
Abstract
Arabidopsis AP2 (AtAP2) is a member of AP2 subfamily of the AP2/EREBP class of transcriptional factors and plays crucial roles in regulating floral meristem determinacy, floral organ identity and ovule and seed development. In order to examine functional conservation and diversification of AP2 genes in monocots, here we analyze a rice AtAP2-like gene, OsAP2-1. This gene was expressed at high levels in both flowers and roots, and at faint levels in buds or leaves. Its transcript was also present in mature pollen grains. The OsAP2-1 down-regulated lines mediated by RNA interference showed obvious alternation in floral organs of innermost two whorls, including the reduced stamens, fused anther filaments and increased number of pistils. Abortive seeds were observed in the down-regulated lines. These results suggest that OsAP2-1 is required for flower and seed development, consistent with the roles of AtAP2 in Arabidopsis. Furthermore, our results demonstrated that the down-regulation of the endogenous transcripts led to decreased pollen viability and germination activity, suggesting that OsAP2-1 has roles in controlling pollen development.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
EREBP/AP2 proteins, composing a superfamily of plant-specific transcription factors (TFs), are characterized by the presence of a highly conserved ∼70 amino acid region, termed AP2 DNA-binding domain (Weigel 1995; Okamuro et al. 1997). According to the number of AP2 domains, EREBP/AP2 superfamily is divided into two subfamilies: EREBP (ethylene-responsive element-binding proteins), which contains a single AP2 domain, and AP2, that has two AP2 domains (repeat 1 and repeat 2, respectively) in tandem (Weigel 1995; Okamuro et al. 1997). It is generally accepted that EREBP proteins are involved in the response to phytohormones, pathogeny attacks and environmental stress such as cold, drought and high salt (Liu et al. 1998; Haake et al. 2002; Dubouzet et al. 2003; Gutterson and Reuber 2004), whereas AP2 proteins function mainly in the regulation of plant development (Jofuku et al. 1994, 2005; Klucher et al. 1996; Boutilier et al. 2002).
Arabidopsis APETALA2 (AtAP2) is the first identified member of AP2 subfamily by T-DNA tag (Jofuku et al. 1994), and is involved in the establishment of floral meristem identity and the specification of floral organ identity (Kunst et al. 1989; Bowman and Meyerowitz 1991; Jofuku et al. 1994). For example, strong atap2 mutants show the absence of sepals, reduced number of stamens and the replacement of sepals and petals by carpels and stamen identity, respectively (Kunst et al. 1989; Jofuku et al. 1994). AtAP2 is also necessary for the normal development of ovules and seeds (Jofuku et al. 1994) and for the control of seed mass (Ohto et al. 2005; Jofuku et al. 2005). Recent studies have identified numerous AtAP2-like proteins from dicots and monocot maize. Arabidopsis AINTEGUMENTA (AtANT), an AtAP2-like gene, is required for ovule and female gametophyte development (Klucher et al. 1996; Elliott et al. 1996). atant mutants also showed alteration in morphology and number of flower organs in the outer three whorls (Klucher et al. 1996; Elliott et al. 1996). More complete loss of floral organs occurred when atant mutation was combined with atap2 mutation, suggesting that the two genes share the function in initiating floral organ development (Klucher et al. 1996; Elliott et al. 1996). Moreover, AtANT regulates cell proliferation and organ growth by maintaining the meristematic competence of cells during organogenesis (Mizukami and Fischer 2000). BABY BOOM (BnBBM), an AtAP2-like gene from Brassica napus, plays a role in promoting cell proliferation and morphogenesis during embryogenesis (Boutilier et al. 2002). ZmIDS1, the AtAP2-like gene from Zea mays, is required for the control of spikelet meristem fate (Chuck et al. 1998). GLOSS15 (ZmGL15), another AtAP2-like gene from maize, is implicated in specification of leaf epidermal cell identity (Moose and Sisco 1996). All these studies suggest that AP2 proteins should have a more global function besides the role in flower development. Further experiments for additional AP2 proteins from other model plants, such as rice, will be necessary for us to understand the importance of AP2 proteins in plant development.
Here we reported an AtAP2-like gene from rice, termed OsAP2-1. The down-regulation of endogenous OsAP2-1 transcripts mediated by RNA interference cause abnormal flower development and abortive seeds. Furthermore, the down-regulated lines showed the decreased pollen viability, which suggests that OsAP2-1 may be involved in pollen development.
Materials and methods
Plant materials and nuclear acid extraction
Rice plants (Oryza sativa L. japonica cv. Zhonghua 10) were grown under standard greenhouse conditions with a 12-h light/12-h dark period. Flowers were collected from flowering plants. Leaves were collected from 2-week-old seedlings. Roots and buds were harvested from seeds germinated on sterile-water-soaked filter papers. Total RNA was isolated with Trizol kit (Invitrogen) according to the manufacturer’s protocol and treated with RNase-free DNase (Takara) to remove residual genomic DNA. Genomic DNA was extracted from young leaves by cetyltrimethyl ammonium bromide (CTAB) according to the method of Murray and Thompson (1980).
Isolation and sequence analysis of OsAP2-1 cDNA
To isolate OsAP2-1 cDNA, 5 μg of total RNA from rice roots was reversely transcribed into first-strand cDNAs using the ThermoScript kit (Invitrogen), and the first-strand cDNA was used as a template for PCR with OSAP2-1 gene-specific primers P1 (5′-TCGTCTCCTTCCTCCTCTTC-3′) and P2 (5′-TCAGTTGCACACACTGATC-3′). Purified PCR products were cloned into pGEM-T vector (Promega) and confirmed by sequencing.
Amino acid sequence similarity was determined with the BLAST program. Multiple sequence alignment was performed with the DNAMAN, version 4.0 (Lynnon Biosoft) using PAM series matrix, an open gap penalty of 10, an extend gap penalty of 0.05 and a divergent delay of 40%. Protein-sorting signals and localization sites were predicted with the PSORT program (http://www.psort.nibb.ac.jp). Protein motifs were searched using the PPSEARCH (http://www.ebi.ac.uk) and PSI-/PHI-BLAST (http://www.ncbi.nlm.nih.gov) programs.
RT-PCR analysis of OSAP2-1 mRNA accumulation
Five micrograms of total RNA extracted from different organs were reverse-transcribed into first-strand cDNAs, respectively. RT-PCR was performed in a 50 μl mixture containing 1 μl of first-strand cDNA, 10 pmol each of the gene-specific primers P1 and P2 (sequences described in above section), 200 μM dNTPs, 1× PCR buffer and 2.5 U DNA polymerase (5 U/μl, Takara) for 30 cycles. The PCR products were separated on 1% agarose gels and photographed under UV. Rice tubulin mRNA (tubA) was used as a constitutive control (Ding et al. 2001).
Construction of OsAP2-1 RNAi expression vector
In order to easily analyze the functions of a gene by RNA interference (RNAi) technique, we have constructed a RNAi tool vector pWTC605 derived from pCAMBIA1300 (CAMBIA), which contains a construct of 35S pro:MCS1:loop sequence:MCS2 (Zhang et al. 2006). An RNAi expression vector for a given gene can be constructed by inserting a cDNA fragment of the gene into MCS1 and MCS2 in sense and antisense orientations, respectively (Zhang et al. 2006).
Here we constructed OsAP2-1 RNAi construct by inserting a 335 bp fragment spanning from nt 303 to 637 of OsAP2-1 cDNA into pWTC605. To do it, the cDNA fragment was first amplified using primer pair P3 (5′-ACGGATCCTTGCTCCACTTCACTT CAGA-3′, with an added BamHI site) and P4 (5′-CGGGTACCACCCAGGTAGAGGTACTTGTTC-3′, with an added KpnI site), digested with the two restriction enzymes and ligased into pWTC605 in sense orientation. And then it was amplified by primers P5 (5′-ATTGGTTACCTTGCTCCACTTCACTTCAGA-3′, with an added BestEII site) and P6 (5′-ATGTCGACACCCAGGTAGAGGTACTTGTTC-3′, with an added SalI site), digested doubly with BestEII/SalI and ligased into the vector in antisense orientation.
The resulting RNAi construct, OsAP2-1i, was first introduced into Agrobacterium tumefaciens strain EHA105 and then into rice embryonic calli to generate RNAi plants according to the method described by Hiei et al. (1994).
Identification of transgenic plants
After embryonic calli were infected with A. tumefaciens EHA105 carrying OsAP2-1i and subsequently selected on a hygromycin (hyg)-selection medium, 20 independent T0 transformants generated from hyg-resistant independent calli were obtained and planted under standard growth conditions. Resistant plants were confirmed as OsAP2-1i lines using the following strategies: (1) PCR amplification of hyg phosphotransferase gene using the gene-specific primers P7 (5′-TGCTGCTCCATACAAGCCAACC-3′) and P8 (5′-AAGACC TGCCTGAAACCGAACT-3′), and (2) southern blotting examination of insertion of the transgene in these lines.
Southern blot hybridization analysis
For examining the copy number of OsAP2-1 in rice genome, rice genomic DNA (20 μg for each digestion) was digested completely with either KpnI or SaII or SmaI or Xbal. The digested DNAs were subjected to electrophoresis on a 0.8% agarose gel and then blotted onto a Hybond N+ nylon membrane (Amersham Biosciences). Hybridization was carried out overnight at 65°C with a α-32P dCTP-labeled OsAP2-1 cDNA fragment (spanning nt 60–1,341 of the cDNA), which was labeled by a primer-a-gene-labeling system according to the manufacturer’s protocol (Promega).
For detecting insertion of the transgene in OsAP2-1i lines, Genomic DNA (20 μg) from each independent RNAi line was digested with Hind III. Hybridization was carried out overnight at 65°C with a 470-bp α-32P dCTP-labeled DNA fragment of hygromycin phosphotransferase gene.
Each resulting membrane was washed twice in 2× SSC, 0.5× SDS, for 30 min each washing, and twice in 0.1× SSC, 0.1× SDS to suitable signals, and finally autographed at −80°C.
Northern blot analysis of OsAP2-1 transcripts in OsAP2-1i lines
Thirty micrograms of total RNAs extracted from flowers of each of OsAP2-1i and wild-type plants was subjected to 1.5% agarose-formaldehyde gel separation and then transferred to Hybond-N+ membranes (Amersham Biosciences). RNAs in the membrane were fixed by incubation at 80°C for 2 h. Hybridization was performed by a α-32P dCTP-labeled OsAP2-1 cDNA fragment (spanning nt 60–1,341 of the cDNA) at 65°C. The resultant membrane was autographed for 2–3 days at −80°C.
Pollen staining and in vitro germination
Mature pollen grains collected from OsAP2-1i lines as well as wild-type plants during flowering were employed for pollen viability examination by I2–KI or triphenyltetrazolium chloride (TTC) (1% TTC in 50% sucrose) staining. These stained pollen grains were observed under the microscope (ZEISS AXIOSKOP40).
In vitro germination experiments were conducted according to the method described by Wang et al. (2000). Briefly, the freshly collected pollen grains were immediately transferred into a germination medium (20% sucrose, 10% PEG4000, 3 mM Ca(NO3)2, 40 mg/l H3BO3, 3 mg/l VB1) and cultured for a given time at 28°C. The germinated pollen grains were counted from at least six independent observations for each line under the microscope.
Results
OsAP2-1 is a member of AP2 group
Our analysis about the expression of 12 genes encoding deduced rice AtAP2-like proteins, which are predicted from TIGR rice genomic database (accession no. AC093088, AC104433, AC121363, AL732335, AP003573, AP004303, AP005300, AC103891, AP003313, AP004803, AL606446 and AC123525), has identified a gene that showed expression in flowers (data not shown). Genomic sequence of the gene came from BAC clone OSJNBa0056E15 (AC123525, chromosome 11). We further obtained its cDNA sequence by RT-PCR. The cDNA has an open reading frame (ORF) of 1,341 bp encoding a deduced polypeptide of 446 amino acids (accession no. AB247626), here designated as OsAP2-1. The predicted protein has a calculated molecular mass of 49.6 kDa and a pI of 4.65 with relatively hydrophilic feature.
OsAP2-1 contains two AP2 domains in tandem (amino acids 79–147 and 183–243), spaced by a linker of 35 amino acids, at its central region. Besides, it has a serine-rich region of 29 amino acids (AA 5–33) and a bipartite nuclear-targeting sequence (KRRARPETDKSTGKAKRPKK) (AA 40–59) in the N-terminus, and a glutamine-rich region (AA 248–270) immediately downstream of the second AP2 domain. The serine- and glutamine-rich motifs in other transcriptional factors have been proposed as transcriptional activation domains (Jofuku et al. 1994; Elliott et al. 1996).
A blast analysis in the present database using PSI-/PHI-BLAST programme revealed that OsAP2-1 had sequence similarity with Arabidopsis ANT (AtANT, Klucher et al. 1996; Elliott et al. 1996) and AP2 (AtAP2, Jofuku et al. 1994), tobacco ANT (NtANTL, Rieu et al. 2005), Brassica BBM (BnBBM, Boutilier et al. 2002), and IDS1 and GL15 from Maize (ZmIDS1 and ZmGl15, Moose and Sisco 1996; Chuck et al. 1998). The similarities are restricted to the AP2 domains and the linker between them (Fig. 1). Over the region, OsAP2-1 displayed nearly 60% amino acid identity to AtANT, NtANT and BnBBM, otherwise about 40% to AtAP2, ZmIDS1 and ZmGL15. The two AP2 domains in OsAP2-1 both contain the 18 amino acid conserved core regions (Fig. 1), which in AtAP2 are proposed to participate in protein–protein interaction (Jofuku et al. 1994). Furthermore, the key amino acids in the AP2 domains and the linker, which have been proved to be necessary for AtAP2 and AtANT function by mutation analysis (Jofuku et al. 1994; Klucher et al. 1996), are invariable in OsAP2-1 (Fig. 1). All these results suggest that OsAP2-1 is a member of AP2 group. However, a marked difference between AP2 domains of these proteins is that an additional stretch of about 12 amino acids is present in the first AP2 domain of OsAP2-1, AtANT, NtANT and BnBBM, but not in that of AtAP2, ZmIDS1 and ZmGL15 (Fig. 1). In addition, we performed a phylogenetic analysis involving Clustal W alignments of the conserved AP2 domain sequences of these proteins. This result showed that OsAP2-1 tended to group with AtANT, NtANT and BnBBM, but not with AtAP2, ZmIDS1 and ZmGL15 (data not shown). These data indicate that OsAP2-1 is more closed to AtANT, NtANT and BBM proteins than to the others.

OsAP2-1 encodes a two AP2 domain proteins. A multiple alignment of the region spanning the two AP2 domains from OsAP2-1 (Oryza sativa, accession no. AB247626, this study), AtANT (Arabidopsis thaliana, accession no. U41339, Elliott et al. 1996), NtANT (Nicotiana tabacum, accession no. AY461432, Rieu et al. 2005), BnBBM (Brassica napus, accession no. AF317906, Boutilier et al. 2002), AtAP2 (accession no. U12546, Jofuku et al. 1994), ZmIDS1 (Zea mays, accession no. AF048900, Chuck et al. 1998) and ZmGL15 (accession no. AY714877, Moose and Sisco 1996). The first and second AP2 domains are underlined and doubly underlined, respectively. The 18 amino acid conserved core regions proposed to form α-helix in AtAP2 (Jofuku et al. 1994) are shown with filled triangle. The amino acids identified by mutation to be required for AtAP2 (in AP2 domains) and AtANT (in the linker) function are highlighted with “*”
OsAP2-1 is expressed preferentially in flowers and roots
In order to determine copy number of this gene in the rice genome, genomic DNA was digested to completion with either KpnI or SaII or SmaI or Xbal, which does not cut into the cDNA probe, and then subjected to Southern blot analysis. This cDNA probe only detected one band in each case (data not shown). This suggests that the gene is present in rice genome as a single copy gene.
Next, we analyzed the expression pattern of OsAP2-1 using semiquantitative RT-PCR. As shown in Fig. 2a, OsAP2-1 transcripts were accumulated at high levels in flowers, where pollen mother cells in meiotic phase, and roots with faint signals in buds and leaves under the experimental conditions, suggesting its expression is restricted mainly to flowers and roots.

Accumulation patterns of OsAP2-1 mRNA in different tissues, developing flowers and mature pollen grains. Semiquantitative RT-PCR analyses of OsAP2-1 mRNA accumulation were performed by use of the total RNAs extracted from these materials below. Constitutively expressed tubA and/or ribosome RNA were used as controls. a Accumulation patterns of OsAP2-1 transcript in buds (B), flowers (F) which were meiotic stage of male meiocytes, leaves (L) and roots (R). b Expression profiles of the gene in developing flowers, which were in uninucleate microspore stage (F1), bicellular pollen stage (F2) or tricellular pollen stage (F3), and mature pollen grains (P)
Moreover, we collected flowers, which were in uninucleate microspore stage (F1), bicellular pollen stage (F2) or tricellular pollen stage (F3) (Zhang et al. 2006), and mature pollen grains (P), and examined the expression profiles of these genes in the developing of flowers and mature pollen grains. RT-PCR results showed that this gene was expressed in these flowers and mature pollen grains (Fig. 2b).
Generation of OsAP2-1 RNAi transgenic plants
RNA interference (RNAi) can specifically and efficiently result in the degradation of endogenous target transcripts, and subsequent down-regulation of the target gene, so it is a convenient tool of reverse genetics to study gene functions (Chuang and Meyerowitz 2000; Wesley et al. 2001). Therefore, RNAi was used to evaluate the function of OsAP2-1 in rice development. The RNAi construct, OsAP2-1i, was generated by inserting a 335-bp fragment spanning nt 303–637 of OsAP2-1 cDNA, with no similarity to the 11 other rice AtAP2-like genes, into pWTC605 in the sense and antisense orientations separated by a loop sequence (Fig. 3a, also see Materials and methods). In OsAP2-1i, a CaMV35S promoter drove transcription of the sense:loop:antisense fusion sequence (Fig. 3a).

OsAP2-1i lines contain down-regulated endogenous OsAP2-1 transcripts. a A schematic picture of OsAP2-1i constructed by inserting a 335 bp cDNA fragment of OsAP2-1 into pWTC605, an RNAi tool vector (Zhang et al. 2006), at sense and antisense orientations, respectively. b Examination of insertion of the transgene in the RNAi lines L2, L6, L9 and L20 by Southern blot hybridization. 20 μg of genomic DNA extracted from the RNAi lines along with wild-type plants (WT) was separately digested with HindIII. Hybridization was carried out overnight at 65°C with an α-32P dCTP-labeled 470-bp DNA fragment of hygromycin phosphotransferase gene. c Northern blot analysis of OsAP2-1 mRNA in L2, L6, L9 and L20, as well as WT by use of an OsAP2-1 cDNA probe. 28S and 18S RNAs were used as constitutive controls. Quantification of mRNA levels (ratios of normalized data for transgenic lines vs WT) is listed below the picture
After embryonic calli were infected with A. tumefaciens EHA105 carrying OsAP2-1i and subsequently selected on the hygromycin (Hyg)-selection medium, independent transformed plants generated from Hyg-resistant independent calli were planted under standard growth conditions. From these plants, we identified 20 transgenic lines, and each of them contained the insertion of the RNAi construct in the genome, using PCR examination with the Hyg selectable marker gene-specific primer pairs (see Materials and methods). Compared with wild-type plants, which had a >95% seed-setting rate, all the 20 lines had a heavily decreased seed-setting rate (ranging from 10 to 60%). Aberrant flower organs were also observed in these lines. The phenotypes of decreased seed production and aberrant flower development in each line were stable in their regenerated plants from T0 stalks and T1 plants. These results suggest that OsAP2-1 is involved in rice flower development and fertility. T1 plants of 4 (L2, L6, L9 and L20) of the transgenic lines were used for further experiments below.
We examined the insertion copy number of the transgene in L2, L6, L9 and L20 by Southern blotting with a 32P-labeled DNA fragment of the Hyg selectable marker gene of the RNAi construct. Wild-type plants were used as a control. As shown in Fig. 3b, two insertions were detected in L6 and L20, one in L2 and L9, and none in the wild-type control. We found no relation between the insertion number of the transgene and the aberrant phenotypes (Table 1). The uncertainty reflects significant variation of the transgene expression level, which may be affected by many factors, including integration sites and rearrangement (Kohli et al. 2003).
Endogenous OsAP2-1 transcripts are severely down-regulated in the transgenic lines
To investigate whether the OsAP2-1 transcripts were down-regulated in the transgenic lines, endogenous OsAP2-1 transcripts were determined by Northern blot hybridization with RNAs prepared from the flowers of L2, L6, L9 and L20, as well as the wild-type plants, with rRNA as a control to indicate that equal amount of RNAs were loaded in the Northern blot experiments. Compared with the wild-type plants, all the 4 lines contained greatly down-regulated OsAP2-1 transcripts. The normalized data showed that the transcript in L2, L6, L9 and L20 were decreased by 74, 54, 47 and 72%, respectively, as compared with that of the wild-type plants (Fig. 3c). The result indicates that the integration of the RNAi construct in genomes of these lines severely down-regulated endogenous OsAP2-1 transcripts.
Down-regulation of OsAP2-1 results in aberrant flowers
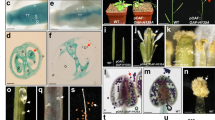
The flowers from wild-type plants have one pistil with two separate stigmas, six stamens, a pair of lodicules, one palea, one lemma and a pair of glumes (Fig. 4a). Compared with wild-type plants, OsAP2-1 down-regulated lines showed abnormal flower organ development (Fig. 4b–f). A quantitative analysis revealed that on average 35% of flowers in these down-regulated lines had aberrant floral organs. These aberrant flowers showed variable phenotypes. Mostly obvious phenotypes observed in the down-regulated lines were alteration in stamens and pistils, including fewer stamens (Fig. 4b–f), additional pistil (Fig. 4e) or additional stigma (Fig. 4b, f). The number of stamens was reduced from six in wild-type flowers to five, four, or even two (Fig. 4b–f) in the mutant flowers. In addition, abnormal stamens that had one anther filament and two separate anthers (Fig. 4b–d) were often seen, suggesting a fusion of filaments from two stamens. In some severely affected flowers, lemma-like structure (Fig. 4e) or palea-like glume (Fig. 4f) was observed. In contrast to OsAP2-1 down-regulated lines, the lines overexpressing OsAP2-1 displayed an increase in the number of stamens (data not shown). These results suggest that OsAP2-1 may be involved in the development of floral organs, specifically of stamens.

Aberrant floral organs in OsAP2-1i lines. a A flower from wild-type plants, which has six stamens and one pistil with two separate stigmas, a pair of lodicules, one palea, one lemma and a pair of glumes. b A flower from the RNAi lines, showing fused anther filaments (arrow) and additional stigmas (arrowhead). c A flower from the RNAi lines, showing fewer stamens with fused anther filaments (arrow). d A flower from the RNAi lines, showing fused anther filaments (arrow) and transformation in shape of paleae and lemma (

OsAP2-1 down-regulated lines display decreased pollen fertility
Since pollen is a key regulator of successful fertilization in sexual plants, we compared the pollen viability of OsAP2-1 down-regulated lines (data only shown for L2) with wild-type control by I2–KI and TTC (2,3,5-triphenyltetrazolium chloride) staining, and in vitro germination experiments (Fig. 5).

Pollen viability of OsAP2-1i lines is severely affected. Viability of mature pollen grains from wild-type (a, c, e) and the RNAi lines (data only shown for L2) (b, d, f) were examined with I2–KI staining (a, b), TTC staining (c, d) and in vitro germination experiments (e, f). Scale bars in a = 50 μm for (a) to (f), t pollen tubes
In an I2–KI staining assay, ∼98% of pollen grains (n = 537) from the wild-type control were round and had a uniform size, with a dark blue-black reaction (Fig. 5a). Pollen grains of the down-regulated lines had a similar I2–KI staining reaction (96%, n = 560) and morphological characterization (Fig. 5b), suggesting down-regulation of OsAP2-1 may not affect early development of pollen. Furthermore, in TTC staining, where TTC can be reduced to a red product by NADPH from the redox reaction of respiration metabolism occurring in viable pollen grains when they are in a rehydration environment, only ∼43% of pollen grains (n = 932) from L2 was found to be “viable” (red) and the others are “nonviable” (yellow) (Fig. 5d), and a similar situation was observed in L9, whereas ∼90% of pollen grains (n = 1,743) were viable in the wild-type (Fig. 5c).
Finally, we checked in vitro germination activity of mature pollen grains from the RNAi lines as well as wild-type plants. Under the in vitro germination condition, 75% (n = 610) of mature pollen grains from the wild-type plants germinated (Fig. 5e), whereas only 22% (n = 660) of mature pollen grains germinated in the RNAi lines (Fig. 5f). This indicated that OsAP2-1 deficiency affected pollen viability.
OsAP2-1 down-regulated lines displays aborted seeds
When examining the seed-setting rate of the down-regulated lines and wild-type plants, we found a high proportion of abortive seeds in the RNAi lines (ranging 16–30% in examined the four lines) (Table 1), which had less than half the size of fully developed seeds and appeared shrunken at mature and dehydrate stage of seeds. This indicated that the seed development in the RNAi lines was disrupted. Under the same growth condition, wild-type plants only had 3.88% of abortive seeds. The result demonstrated that OsAP2-1 is required for normal development of seeds in rice.
Discussion
OsAP2-1 encodes a double AP2 domain protein. Its relatively hydrophilic and acidic features, combined with the presence of nuclear-targeting motif, suggest that this protein has a potential nucleus-localizing function. The serine- and glutamine-rich motifs identified in OsAP2-1 are also present in the Arabidopsis ANT protein and some other transcriptional factor, and have been proposed to act as transcriptional activation domains (Elliott et al. 1996). A recent study (Ye et al. 2004) has isolated a partial cDNA sequence (AJ575217) encoding N-terminal part of OsAP2-1 by yeast two hybridization. The partial polypeptides have an ability to activate transcription of reporter gene in this yeast-based system (Ye et al. 2004). This suggests that OsAP2-1 may act as a transcriptional regulator in rice.
OsAP2-1 displayed amino acid sequence similarity with the present known members of AP2 group, such as AtAP2 (Jofuku et al. 1994), AtANT (Klucher et al. 1996; Elliott et al. 1996), NtANT (Rieu et al. 2005), BBM (Boutilier et al. 2002), ZmIDS1 (Chuck et al. 1998) and ZmGL15 (Moose and Sisco 1996), in the two domains and the linker between them (Fig. 1). Outside the AP2 domain region there is little sequence similarity between OsAP2-1 and each of these proteins. The key amino acids in the AP2 domains and the linker required for AtAP2 and AtANT function, identified by mutation analysis (Jofuku et al. 1994; Klucher et al. 1996), are nonviable in OsAP2-1 (Fig. 1). This indicated that OsAP2-1 is a new member of AP2 group. Furthermore, Like AtANT and BnBBM, OsAP2-1 contains a 12-AA stretch in its first AP2 domain, but the stretch is absent in AtAP2, ZmGL12 and ZmIDS1 (Fig. 1). A phylogenetic analysis reveals that OsAP2-1 tends to group with AtANT, NtANT and BnBBM, indicating OsAP2-1 may be an ANT/BBM-like protein.
We obtained OsAP2-1 down-regulated lines mediated by RNAi. The down-regulated lines contained severely down-regulated OsAP2-1 transcripts and displayed abnormal floral organs including the modified palea, lemmas and glumes in shape, additional lemma-like organs, additional pistil or stigma and a decrease in stamen number (Fig. 4), similar to those of Arabidopsis ant mutants and ap2 mutants (Jofuku et al. 1994; Klucher et al. 1996; Elliott et al. 1996). This indicates that OsAP2-1 is required for floral organ development in rice. However, the fused anther filaments in OsAP2-1 RNAi lines were not observed in the two arabidopsis mutants (Jofuku et al. 1994; Klucher et al. 1996; Elliott et al. 1996). These phenotypes including fused anther filaments, fewer stamens and more than one carpel observed in the down-regulated lines indicate the possible roles of OsAP2-1 in controlling floral meristem determinacy.
Besides abnormal floral organs, the OsAP2-1 down-regulated lines also showed decreased seed-setting rate. Although stamen number in the down-regulated lines was reduced, the surviving stamens appeared to give rise to a similar amount of pollen grains to those from wild-type plants. However, most of mature pollen grains from the down-regulated lines were metabolically nonviable and unable to germinate (Fig. 5). RT-PCR analyses revealed that the gene was expressed in the examined flowers from meiotic stage of male meiocytes to biuncleate pollen stage, and mature pollen grains. Taken together, these results indicate that OsAP2-1 is involved in pollen development. Further experiment is necessary to examine how this gene affects pollen development.
Furthermore, the down-regulated lines had a higher proportion of abortive seeds. A previous expression pattern analysis of the gene in developing rice seed has revealed that OsAP2-1 transcripts were accumulated preferentially in early developing seeds (Ye et al. 2004). Taken together, these results suggest that the gene is required for seed development. A recent study revealed that AtAP2 and BnBBM have roles in promoting cell proliferation and morphogenesis during embryogenesis. The high similarity between AP2 domain region of OsAP2-1 and BBM (60% identity) suggest that abortive seeds in the down-regulated lines may result from disrupted cell proliferation and/or morphogenesis of fertilized ovaries. However, the present results do not exclude the possibility that abortive seeds come from the enlargement of unfertilized carpels.
In summary, we have isolated two AP2 domain proteins from rice, OsAP2-1. This protein is more closed to AtANT, NtANT and BnBBM proteins in the region spanning the two AP2 domains. Furthermore we obtained OsAP2-1 RNAi lines, whose endogenous OsAP2-1 transcripts were severely down-regulated. These down-regulated lines displayed abnormal floral organs and abortive seeds. Our results also revealed that down-regulation of OsAP2-1 in the RNAi lines led to decreased pollen viability and germination activity.
References
Boutilier K, Offringa R, Sharma VK, Kieft H, Ouellet T, Zhang L, Hattori J, Liu CM, van Lammeren AAM, Miki BLA, Custers JBM, van Lookeren Campagne MM (2002) Ectopic expression of BABY BOOM triggers a conversion from vegetative to embryonic growth. Plant Cell 14:1737–1749
Bowman JL, Meyerowitz EM (1991) Genetic control of pattern formation during flower development in Arabidopsis. Symp Soc Exp Biol 45:89–115
Chuang CF, Meyerowitz EM (2000) Specific and heritable genetic interference by double-stranded RNA in Arabidopsis thaliana. Proc Natl Acad Sci USA 97:4985–4990
Chuck G, Meeley RB, Hake S (1998) The control of maize spikelet meristem fate by the APETALA2-like gene indeterminate spikelet1. Genes Dev 12:1145–1154
Ding ZJ, Wang T, Chong K, Bai SN (2001) Isolation and characterization of OsDMC1, the rice homologue of the yeast DMC1 gene essential for meiosis. Sex Plant Reprod 13:285–288
Dubouzet JG, Sakuma Y, Ito Y, Kasuga M, Dubouzet EG, Miura S, Seki M, Shinozaki K, Yamaguchi-Shinozaki K (2003) OsDREB genes in rice, Oryza sativa L., encode transcription activators that function in drought-, high-salt- and cold-responsive gene expression. Plant J 33:751–763
Elliott RC, Betzner AS, Huttner E, Oakes MP, Tucker WQJ, Gerentes D, Perez P, Smyth DR (1996) AINTEGUMENTA, an APETALA2-like gene of Arabidopsis with pleiotropic roles in ovule development and floral organ growth. Plant Cell 8:155–168
Gutterson N, Reuber TL (2004) Regulation of disease resistance pathways by AP2/ERF transcription factors. Curr Opin Plant Biol 7:465–471
Haake V, Cook D, Riechmann JL, Pineda O, Thomashow MF, Zhang JZ (2002) Transcription factor CBF4 is a regulator of drought adaptation in Arabidopsis. Plant Physiol 130:639–648
Hiei Y, Ohta S, Komari T, Kumashiro T (1994) Efficient transformation of rice (Oryza sativa L.) mediated by Agrobacterium and sequence analysis of the boundaries of the T-DNA. Plant J 6:271–282
Jofuku KD, den Boer BGW, Van Montagu M, Okamuro JK (1994) Control of Arabidopsis flower and seed development by the homeotic gene APETALA2. Plant Cell 6:1211–1225
Jofuku KD, Omidyar PK, Gee Z, Okamuro JK (2005) Control of seed mass and seed yield by the floral homeotic gene APETALA2. Proc Natl Acad Sci USA 102:3117–3122
Klucher KM, Chow H, Reiser L, Fischer RL (1996) The AINTEGUMENTA gene of Arabidopsis required for ovule and female gametophyte development is related to the floral homeotic gene APETALA2. Plant Cell 8:137–153
Kohli A, Twyman RM, Abranches R, Wegel E, Stoger E, Christou P (2003) Transgene integration, organization and interaction in plants. Plant Mol Biol 52:247–258
Kunst L, Klenz JE, Martinez-Zapater J, Haughn GW (1989) AP2 gene determines the identity of perianth organs in flowers of Arabidopsis thaliana. Plant Cell 1:1195–1208
Liu Q, Kasuga M, Sakuma Y, Abe H, Miura S, Yamaguchi-Shinozaki Y, Shinozaki K (1998) Two transcription factors, DREB1 and DREB2, with an EREBP/AP2 DNA binding domain separate two cellular signal transduction pathways in drought- and low-temperature-responsive gene expression, respectively, in Arabidopsis. Plant Cell 10:1391–1406
Mizukami Y, Fischer RL (2000) Plant organ size control: AINTEGUMENTA regulates growth and cell numbers during organogenesis. Proc Natl Acad Sci USA 97:942–947
Moose SP, Sisco PH (1996) Glossy15, an APETALA2-like gene from maize that regulates leaf epidermal cell identity. Genes Dev 10:3018–3027
Murray MG, Thompson WF (1980) Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res 8:4321–4325
Ohto MA, Fischer RL, Goldberg RB, Nakamura K, Harada JJ (2005) Control of seed mass by APETALA2. Proc Natl Acad Sci USA 102:3123–3128
Okamuro JK, Caster B, Villarroel R, Van Montagu M, Jofuku DK (1997) The AP2 domain of APETALA2 defines a large new family of DNA binding proteins in Arabidopsis. Proc Natl Acad Sci USA 94:7076–7081
Rieu I, Bots M, Mariani C, Weterings KAP (2005) Isolation and expression analysis of a tobacco AINTEGUMENTA ortholog (NtANTL). Plant Cell Physiol 46:803–805
Wang SH, Chen F, Zhou KD (2000) In vitro germination of rice pollens. Acta Agronomica Sinica 26:609–611
Weigel D (1995) The APETALA2 domain is related to a novel type of DNA binding domain. Plant Cell 7:388–389
Wesley SV, Helliwell CA, Smith NA, Wang M, Rouse DT, Liu Q, Gooding PS, Singh SP, Abbott D, Stoutjesdijk PA, Robinson SP, Gleave AP, Green AG, Waterhouse PM (2001) Construct design for efficient, effective and high-throughput gene silencing in plants. Plant J 27:581–590
Ye R, Yao QH, Xu ZH, Xue HW (2004) Development of an efficient method for the isolation of factors involved in gene transcription during rice embryo development. Plant J 38:348–357
Zhang L, Tao J, Wang S, Chong K, Wang T (2006) The rice OsRad21–4, an orthologue of yeast Rec8 protein, is required for efficient meiosis. Plant Mol Biol 60:533–554
Acknowledgments
The research was supported by grants from NSFC (No 30370138) and The Chinese Ministry of Sciences and Technology (Nos: 2005CB120802 and 2005CB120804).
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Thomas Dresselhaus
Rights and permissions
About this article
Cite this article
Zhao, L., Xu, S., Chai, T. et al. OsAP2-1, an AP2-like gene from Oryza sativa, is required for flower development and male fertility. Sex Plant Reprod 19, 197–206 (2006). https://doi.org/10.1007/s00497-006-0036-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00497-006-0036-2