flabellate-growth columns. Intramound rocks are brachiopod floatstone and dark thin-bedded laminated micrite limestone. Individual stromatolites at Mengcun village are generally 3–6 cm thick and morphologically represent relatively shallow-water laminar (planar and wavy-undulated stromatolites) and deeper-water domal, bulbous and columnar forms. Where mounds were formed, the stromatolites continued growing upward up to 60 cm thick. Thrombolitic fabrics also occur but are not common. Stromatolite microscopic structure shows the bulk of the lamination to consist of wavy microbialite and discrete thin micritic laminae. These mounds are intercalated in deep-water fore-reef talus breccia, packstone formed as a bioclastic debris flow and thin-bedded limestone containing common chert layers of the Tatang Formation (late Viséan). Further evidence supporting the deep-water setting of the stromatolite mounds are: (1) a laterally thinning horizon of brachiopod floatstone containing deep-water, small, thin-shelled brachiopods, peloidal micritic sediments and low-diversity, mixed fauna (e.g., thin-shelled brachiopods, tube-like worms and algae) that have been interpreted as storm deposits, (2) common fore-reef talus breccias, (3) lack of sedimentary structures indicating current action, (4) preservation of lamination with sponge spicules, and (5) lack of bioturbation suggesting that the stromatolites grew in a relatively low energy, deep-water setting. The stromatolite mounds are the first described stromatolite mounds in Mississippian strata of South China and contain evidence that supports interpretations of (1) growth history of Mississippian microbial buildups and (2) environmental controls on stromatolite growth and lithification.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Mississippian (Early Carboniferous) carbonate buildups have been the subject of much attention (e.g., Lane and Ormiston 1982; James 1983; West 1988; Bridges et al. 1995; Webb 2002) because of their palaeoclimatic importance and to the insights they provide into the recovery processes of skeletal reef builders associated with the Frasnian-Famennian and end-Devonian extinction events (Newell 1972; Fisher 1981; James 1983; Sheehan 1985; West 1988; Frakes et al. 1992; Bridges et al. 1995; Caplan and Bustin 1999; Saltzman et al. 2000; Aretz and Chevalier 2007). Only scarce and greatly impoverished reef communities continued to grow during the Tournaisian. The communities consist mainly of calcimicrobes and microbialite (stromatolites and thrombolites). It is only just after the beginning of the early Viséan stage that there was a resurgence of skeletal metazoan in the reef biome (Adams 1984; Dix and James 1987; Bancroft et al. 1988; Webb 1989, 1998, 1999, 2005; Horbury 1992; Mundy 1994; Ahr and Stanton 1994). Mississippian carbonate buildups are known in eastern Australia (Jull 1968; Webb 1989, 1998, 1999; Shen 2002; Shen and Webb 2005, 2008), western Newfoundland of Canada (Dix and James 1987), Europe (Adams 1984; Bancroft et al. 1988; Horbury 1992; Lauwers 1992; Somerville et al. 1992, 1996; Somerville 2003; Pickard 1992, 1996; Mundy 1994; Bridges et al. 1995; Kirkham 2005; Aretz and Chevalier 2007), North Africa (Pareyn 1959; Bourque et al. 1995; Madi et al. 1996); northern Arkansas, New Mexico, Virginia and Texas of USA (Webb 1987; Ahr 1989; Christopher 1990; Ahr and Stanton 1994), Urals of Russia (Antoshkina 1998; Skompski et al. 2001), and Japan (Ota 1968; Haikawa and Ota 1978; Sano and Kanmera 1996; Nakazawa 2001). Relatively deep-water Pennsylvanian microbial-algal mounds described from Cantabrian Mountains in northern Spain (Della Porta et al. 2003) show an analogous depositional setting to those at Laibin.
Mississippian reefs and mounds are poorly documented in China and only one reef was previously described at Langping, Tianlin County, Guangxi (e.g., Fang and Hou 1989). It was described as a bryozoan-coral patch reef (Fang and Hou 1985, 1989) constructed mainly by fasciculate corals and fistuliporoid bryozoans. Coral corallites are generally encrusted by microbialites (Shen 2002). Worldwide, few stromatolite reefs and mounds have been described from Mississippian strata (Bertrand-Sarfati and Monty 1994; Webb 2005), especially from the fore-reef slope settings. This paper presents the first study of the earliest known Mississippian stromatolite mounds in South China (Fig. 1).

Location of the Mississippian stromatolite mounds at Mengcun and Helv villages, Laibin City, Guangxi Province, South China. a Location of Guangxi Province in China, b location of Laibin City in Guangxi Province, c locations of Mengcun and Helv near Laibin City and the Mississippian stromatolite mounds at Mengcun and Helv villiages
The stromatolite mounds reported in the study are of particular interest because they are unique Mississippian stromatolitic buildups in South China; with microbially constructed rigid frameworks in a deep-water fore-reef slope setting; they have a diversity of stromatolite morphologies, which have not been described in other analogous depositional settings. The depositional settings of stromatolite mounds are referred to a fore-reef slope environment based on the following evidence: (1) rare benthic fossils in the mound sequences; (2) lateral facies changes from platform facies with massive limestones containing mega crinoids, Gigantoproductus, and Kueichouphyllum in the southeast to marginal slope and basin facies with manganiferous limestone intercalated with chert layers to the northwest; (3) mounds are intercalated in fore-reef talus breccias, packstone formed by bioclastic debris flow and thin-bedded cherts. Thin-bedded, laminated, dense, uniform, fine-grained laminated micrite limestones intercalated with stromatolite boundstone contain sponge masses and spicules, but lack other fossils and algal/microbial remains, indicating a deep-water and low sedimentary rate and suggesting that it is deposited in generally quiet water, in a deep slope or ramp setting, below wave base (e.g., Madi et al. 1996). The documentation and description of their structure and composition can help us to interpret the growth history of Mississippian reefs and mounds in South China and helps us to understand environmental controls on stromatolite growth and morphology, in general.
Geological setting
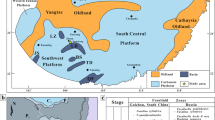
The Mississippian tectonic setting in Guangxi, South China was a continuation of the Devonian tectonic configuration with the presence of a craton in the Upper Yangtze area to the north, a rift basin in the Youjiang region, and a deep-water trough in the Fangcheng-Qingzhou area to the south (Chen et al. 1994) (Fig. 2a). The structural trends were inherited from major fault trends of Devonian age, with NE–SW trending; NW–SE trending rifts forming a series of interconnected troughs (Chen et al. 1994). However, faulting activity was relatively weak in the Mississippian (Chen et al. 1994) (Fig. 2a). Mississippian sedimentation occurred mainly in a passive continental margin basin of the craton, characterized by near shore siliciclastic facies and carbonate platform facies. Carbonate buildups formed locally on the platform margin adjoining the rift basin (intraplatform depression) (Fig. 2a).

Mississippian strata in South China have been extensively studied by Chinese stratigraphers and palaeontologists (Hou et al. 1982; Zhang 1987; Wang et al. 1987, 2006; Wang 1990; Wang and Jin 2000). The Mississippian sedimentary rocks in the Guangxi Province consist of the Fengningian subsystem, which is subdivided into two series, and four stages, namely: the Aikuannian and Tatangian Series; and the Tangbagouan, Jiusian, Shangsian, and Dewuan stages. Stromatolite mounds were developed during the Shangsian Stage and occur in the Luocheng Member of the Tatang Formation in the central part of Guangxi Province (Fig. 2). The Tatang Formation, about 422 m thick, is composed of three members; the Huangjin, the Shimen, and the Luocheng members from bottom to top (Fig. 2). The Huangjin Member, about 260 m thick, is characterized by dark-gray, bedded limestone and calcareous mudstone. The Shimen Member, about 54 m thick, is composed of gray to dark gray, bedded peloidal and bioclastic wackestone, interbedded with thin-bedded argillaceous limestone, with local micrite cemented intraclastic limestone interpreted as storm deposits (Guangxi Bureau of Geology and Mineral Resources 1985). Chert nodules and concretions are common. The Luocheng Member, about 108 m thick, consists of gray to light gray, bedded bioclastic packstone, wackestone, and mudstone, interbedded with micritically cemented intraclastic limestone interpreted as storm deposits (Guangxi Bureau of Geology and Mineral Resources 1985). This member locally contains chert nodules and concretions as well as argillaceous layers. Fossils are dominated by foraminifers, including Bradyina rotula, Cribrospira panderi, and Endothyranopsis crassa. Corals (e.g., Yuanophyllum sp. and Kueichouphyllum sinense) and brachiopods (e.g., Gigantoproductus edelburgensis and Delepinea comoides) are common. Foraminifer, brachiopod and coral data establish a late Viséan age for the Luocheng Member (see Fig. 2).
The study was carried out in two locations, Helv village and Mengcun village (Fig. 2b) that are 10 km apart. Stratigraphic sections throughout the mounds were measured, described and sampled. The stromatolite mound at Helv village occurs on the east side of NW–SE trending fault on the east flank of the Sijiao anticline. The outcrop of the Helv mound is 11–21 m in thickness, 45–60 m in diameter, and has an S–N orientation. The stromatolite mound at Mengcun occurs on the west flank of the Sijiao anticline (Fig. 2b). The Mengcun mound is 100–150 m in diameter with many thick limestone outcrops within farmland. These are arranged into a mound-shaped structure that thins to the north and south. To the south or southwest, the stromatolite mounds pass laterally into the surrounding slope sediments. The sides of the mounds are not very distinct to the north or northeast. The thickness reaches 60 m, which is much thicker than the surrounding slope sediments. In both locations, the mounds were developed in the late Viséan strata of the Loucheng Member. The base of the Helv and Mengcun mounds is developed at approximately the same stratigraphic horizon in the Luocheng Member on the dark gray micrite with chert layers. The stromatolite mound at Helv is also intercalated within the dark gray medium-bedded micrite with chert. The upper part is not preserved due to a fault. At Mengcun, stromatolite mounds show alternations of laminated to stromatolitic limestone and dark gray to black micrite with fine-medium grained (<0.5 mm) bioclasts of the mound facies with occasional intervals of fore-reef talus breccias. Two horizons of thick-bedded or wedge-shaped accumulations of fore-reef talus breccias occur in the lower part of the stromatolite mound at Mengcun and in the upper part of the mound succession (Fig. 3), which forms an accumulation of allochthonous lithoclasts mixed with autochthonous fine-grained matrix at the front of platform margin and upper slope settings. Underlying strata are dark gray to dark thin-bedded limestones of marginal slope facies (Kuang et al. 1999). The talus breccias consist mostly of coarse, poorly sorted lithoclasts with poor roundness in a fine-grained matrix. The lithoclasts are mainly laminated limestones and bioclastic limestones that represent platform facies, but some micritic limestones of deep-water slope occur. The stratigraphic section at Mengcun village was used as the major section (Fig. 3), because of the good exposure and the fact that the base and top of the mound are visible. The Mengcun section was sampled bed by bed, from fore-reef talus breccias below to 2 m above stromatolite mounds, in order to record the vertical succession and to decipher the growth history of stromatolite mounds. Polished vertical slabs and their acetate peels were used to identify rock types, textures, structures, and biotic components of the stromatolite interval. Microtextures and microbial composition of stromatolites were determined from thin sections. In addition, petrographic study focused on the stromatolite interval and the underlying fore-reef talus breccias, the intercalated laminated limestone, brachiopod floatstone and micrite of the intramound facies, and overlying fore-reef talus breccias.

Measured section at Mengcun village showing stacking pattern of the stromatolite mounds, fore-reef talus breccias, and intercalated laminated micrite limestone
Growth morphologies of stromatolites
The semantics of microbially constructed carbonates used in this study is based on terminology proposed by Burne and Moore (1987), Webb (1996) and Riding (1999, 2000). Microbialite is a general term referring to sedimentary structures constructed primarily by microbial communities (Burne and Moore 1987). Stromatolite refers to microbialites with a laminated internal structure (Riding 1999), thrombolite refers to those with a clotted to irregular morphology (Riding 2000) and calcimicrobes are microbial morphologies that are preserved and include recognizable taxa (Webb 1996).
Stromatolites in the Luocheng Member of the Tatang Formation at Mengcun and Helv villages are compound structures that developed from laminar mats, to stromatolites, and then to stromatolite mounds, marking clear differences in scale from micro to meso to macroscopic. The laminae in the stromatolites are of microbial origin because dark-colored, thin microbial filaments (Fig. 4b) are preserved between the laminae. The thickness of the microbial layers varies from 0.25 to 3 mm (Fig. 4b, c), but can be up to 6 mm (Fig. 4c). Thick laminae (0.25–3 mm) are light colored and contain more spar calcite, and thinner laminae (0.10–0.5 mm) are dark colored and contain much more micrite (Fig. 4a). Thick laminae may be laterally linked when calcite-rich laminae were locally thick enough (2–6 cm thick). Later laminae followed the relief of earlier laminae and show a larger-scale (15 cm high, 30 cm long) wavy form (Fig. 4a). Under the microscope, laminae are parallel and slightly undulated (0.25–0.75 mm thick) alternating with dark-colored irregular microbial layers (Fig. 4b). Laminated limestone is also composed of various kinds of wavy laminae including (1) small and low-relief wavy laminae (amplitude less than 5 mm), (2) large and high-relief wavy laminae (amplitude more than 5 mm), (3) symmetric wavy laminae, (4) asymmetric wavy laminae, (5) conical wavy laminae, and (6) domal wavy laminae. Generally, the lower and upper wavy laminae are more or less parallel, but with different amplitudes. In some cases, laminated limestone may pass upward into stromatolite limestone.

Morphologies of outcropping stromatolites and thin-section photomicrographs in plane-polarized light. a Laminar mats, laminae show wavy growth form and become thick in some parts to form domal relief, pen = 14.5 cm, Helv section, b laminae that construct stromatolite, showing alternating layers of microbialite and sediment, microbes with dark, tubular outlines (arrow), occur between microbialite and sediment, Helv section, c laminae of stromatolite, showing thick microbialite layers, microbes (arrow) preserved between microbialites, and the surfaces of microbialites are not smooth; Mengcun section, d domal stromatolites show regular mats, which grew on limestone containing fine bioclasts, Mengcun section
Various forms of stromatolites occur in the Mengcun and Helv sections, including laminar to wavy-laminar (e.g., Buick et al. 1981; George 1999), domal or hemispheroidal, bulbous, and regular flabellate columns. On outcrops the microbial laminae appear as a laminated limestone; showing thin (2–5 mm) dense (10 layers/20 cm) laminae on weathered surfaces. Laminated limestones generally occur below stromatolites in the lower part of the section (Fig. 3) and contributed to the initial stabilization of sediments, or are intercalated within stromatolite mounds. The surface (Fig. 4a, arrow) between earlier and later laminae is distinct and wavy and marks a transition to darker laminae.
Domal forms occur where wavy laminae grew into asymmetric and domal accumulations (Buick et al. 1981) and are common in the Mengcun mound (Fig. 4d). Individual stromatolites are 10–40 cm in diameter and 5–6 cm high with low relief (0.5–1 cm) of laminae. The upper surfaces are domal and the edges are smooth or irregular if stromatolites are separated as branches (Figs. 6b, 8d). Relief of the stromatolite above the substrate ranges from 0 to 6 cm. The surface is smooth or slightly crenulated. After the microbial laminae stabilized, the laminae progressed from wavy forms to laterally linked, to mammillary structures. By growing upward and laterally, some individual stromatolites coalesce and become laterally linked. At Mengcun and Helv sections, many stromatolites were separated in the earlier stage and linked in the later stage.
Hemispheroidal forms are where concentric laminae grew on a plane along a same axial line with the radius increasing from internal to external laminae (Gebelein 1976). Hemispheroidal stromatolites are generally separated, but can be locally linked at very early growth stage. The sizes of the hemispheroidal stromatolites are variable, and in most cases they are 4.5–7 cm in diameter and 3.5–5 cm high (Fig. 5a). Microbial laminae grew convex-upwardly on the substrates, showing concentric arrangement of laminae with increasing diameters from the inner part to the outer part. Hemispheroidal stromatolites are not stacked vertically but are overlain by wavy laminated limestone, and in turn by well-bedded non-stromatolitic sediments. In other words, the overall structure of these stromatolites is a combination of unstacked and laterally linked hemispheroids of variable diameters.

Various stromatolite morphologies: a Laterally linked hemispheroidal stromatolites, which are interbedded in wavy laminae mats; some laminae, especially the top laminae and overgrown laminae of domed stromatolite, are silicified, pen = 14 cm, Helv section, b spheroid or bulbous stromatolites, generally 2–3 cm in diameter, pen = 15 cm, c cross-section of the stromatolites, showing laminations in a concentric form, and the later growth inherits the form of the early laminae (arrow point to stratigraphic top), d stromatolites show regular flabellate-growth columns, pen = 15 cm. b, c, d Mengcun section
Bulbous forms are stromatolite with lower eccentric growth centers and concentric laminae. Generally the lower part is small and upper part is larger and it resembles an inverse cone (e.g., Southgate 1989). Bulbous stromatolites (Fig. 5b) look like cauliflowers and are composed of eccentric laminae or discontinuous laminae surrounding a nucleus that is eccentric in the lower part. The laminae may be irregular, wavy, or crinkly in shape because of the uneven concentration of the mats and the uneven distribution of sediment across the mat. The sizes of bulbous stromatolites are 2–3 cm in diameter and 2–5 cm in height. Bulbous stromatolites may grow on top of one another and accumulate upward and laterally to form large stromatolite colonies, 50–60 cm in height and 40–50 cm in diameter (Fig. 5b). In plain view, these stromatolites are circular, irregular, or elliptical, with irregular, concentric and crinkly laminae (Fig. 5c).
The term ‘flabellate column’ is defined in this paper as where laminae grew into a column with changeable or unchangeable radius, and later diverged into several columns. These columns are generally built up into flabellate columnar stromatolites and are the most common microbial structures in the Mengcun and Helv mounds. These stromatolites display regular, flabellate-expanded overlapping growth patterns in longitudinal section (Fig. 5d) and expanded at 60°–80° angles between two edges. Individual stromatolites look like inverted cones with domal upper surface and round edges. The laminae grew upward in a columnar pattern. The convex-upward laminae developed from pseudo-columnar forms, which can be laterally linked to hemispherical structures. In the later stage, stromatolite columns may branch into more columns that gradually thickened upward. The initial growth forms of flabellate-columnar stromatolites developed into relatively small spheroid structures (1.5–2 cm in diameter) or were stabilized directly on the lime mud. Later, laminae growing on spheroid structures developed into a typical flabellate stromatolite. No evidence of hardground or disconformity surface was identified. When these stromatolites were growing on lime mud, they generally developed into an incomplete or half-flabellate form (Fig. 6a). Flabellate stromatolites are generally laterally linked and grew continuously upward to form large stromatolite mounds (Fig. 6b). In some cases, sediments separate fan-like stromatolite columns during their growth, but they were still stacked to form thick stromatolite limestone. Continuous growth forms are generally 12 cm high (Fig. 5d), and discontinuous growth forms range from 5 to 6 cm high but are stacked (Fig. 6a). Discontinuous growth forms did not grow directly on top of one another but on the lime mud substrate intercalated within stromatolites (Fig. 6a).

Various stromatolite morphologies: a gently convex-growth columnar, showing incomplete or half flabellate form, b continuing upward growth flabellate columnar, distinct, regular and relatively thick cements were interlaminated with microbial laminae. c and d Thin-section photomicrographs in plane-polarized light. c Thrombolitic fabric, some intraclasts are encrusted by thrombolitic fabric, d thrombolite containing intraclasts is in the lower part and foraminifers (arrow) cemented by sparry calcite are in the upper part. a–d Mengcun section
Various morphologic stromatolites stacked together to form high relief mounds. These compound stromatolites are generally 1–30 m in diameter and 4–17 m thick, and form discrete mounds with a total stacking thickness reaching 60 m (Fig. 3). They initially contained horizontal (laminar and tabular) stromatolites, later becoming domal, equidimensional to asymmetric structures (bulbous forms), and finally formed flabellate columnar stromatolites. The stacking pattern of the stromatolite mounds, fore-reef talus breccias, intercalated laminated micrite limestone, and thin-shelled brachiopod floatstone is shown in Fig. 3. The ratio of stromatolite boundstone to non-boundstone facies thickness in the mound facies is 70:30%.
Microtextures of the thrombolites/stromatolites
Thrombolitic fabrics are more easily recognizable in thin-sections but may also be recognized on outcrop. They are characterized by massive or irregularly clotted fabrics (Fig. 6c) with intraclasts, coarse skeletal debris and some void space. Intraclasts are generally rounded, variable in size, unevenly distributed, and are coated. In some cases, the thrombolitic fabrics were overgrown by microbial laminae on which foraminifer grains were cemented by sparry calcite (Fig. 6d). These foraminifer grains may have been washed in from neighboring platform areas.
Under the microscope, stromatolites generally show a variety of forms, including slender columnar, irregular microscale columns, regular bulbous and digitate morphologies. Slim columnar forms developed on hard substrates and are parallel or integrated in late growth stages. Cryptic interstices were formed between the slender stromatolite columns or within the dichotomous stromatolite. Some cryptic microbes and microbial deposits occur in these interstices (Fig. 7a).

Thin-section photomicrographs and in plane-polarized. a Stromatolitic fabric, columnar stromatolites show upward growth form and parallel divergents, some cryptic microbes (arrow) grew within the space between stromatolites, b stromatolites, laminae are indistinct due to recrystallization and stromatolite growth is discrete, c two separated stromatolite columns, showing mushroom-like outline and divergent growth form, white-colored spars are needle-like arrays of cement fans (arrow), d digitate stromatolites are in the lower part and cross-sections of digitate stromatolites are in the upper part, some problematic microbes can be seen in the cross-section of stromatolites. a–d Mengcun section
Irregular microscale columns (Fig. 7b) are related to discontinuous growth of stromatolites. The top is crenulated, and irregular. Interstices between discrete microscale columns contain micrite. The stromatolite tubers with indistinct laminae were probably affected by recrystallization (Fig. 7b). Sparitic laminae have a rather constant thickness and are traceable for several millimeters. Perpendicular sparitic protuberances of single laminae are common.
Regular bulbous forms (Fig. 7c) are mushroom-like in shape. They appear like an inverted taper, with small diameter in early growth increasing in the later stages with well-developed and convex-upward laminae. Stromatolitic laminae are composed of relatively thick, dark-colored layers (0.2–0.5 mm) alternating with thin, light-colored parts (0.02 mm). The former contains indistinct successive needle-like arrays of cement fans growing up, probably a replacement fabric of earlier aragonite cements. The spaces between neighboring stromatolites are filled with calcite spar and dark-colored peloids (Fig. 7c).
Digitate forms (Fig. 7d) are tightly packed with an upward-pointing inverted fingertip shape 6–8 mm in diameter. Laminae may not be recognizable due to recrystallization. The tops and edges of the stromatolites are generally domed and smooth where they are close together, but may be uneven when they are isolated. Layers of microbialite, about 1.5–2 mm thick, overgrew the fingertip-shaped stromatolites that have eroded tops. In plain view, stromatolites are circular and ellipsoid, showing radial fibrous calcite textures (Fig. 7d) that are considered to reflect recrystallization (e.g., Bertrand-Sarfati and Monty 1994). In the limestone containing digitate stromatolites, microbial fabrics are 1.5–6 mm thick and consist of dark tiny bush-like structures (Fig. 8a), similar to dendrolite. Sub-spherical and elliptical growth microbes, probably Ortonella, show fan-shaped radial calcite fabric and the 45°–60° branching typical of Ortonella (Fig. 8c). The tops and sides of algal filament fabrics are encrusted by microbial laminae that thicken to form low-relief stromatolites (Fig. 8c). Locally stromatolites may have divergent tops, showing irregular, serrated and short branches (Fig. 8d) that are the result of organic boring. The interstices between branches or interstices between stromatolites are filled with dark-colored microbial sediments (Fig. 8d), containing tangled tubular microbes (probably Girvanella) and fine peloids, indicating microbial sediment on stromatolites.

Thin-section photomicrographs in plane-polarized light. a Cross-section of stromatolites are in the lower part and microbial matter is in the upper part, b enlarged from part a showing some problematic microbes (arrow) in cross-sections of stromatolites, c sub-spherical and elliptical growth microbe a shows fan-expanded radial calcite fabric that is encrusted by microbial laminae (arrow), d stromatolites showing serrated tops that are overgrown by microbes and microbial carbonates. a–d Mengcun section
Some unidentified microbes occur within stromatolites (Fig. 8b) and in the interstices between stromatolites. One of the problematic microbes is hollow and shows spherical cross-sections. Internal structure is slightly radiating fibrous calcite. The wall of the microbes is relatively thick (Fig. 8b). Tube-like worm communities occur in the brachiopod limestone. Worm tubes are hollowed and dumbell-shaped with expanded ends, generally 18 mm in length and 2–3 mm in diameter. The hollow ends of worm tubes are filled with light-colored calcite. The wall of the worm tubes is relatively thick (Fig. 12a) and differs from Terebella sp. of Della Porta et al. (2003).
Other related depositional features
Fore-reef talus breccias are gray in color and are composed of stromatolite and laminated lithologies. They form accumulations of allochthonous lithoclasts mixed with a matrix at the front edge of the platform margin and upper slope settings. Underlying strata are dark gray to dark thin-bedded limestones of marginal slope facies (Kuang et al. 1999). The talus breccias consist mostly of coarse and poorly sorted lithoclasts with poor roundness, set in a fine-grained matrix. The lithoclasts are mainly laminated limestones and bioclastic limestones that represent platform facies, but some micritic limestone of deeper-water origin also occurs. Talus breccias appear to be predominantly composed of platform facies and mound facies that overlie the deep-water, thin-bedded micrite with chert layers of the slope and basin facies, or they are directly associated with stromatolite mound facies. Breccias have clasts ranging from 7 to 16 cm in diameter (Fig. 9). Most of them are angular or sub-angular, and others are sub-rounded. Large ones are lath-shaped and prismatic, ~1.5 m in length, and have a disorganized fabric. Fore-reef talus breccias occur below the mound and are interbedded with the mound facies where they are composed of complex clasts including micrite, dolomite, and cherts, reflecting typical clast-supported breccias of fore-reef facies. To the north of the Mengcun mound, the breccia passes laterally into laminate-stromatolite mound with dark gray micrite containing chert layers along the bedding.

Fore-reef talus breccia of the Mengcun stromatolite mound (pen = 14.5 cm), lower part of the Mengcun section
Generally, metazoan fossils are not common in the stromatolite mound successions in the Laibin area. Small-sized foraminifers, fragments of crinoids, thin-shelled brachiopods and trilobite bioclasts have been observed in the brachiopod floatstone and laminated micrite limestone (Fig. 3) of the intramound facies. Some sponge masses and spicules occur in dark colored, thin-bedded laminated micrite limestone. Problematic red algae are abundant locally, such as in the brachiopod floatstone within the middle part of stromatolite mound. They are assigned to Ungdarella and occur as unsorted bioclasts (Fig. 10a). Other problematic algae occur together with Ungdarella and have some characteristics similar to those of Koninckopora (e.g., in Fig. 10). However, these algae do not belong to Koninckopora according to Prof. Riding (personal communication). They show thin and hollowed sticks like Amphipora with a thin dark wall and central canaliculus. They are dense and have well-spaced dissepiments that developed between the wall and canaliculus (Fig. 10b). These problematic algae generally occur as fragments and bioclasts and are overgrown by marine cements or are encrusted by cement crusts to form oncoid-like and stromatolite-like fabrics (Fig. 10c). Oncoids, with irregular shapes and different sizes (Fig. 10d), and irregular intraclasts, with laminated crusts (Fig. 10d), also occur with Ungdarella, problematic algae and small and thin-shelled brachiopods. No further skeletal micro/macrofossils have been recorded in the stromatolite boundstone lithology.

Thin-section photomicrographs in plane-polarized light. a Problematic red alga (Ungdarella sp.) and problematic alga similar to Koninckopora (arrow). b Fragments and bioclasts of problematic algae, showing transverse section and longitudinal section. c Problematic algae (arrow) are overgrown by marine cements or encrusted by cement crusts to form oncoid-like and stromatolite-like fabrics. d Oncoids with irregualar shapes and different sized (arrow) and irregular intraclasts with laminated pendent cement crusts. a–d Mengcun section
A noticeable bed, ~60 cm thick, containing small and thin-shelled brachiopods (Fig. 11) occurs as a lens in the middle part of the stromatolite mound succession. Brachiopods are 7–10 mm in diameter. The general shape with one valve overlapping the other shows a resemblance to brachiopods, but the lack of an obvious hinge shows affinities to ostracodes (Fig. 12b). However, the valves are characterized by internal cardinal structures as in brachiopods. The section cut through a shell shows the texture of spine bases (Fig. 12c) and also exhibits a crinkly surface that may represent growth bands. This brachiopod floatstone is interbedded within the stromatolite boundstone (Fig. 3), but it has not been found in Helv mound. Cement crusts, coat small brachiopods to form isopachous rinds (Fig. 12b, c), show concentric bands with radiaxial fibrous fabric. Thickness of the cement crusts varies from 0.2 to 5 mm. Most of bioclasts are also coated by cement crusts and microbial envelopes. Coated grains and oncoid-like forms are unevenly distributed in the rock. The interstices within loosely packed coated grains and oncoid-like forms are filled with sparry calcite (Fig. 12d).

Brachiopod floatstone occur in the Mengcun mound (see Fig. 3) (lens cover = 5.9 cm)

Thin-section photomicrographs in plane-polarized light, Mengcun section. a Tube-like worm forms (arrow) in the central part of the photograph. b Oncoid floatstone, small brachiopods are encrusted by crinkly and convolute cement crusts and formed oncoid-like forms bearing cores of brachiopods. c The section cutting shell surface shows spine base remains, the outside of the cross-section exhibits a crinkly surface that may represent growth bands. d Skeletal grains, oncoid-like form, algal and foraminifer grains and microbialites are encrusted by microbial micrite that is cemented by sparry calcite
Distinct and relatively thick cement deposits were interlaminated with microbial laminae (Fig. 6b) showing regular or irregular layers. Thick cement deposits are regular with constant thickness, but microbial laminae are irregular with variable thickness. These cements have a marine origin because they are the first cement phase, are overgrown by microbial crusts, are non-luminescent, with bright luminescent patches, and do not occur as generations of cements or replace earlier fabrics.
Discussion
Since the end of the 1960s, stromatolite morphology has been interpreted as having been controlled by hydraulic energy and sedimentation rate (Schenk 1967; Horodyski 1977; James and Bourque 1992; Hofman 1994). Stromatolites with high synoptic relief commonly occur in deeper-water settings, whereas flat-laminated mats are typically restricted to shallow-water environments (Logan et al. 1974; Hoffman 1974; Peryt 1975).
A wide variety of stromatolite morphologies exist in the Mengcun and Helv mounds including laminar, wavy-laminar, domal or hemispheroidal, bulbous, and flabellate-growth columnar forms. Stromatolite morphologies change in vertical sequences, from wavy to planar laminar stromatolite morphologies to domal or hemispheroidal, to bulbous, and to flabellate columnar forms, which may indicate a cumulative effect of changing environmental conditions from deep subtidal and proximal fore-reef slope to a deep-water, lower slope setting through time. The stromatolite mounds at Mengcun and Helv appear to have developed in a probable deep subtidal to proximal fore-reef slope setting as these stromatolite mounds were directly developed on deep-water, thin-bedded micrite with chert layers (Fig. 13).

Sketch of the depositional facies for the Mississippian stromatolite mounds at Mengcun and Helv, Laibin City, Guangxi Province, South China
Laminations with alternations of darker, organic-rich micritic laminae and coarser, microsparitic increments may characterize the shallow part of deep, lower- to moderate-energy subtidal areas of the open platform setting (Beukes and Lowe 1989). In Mengcun and Helv sections, laminated forms developed into planar and wavy-undulated stromatolites. Planar stromatolites with typical relief of less than 3–5 cm alternate with wavy-undulated stromatolitic structures, indicating a deep subtidal, transitional platform margin to proximal fore-reef slope setting (Fig. 13). Domal stromatolites, which are discrete at the early growth stage but merge at the later stage to form continuous laminae covering several domal stromatolites, indicate increasing of water energy and decreasing of sedimentation rate, suggesting a proximal fore-reef slope setting (e.g., Beukes and Lowe 1989; Denver and Kaesler 1992; Gòmez-Pérez 2003) (Fig. 13). Bulbous forms are considered to form below normal wave base in a deep-water slope setting with moderate circulation and low-energy (Fig. 13). This interpretation is based on the fact that sediments between bulbous stromatolites are mostly micrite and do not contain shallow water benthic fossils. The analogs such as the bulbous stromatolites in the Viséan Strathclyde Group, eastern Midland Valley of Scotland (Guirdham et al. 2003) are believed to be precipitated in situ by microbial mediation, rather than by trapping and binding of allochthonous material, although the latter occurs in non-marine shallow water lacustrine environments under hypersaline conditions (Guirdham et al. 2003). The columnar forms in the Mengcun and Helv mounds are interpreted to relate to bioherms. The stromatolites consisting of flabellate-growth columns may indicate a deep-water fore-reef slope setting (Fig. 13) as these stromatolites are generally intercalated with the dark gray, thin-bedded micrite containing chert layers but rare benthic fossils.
Calcareous algae are common in packstone of bioclastic debris flow origin and brachiopod floatstone. Ungdarella is most abundant, but some algae are problematic. Ungdarella was erected by Maslov (1956) and the genus ranges from Mississippian (upper Viséan) to the Upper Permian (Guadalupian) (Toomey and Johnson 1968). However, it typically occurs in the Moscovian/mid-Pennsylvanian (Chuvashov and Anfimov 2007). It is widespread geographically, but it is a problematic fossil that has most commonly been placed in the red algae. Calcareous algae are important bathymetric indicators and have been used to delineate three bathymetric zones for a Mississippian (upper Viséan) ramp of western Algeria based on light penetration (Madi et al. 1996). Ungdarella was thought to occur in the dysphotic zone, where there is little ambient light, and which is characterized by the presence of red algae and absence of green algae. On that basis, the Mengcun mound most probably formed in a dysphotic zone of the proximal fore-reef slope setting. However, Ungdarella algae occurs in growth position in upper Viséan microbial reefs in Australia that were interpreted to be shallow-water buildups in near shore settings (see Pl. 30, Fig. 4 in Webb 1999). Comparably, the Helv mound lacks calcareous algae and shallow-water metazoans and may represent a deeper, aphotic zone of the slope.
The importance of the stromatolite mounds described in this paper is that the stromatolite boundstones, which are intercalated with fore-reef talus breccias, packstone formed by bioclastic debris flow, and thin-bedded laminated micrite limestone of the late Viséan Tatang Formation, constitute the first known examples of Mississippian deep-water stromatolites in South China. They are also characterized by rare benthic fossils, no burrows, no ooids, or aggregates, and scarce packstones. The Mengcun mound was developed in a proximal fore-reef slope setting, whereas the Helv mound grew on a slightly more distal, deeper-water slope setting. The former presents various stromatolite types, thick mound sequence packstone formed by bioclastic debris flow, and especially fore-mound talus breccias. However, the latter is intercalated with dark thin-bedded micritic limestones with common chert layers. The Helv mound is thinner (about 7 m) and shows a change from wavy laminar, hemispheroidal, to flabellate-growth columnar stromatolites. No bioclastic packstones and fore-mound talus breccias have been found there.
The described stromatolites (domal, bulbous and columnar) are more or less morphologically or genetically analogous to other deep-water stromatolites reported from the Precambrian to the Pleistocene (e.g., Hoffman 1974; George 1999; Brachert 1999; Petrov and Semikhatov 2001; Gómez-Pérez 2003). Deep-water stromatolites occur in the Lower Proterozoic platform-to-basin facies in (1) the Great Slave Lake in Canada (Hoffman 1974); (2) in the massive Mesoproterozoic stromatolitic reef complex from the Burovaya Formation in Siberia (Petrov and Semikhatov 2001); (3) in the Devonian reef complexes of the Canning Basin, northwestern Australia (George 1999); (4) in the Early Jurassic deep-water stromatolitic bioherms related to possible methane seepage in the Los Molles Formation, Neuquen, Argentina (Gòmez-Pérez 2003); and (5) in the Pleistocene deep sea (Last Glacial maximum), Red Sea (Brachert 1999). Like other Phanerozoic deep-water stromatolites, the Mississippian examples at Mengcun and Helv villages grew from a deep subtidal and proximal fore-reef slope to a deep-water, lower slope setting.
Pennsylvanian deep-water microbial-algal mounds have been described from the Cantabrian Mountains in northern Spain (Della Porta et al. 2003). These mounds are more or less similar to the Laibin stromatolite mounds in development in a slope setting, association with breccias, content of red algae (Ungdarella sp.), and abundant cement. However, the Laibin mounds are characterized by stromatolites and low-diversity fauna, which differ from microbial-algal mounds from the Cantabrian Mountains. Mississippian (Upper Viséan) thrombolitic-Ortonella reefs of the Clifton Down Limestone, SW England are associated with well-laminated stromatolites, and contain calcispheres, foraminifers and ostracodes. Such benthic assemblages and their muddy carbonate matrices suggest a similar environment to that of Laibin mounds with a relatively low-energy, deep subtidal, marine depositional environment (Kirkham 2005). However, burrow structures and oolitic sediments in the former indicate a high energy; shallow-water setting that is different from the deep-water slope setting of Laibin mounds. Stromatolite-rich reefs that occur in the shallow-water lower Tournaisian Gudman Formation of eastern Australia contain a diverse flora and fauna and occur in even higher-energy settings within active intertidal ooid shoals (Webb 2005).
Mississippian stromatolite mounds are uncommon worldwide. Bertrand-Sarfati (1994) described stromatolite domes in the Mississippian of the Ajjers Basin, Eastern Sahara, Algeria. These stromatolites formed in a mixed siliciclastic-carbonate depositional setting that is quite different from the carbonate depositional setting of the Laibin stromatolite mounds in South China. Also, stromatolite morphologies in the Ajjers Basin are dominated by larger domes (Bertrand-Sarfati 1994), whereas the Laibin stromatolites show several, but mostly columnar, morphologies. Antoshkina (1998) described stromatolite bioherms and biostromes from the large late Viséan to Serpukhovian shelf edge reefs in the subpolar Urals. There, stromatolite boundstones are characterized by laminated fabrics associated with calcified microbes and phylloid algae. The stromatolites that dominate these mounds are similar to those in the Laibin stromatolite mound having horizontally and concentrically laminated fabrics. Viséan algal-brachiopod-coral bioherms from Bol’shaya Nadota carbonate complex, Subpolar Urals (Skompski et al. 2001) differ from the Laibin stromatolite mounds in having an associated shallow-water benthic fauna, different reef builders, and high-energy water condition (e.g., interfingering with organodetrital-oolitic facies). Stromatolite biostromes are also described from the Pennsylvanian of northeast Kansas (Sawin et al. 1985). They were formed in an intertidal to slightly deep subtidal environment associated with a coastal embayment that is different from the fore-reef slope setting of the Laibin stromatolite mounds. Also, the stromatolite morphologies of Northeast Kansas differ from those in the Laibin mounds as they are dominated by inverted cones and abundant oncolites.
One interval of low-diversity fauna in the Mississippian stromatolite mounds at Mengcun, small, thin-shelled brachiopods locally associated with clusters of tube-like worm communities and may indicate deeper, quieter water conditions (Gao et al. 2005; He et al. 2005). Also, some microbes, including those in the cryptic interstices of stromatolites, such as tangled tubular (probably Girvanella) and unidentified microbes within stromatolites, may represent a deep, low-energy, and slightly dysphotic, cryptic setting with normal marine salinity (Elicki 1999). Stromatolites at Mengcun and Helv villages are cement-rich, consisting of micritic laminae alternating with cements. Cement crusts are well developed around thin-shell brachiopods and calcareous algae; they are a radial fibrous fabric or isopachous fibrous fabric with growth bands, indicating submarine origin. They may have been microbially mediated to some degree because some layers of isopachous fibrous cements are overgrown by dark-colored microbial laminae. Such concentric crusts consisting of radiaxial fibrous marine cements are well developed at the outer margin and on the proximal slope of Triassic carbonate platforms in Italy (Russo et al. 2000). This is consistent with the suggested proximal fore-reef slope setting for Mengcun mound. Tube-like worms, cement crusts and thin-shelled brachiopods are present in many Carboniferous mounds, especially in those of Mississippian (latest Viséan) seep deposits on the top of the drowned Middle to Late Devonian Iberg atoll reef, in the Harz Mountains, Germany (Peckmann et al. 2001) and Mississippian low-temperature hydrothermal vent communities from Newfoundland, Canada (von Bitter et al. 1990). These examples have limestone with low-diversity but high-abundance faunas of brachiopods and well-developed cement crusts. The brachiopods are generally articulated, and they are closely associated with hydrocarbon-derived carbonates. Submarine seeps might promote radial fibrous or isopachous fibrous cement growth around brachiopods and algae. However, whether they are seep-related deposits or not could not be determined because other defining criteria for recognizing cold water seep and vent communities are not present. This one bed of brachiopod floatstone within the Mengcun mound, which contains a mixed shallower and deeper-water fauna such as algae and small, thin-shelled brachiopods and it thins out laterally, may represent a storm deposit or possibly represent an interval with upwelling of dysoxic waters from deeper in the basin.
Conclusions
-
(1)
Mississippian stromatolite mounds at Mengcun and Helv villages occur typically in a deep-water succession consisting of fore-reef talus breccias, packstone formed by bioclastic debris flow, thin-bedded laminated limestone, and cherty micrites. Other features such as the lack of phototrophic organisms in the stromatolite boundstone, lack of bioturbation, no ooids, or aggregates, and preservation of lamination with sponge spicules also indicate a deep-water, low energy marine setting.
-
(2)
Stromatolites are compound structures that developed from laminar mats, to stromatolites, and then to stromatolite mounds. Various morphologies of stromatolites occur in the Mengcun and Helv mounds in repeated successions that comprise, laminar to wavy-laminar, domal or hemispheroidal, bulbous, and regular flabellate columns. These may indicate a cumulative effect of changing environmental conditions through time.
-
(3)
Various stromatolite morphologies are stacked together with initially horizontal laminar and tabular growth forms later becoming domal and bulbous forms, and finally flabellate columnar stromatolites to form mounds. The stromatolites are generally 10–30 m in diameter and 4–17 m thick, and form discrete mounds with a total stacking thickness reaching 60 m.
-
(4)
The significance of the stromatolite mounds described in this paper is that the stromatolite boundstones constitutes the first known examples of Mississippian deep-water stromatolites in South China. The stromatolites are more or less morphologically or genetically analogous to other deep-water stromatolites from Precambrian to Pleistocene. Unlike many Paleozoic deep-water stromatolites formed in a relatively narrow subtidal zone, these stromatolites apparently grew contemporaneously across the deep subtidal and proximal fore-reef slope to deeper-water, lower slope settings.
-
(5)
Problematic microbes are embedded in peloidal micrite associated with the stromatolitic laminae, suggesting a microbial origin for the stromatolites. Well-developed oncoid-like and stromatolite-like cement crusts around brachiopods and algae and mixed fauna consisting of deep-water, small, thin-shelled brachiopods and shallower water calcareous algae may suggest storm deposits within the stromatolite mound of proximal fore-reef slope setting.
References
Adams AE (1984) Development of algal-foraminiferal-coral reefs in the Lower Carboniferous of Furness, northwest England. Lethaia 17:233–249. doi:10.1111/j.1502-3931.1984.tb01623.x
Ahr WM (1989) Mississippian reef facies in the southwest: a spectrum of variations in depositional style and reservoir characteristics. In: Flis JE, Price RC, Sarg JF (eds) Search for the subtle trap––hydrocarbon exploration in mature basins. West Texas geological society, symposium publication, pp 1–19
Ahr WM, Stanton RJ Jr (1994) Comparative sedimentology and palaeontology of Waulsortian mounds and coeval level-bottom sediments of the lower lake valley formation (Lower Mississippian) in the Sacramento mountains (New Mexico, USA). Abh Geologischen Bundesanstalt 50:11–24
Antoshkina AI (1998) Organic buildups and reefs on the Palaeozoic carbonate platform margin, Pechora Urals, Russia. Sediment Geol 118:87–211. doi:10.1016/S0037-0738(98)00012-8
Aretz M, Chevalier E (2007) After the collapse of stromatoporoid-coral reefs––the Famennian and Dinantian reefs of Belgium: much more than Waulsortian mounds. In: Alvaro JJ, Aretz M, Boulvain F, Munnecke A, Vachard D, Vennin E (eds) Palaeozoic reefs and bioaccumulations: climatic and evolutionary controls geological society of London special publication, vol 275, pp 163–188
Bancroft AJ, Somerville ID, Strank AE (1988) A bryozoan buildup from the Lower Carboniferous of North Wales. Lethaia 21:1–65. doi:10.1111/j.1502-3931.1988.tb01753.x
Bertrand-Sarfati J, Monty C (1994) Phanerozoic stromatolites II. Kluwer, Dordrecht, pp 1–471
Bertrand-Sarfati JB (1994) Siliciclastic carbonate stromatolite domes, in the Early Carboniferous of the Ajjers basin (Eastern Sahara, Algeria). In: Bertrand-Sarfati J, Monty C (eds) Phanerozoic stromatolites II. Kluwer, Amsterdam, pp 71–100
Beukes NJ, Lowe DR (1989) Environmental control on diverse stromatolite morphologies in the 3000 Myr Pongola supergroup, South Africa. Sedimentology 36:383–397. doi:10.1111/j.1365-3091.1989.tb00615.x
Bourque PA, Madi A, Mamet BL (1995) Waulsortian-type bioherm development and response to sea-level fluctuations: upper Viséan of Béchar basin, Western Algeria. J Sediment Res B65:80–95
Brachert TC (1999) Non-skeletal carbonate production and stromatolite growth within a Pleistocene deep ocean (Last Glacial maximum, Red Sea). Facies 40:211–228. doi:10.1007/BF02537475
Bridges PH, Gutteridge P, Pickard NAH (1995) The environmental setting of Early Carboniferous mud-mounds. In: Monty CLV, Bosence DWJ, Bridges PH, Pratt BR (eds) Carbonate mud-mounds: their origin and evolution. International association of sedimentologists, special publication, vol 23, pp 171–190
Buick R, Dunlop JSR, Groves DI (1981) Stromatolite recognition in ancient rocks: an appraisal of irregularly laminated structures in an early Archean chert-barite unit from North-Pole, Western-Australia. Alcheringa 5:161–181. doi:10.1080/03115518108566999
Burne RV, Moore LS (1987) Microbialites: organosedimentary deposits of benthic microbial communities. Palaios 2:241–254. doi:10.2307/3514674
Caplan ML, Bustin RM (1999) Devonian-Carboniferous Hangenberg mass extinction event, widespread organic-rich mudrock and anoxia: causes and consequences. Palaeogeogr Palaeoclimatol Palaeoecol 148:187–207. doi:10.1016/S0031-0182(98)00218-1
Chen HM, Wu XH, Zhang Y, Li YX, Wen QY (1994) Carboniferous lithofacies, palaeogeography and mineralization in South China. Geological Publishing House, Beijing (in Chinese)
Christopher CC (1990) Late Mississippian Girvanella-bryozoan mud mounds in southern West Virginia. Palaios 5:460–471. doi:10.2307/3514838
Chuvashov BL, Anfimov AL (2007) On the taxonomy of the paleozoic red algae of the family Ungdarellaceae. Paleontol J 41:95–102. doi:10.1134/S0031030107010108
Della Porta G, Kenter JAM, Bahamonde JR, Immenhauser A, Villa E (2003) Microbial boundstone dominated carbonate slope (upper Carboniferous, N Spain): microfacies, lithofacies distribution and stratal geometry. Facies 49:175–208
Denver LE II, Kaesler RL (1992) Paleoenvironmental significance of stromatolites in the Americus Limestone Member (Lower Permian, midcontinent, USA). Kansas University, Paleontological contributions. N Series, vol 1, pp 1–11
Dix GR, James JP (1987) Late Mississippian bryozoan/microbial build-ups on a drowned karst terrain, Port Au Port Peninsula, western Newfoundland. Sedimentology 34:779–793. doi:10.1111/j.1365-3091.1987.tb00802.x
Elicki O (1999) Palaeoecological significance of calcimicrobial communities during ramp evolution: an example from the lower Cambrian of Germany. Facies 41:27–39. doi:10.1007/BF02537458
Fang SX, Hou FH (1985) Bryozoan-coral patch reef of Da-Tang age of Carboniferous period of Langping area, Tianlin County, Guangxi Province. J Southwest Petrol Inst 23:1–15 (in Chinese)
Fang SX, Hou FH (1989) Bryozoan-coral patch reefs of the Carboniferous carbonate platform, Langping area, Tianling County, Guangxi, China. In: XI Congress International de Stratigraphie et de Geologie du Carbonifere, Beijing, 1987, vol 4, pp 19–25
Fisher AG (1981) Climatic oscillations in the biosphere. In: Nitecki MH (ed) Biotic crises in ecological and evolutionary time. Academic Press, New York, pp 103–131
Frakes LA, Francis JE, Syktus JI (1992) Climate modes of the Phanerozoic, Cambridge University Press, Cambridge, 274 pp
Gao YQ, Yang FQ, Peng YQ (2005) Characteristics of late Permian deep-water sedimentary environments: a case study of Shaiwa Section, Ziyun County, Guizhou Province, Southwestern China. J China Univ Geosci 1:12–24
Gebelein CD (1976) Open marine subtidal and intertidal stromatolites (Florida, the Bahamas and Bermuda). In: Walter MR (ed) Stromatolites. Developments in sedimentology. Elsevier, New York, pp 381–388
George AD (1999) Deep-water stromatolites, Canning Basin, northwestern Australia. Palaios 14:493–505. doi:10.2307/3515399
Gòmez-Pérez I (2003) An Early Jurassic deep-water stromatolitic bioherm related to possible methane seepage (los Molles Formation, Neuquen, Argentina). Palaeogeogr Palaeoclimatol Palaeoecol 201:21–49. doi:10.1016/S0031-0182(03)00508-X
Guirdham C, Andrews JE, Browne MAE, Dean MT (2003) Stratigraphic and palaeoenvironmental significance of microbial carbonates in the Asbian Sandy Craig formation of fife. Scott J Geol 39:151–168
Guangxi Bureau of Geology and Mineral Resources (1985) Regional geology of Guangxi Zhuang autonomous region. Geological Publishing House, Beijing (in Chinese)
Haikawa T, Ota M (1978) A Lower Carboniferous coral reef found in the Nagatophyllum satoi zone of the Akiyoshi Limestone Group, southwest Japan. Bull Akiyoshi-dai Mus Nat Hist 13:1–13
He WH, Shen SZ, Feng QL, Gu SZ (2005) A late Changhsingian (Late Permian) deep water brachiopod fauna from the Talung Formation at the Dongpan section, Southern Guangxi, South China. J Paleontol 79:927–938. doi:10.1666/0022-3360(2005)079[0927:ALCLPD]2.0.CO;2
Hoffman P (1974) Shallow and deep-water stromatolites in lower Proterozoic platform-to-basin facies change, Great Slave Lake, Canada. Am Assoc Pet Geol Bull 58:856–867
Hofman HJ (1994) Quantitative stromatolitology. J Paleontol 68:704–709
Horbury AD (1992) A Late Dinantian peloid cementstone-palaeoberesellid buildup from Lancashire, England. Sediment Geol 79:107–137. doi:10.1016/0037-0738(92)90007-E
Horodyski RJ (1977) Environmental influences on columnar stromatolite branching patterns: examples from the middle Proterozoic belt supergroup, Glacier National Park, Montana. J Paleontol 51:661–671
Hou HF, Wu ZJ, Wu XH, Yang SP (1982) The carboniferous system of China (Stratigraphy of China, No.1). Geological Publishing House, Beijing, pp 187–218 (in Chinese)
James NP (1983) Reef envrironment. In: Scholle PA, Bebout DG, Moore CH (eds) Carbonate depositional environments. American association of petroleum geologists, Memoir, vol 33, pp 346–440
James NP, Bourque PA (1992) Reefs and mounds. In: Walker RG, James NP (eds) Facies models: response to sea level change, Chap 17. Geological association of Canada
Jull PK (1968) The Lower Carboniferous limestones in the Monto-Old Cannindah distinct: a brief description and a proposed name. J Qld Gov Min 69:199–201
Kirkham A (2005) Thrombolitic-Ortonella reefs and their bacterial diagenesis, upper Viséan Clifton down limestone, Bristol area, SW England. Proc Geol Assoc 116:221–234
Kuang GD, Li JX, Zhong K, Su YB, Tao YB (1999) Carboniferous of Guangxi, Stratigraphy of Guangxi, China Part 2. China University of Geosciences Press, Wuhan, pp 1–258 (in Chinese)
Lane HR, Ormiston A-R (1982) Waulsortian facies, Sacramento mountains, New Mexico: guide for an international field seminar, 2–6 March, 1982. In: Bolton K, Lane HR, Le Mone DV (eds) Symposium on the paleoenvironmental setting and distribution of the Waulsortian Facies. El Paso, Texas, El Paso geological society and University of Texas at El Paso, pp 115–182
Lauwers AS (1992) Growth and diagenesis of cryptalgal-bryozoan buildup within a middle Viséan (Dinantian) cyclic sequence, Belgium. Ann Soc Geologique Belg 115:187–213
Logan BW, Hoffman P, Gebelein CD (1974) Algal mats, cryptalgal fabrics, and structures, Hamelin Pool, Western Australia. American association of petroleum geologists. Memoir 22:140–194
Madi A, Bourque P-A, Mamet BL (1996) Depth-related ecological zonation of a Carboniferous carbonate ramp: upper Viséan of Béchar basin, western Algeria. Facies 35:59–80. doi:10.1007/BF02536957
Maslov VP (1956) Iskopayemyye izvestkovye vodorosli SSSR (Fossil calcareous algae of the USSR). Akademiya Nauk SSSR. Tr Geologicheskogo Inst 160:1–301
Mundy DJC (1994) Microbialite-sponge-bryozoan-coral framestones in lower Carboniferous (late Viséan) buildups of northern England (UK). Canadian society of petroleum geologists. Memoir 17:713–729
Nakazawa T (2001) Carboniferous reef succession of the Panthalassan open-ocean setting: example from Omi Limestone, Central Japan. Facies 44:183–210. doi:10.1007/BF02668174
Newell ND (1972) The evolution of reefs. Sci Am 226:54–65
Ota M (1968) The Akiyoshi Limestone Group: a geosynclinal organic reef complex. Bull Akiyoshi-dai Sci Mus Nat Hist 5:1–44
Pareyn C (1959) Les récifs carbonifères du Grand Erg occidental. Bull Soc Geologique Fr Ser 71:347–364
Peckmann J, Gischler E, Oschmann W, Reitner J (2001) An Early Carboniferous seep community and hydrocarbon-derived carbonates from the Harz Mountains, Germany. Geology 29:271–274. doi:10.1130/0091-7613(2001)029<0271:AECSCA>2.0.CO;2
Peryt TM (1975) Significance of stromatolites for the environmental interpretation of the Bundsandstein (Lower Triassic) rocks. Geol Rundsch 64:143–158. doi:10.1007/BF01820659
Petrov PY, Semikhatov MA (2001) Sequence organization and growth patterns of late Mesoproterozoic stromatolite reefs: an example from the Burovaya Formation, Turukhansk Uplift, Siberia. Precambrian Res 111:257–281. doi:10.1016/S0301-9268(01)00163-2
Pickard NAH (1992) Depositional controls on lower Carboniferous microbial buildups, eastern Midland Valley of Scotland. Sedimentology 39:1081–1100. doi:10.1111/j.1365-3091.1992.tb01998.x
Pickard NAH (1996) Evidence for microbial influence on the development of lower Carboniferous buildups. In: Strogen P, Somerville ID, Jones GLI (eds) Recent advances in lower Carboniferous geology. Geological society, London, special publications, vol 107, pp 371–385
Riding R (1999) The term stromatolite: towards an essential definition. Lethaia 32:321–330
Riding R (2000) Microbial carbonates: the geological record of calcified bacterial-algal mats and biofilms. Sedimentology 47(Suppl 1):179–214. doi:10.1046/j.1365-3091.2000.00003.x
Russo F, Mastandrea AC, Stefani M, Neri CF (2000) Carbonate facies dominated by syndepositional cements: a key composition of middle Triassic platforms. The Marmolada case history (Dolomites, Italy). Facies 42:211–226. doi:10.1007/BF02562573
Saltzman MR, González lA, Lohmann KC (2000) Earliest Carboniferous cooling step triggered by the Antler orogeny? Geology 28:347–350. doi:10.1130/0091-7613(2000)28<347:ECCSTB>2.0.CO;2
Sano H, Kanmera K (1996) Microbial controls on Panthalassan Carboniferous––Permian ocean build-ups, Japan. Facies 34:239–255. doi:10.1007/BF02546167
Sawin RS, West RR, Twiss PC (1985) Stromatolite biostromes in the upper Carboniferous of Northeast Kansas. In: Dutro JT Jr, Pfeffekorn HW (eds) Neuvième Congrès International de Stratigraphie et de Geologie du Carbonifère Compte Rendu 5, paleontology, paleoecology, paleogeography. Southern Illinois University Press, Carbonadale and Edwardsville, Illinois, pp 361–372
Schenk PE (1967) The significance of algal stromatolites to paleoenvironmental and chronostratigraphic interpretations of the Windsorian stage (Mississippian), Maritime provinces. Special paper geological association of Canada, pp 229–243
Sheehan PM (1985) Reefs are not so different––they follow the evolution-any pattern of the level-bottom communities. Geology 13:23–33. doi:10.1130/0091-7613(1985)13<46:RANSDF>2.0.CO;2
Shen JW (2002) Effects of differing tectono-stratigraphic settings on late Devonian and Early Carboniferous reefs, Western Australia, eastern Australia, South China, and Japan. Unpublished Ph.D. thesis, University of Queensland, Brisbane
Shen JW, Webb GE (2005) Metazoan-microbial framework fabrics in a Mississippian (Carboniferous) coral-sponge-microbial reef, Monto, Queensland, Australia. Sediment Geol 178:113–133. doi:10.1016/j.sedgeo.2005.03.011
Shen JW, Webb GE (2008) The role of microbes in reef-building communities of the Cannindah limestone (Mississippian), Monto region, Queensland, Australia. Facies 54:89–105. doi:10.1007/s10347-007-0116-2
Skompski S, Paskowski M, Krobicki M, Kokovin K, Korn D, Tomas A, Wrzolek T (2001) Depositional setting of the Devonian/Carboniferous biohermal Bol’shaya Nadota carbonate complex, Subpolar Urals. Acta Geol Pol 51/3:217–235
Somerville ID (2003) Review of Irish lower Carboniferous (Mississippian) mud-mounds: depositional setting, biota, facies and evolution. In: Ahr W, Harris AP, Morgan WA, Somerville ID (eds) Permo-Carboniferous carbonate platforms and reefs. Society for economic paleontologists and mineralogists, special publication 78 and American association of petroleum geologists Memoir 83: 239–252
Somerville ID, Pickard NAH, Strogen P, Jones GLI (1992) Early to mid-Viséan shallow water platform buildups, north Co. Dublin, Ireland. Geol J 27:151–172. doi:10.1002/gj.3350270205
Somerville ID, Strogen P, Jones GLI, Somerville HEA (1996) Late Viséan buildups of the Kingscourt Outlier, Ireland: Possible precursors for Upper Carboniferous bioherms. In: Strogen P, Somerville ID, Jones GLI (eds) Recent advances in lower Carboniferous geology. Geological society, London, special publications, vol 107, pp 127–144
Southgate P (1989) Relationships between cyclicity and stromatolite form in the late Proterozoic Bitter Springs Formation, Australia. Sedimentology 36:323–339. doi:10.1111/j.1365-3091.1989.tb00610.x
Toomey DF, Johnson JH (1968) Ungdarella americana, a new red alga from the Pennsylvanian of southeastern New Mexico. J Paleontol 42:556–560
Von Bitter PM, Scott SD, Schenk PE (1990) Early Carboniferous low-temperature hydrothermal vent communities from Newfoundland. Nature 344:145–148. doi:10.1038/344145a0
Wang XD, Jin YG (2000) An outline of Carboniferous Chronostratigraphy. J Stratigr 24:90–98 (in Chinese)
Wang XD, Wang X, Zhang F, Zhang H (2006) Diversity patterns in Carboniferous and Permian rugose corals in South China. Geol J 41(3–4):329–343. doi:10.1002/gj.1041
Wang ZH (1990) Conodont zonation of the lower Carboniferous in South China and phylogeny of some important species. Courier Forschungsintitut Senckenberg 130:41–46
Wang ZH, Lane HR, Manger WL (1987) Conodont sequence across the mid-Carboniferous boundary in China and its correlation with England and North American. In: Wang CY (ed) Carboniferous boundary in China. Science Press, Beijing, pp 89–106
Webb GE (1987) Late Mississippian thrombolite bioherms from the Pitkin formation of Northern Arkansas. Bull Geol Soc Am 99:686–698. doi:10.1130/0016-7606(1987)99<686:LMTBFT>2.0.CO;2
Webb GE (1989) Late Viséan coral-algal bioherms from the Lion Creek formation of Queensland, Australia. In: XI Congress International de Stratigraphie et de Geologie du Carbonifere, Beijing, vol 3, pp 282–295
Webb GE (1996) Was Phanerozoic reef history controlled by the distribution of non-enzymatically secreted reef carbonates (microbial carbonate and biologically induced cement)? Sedimentology 43:947–971. doi:10.1111/j.1365-3091.1996.tb01513.x
Webb GE (1998) Earliest known Carboniferous shallow-water reefs, Gudman Formation (Tn 1 b), Queensland, Australia: implications for late Devonian reef collapse and recovery. Geology 26:951–954. doi:10.1130/0091-7613(1998)026<0951:EKCSWR>2.3.CO;2
Webb GE (1999) Youngst Early Carboniferous (late Viséan) shallow-water patch reefs in eastern Australia (Rockhampton Group, Queensland): combining quantitative micro-and macro-scale data. Facies 41:111–140. doi:10.1007/BF02537462
Webb GE (2002) Latest Devonian and Early Carboniferous reefs: depressed reef building following the middle Paleozoic collapse: In Flügel E, Kiessling W, Golonka J (eds) Phanerozoic reef patterns. SEPM special publication, vol 72, pp 239–269
Webb GE (2005) Quantitative analysis and paleoecology of earliest Mississippian microbial reefs, lowermost Gudman Formation, Queensland, Australia: not just post-disaster phenomena. J Sediment Res 75:875–894. doi:10.2110/jsr.2005.068
West RR (1988) Temporal changes in Carboniferous reef mound communities. Palaios 3:152–169. doi:10.2307/3514527
Zhang LX (1987) Carboniferous stratigraphy in China. Science Press, Beijing, pp 1–160
Acknowledgments
The study was supported financially by the National Natural Science Foundation of China (Grant no. 40872078) and the Important Direction Project of Knowledge Innovation program of the Chinese Academy of Sciences (Grant No. KZCX3-SW-234). The fieldwork was assisted by Profs. Zhou, H.L. and Zhang, Z.X. from the Geological Institute of Guangxi Bureau of Geology and Mineral Resource.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Shen, JW., Qing, H. Mississippian (Early Carboniferous) stromatolite mounds in a fore-reef slope setting, Laibin, Guangxi, South China. Int J Earth Sci (Geol Rundsch) 99, 443–458 (2010). https://doi.org/10.1007/s00531-008-0392-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00531-008-0392-2