
Abstract
The objective of this paper was to evaluate the effects of high temperatures on calves. The hypothesis that the red and white blood cells, health, and performance of calves would be influenced by the temperature period at birth and gender was tested. Sixty-three Holstein calves were used. They were reared in individual hutches from the second day of life to weaning at the age of 8 weeks. All calves were allotted to treatment groups according to the temperature period at birth: moderate temperature period 1 (MT1), high temperature period (HT), moderate temperature period 2 (MT2). The same conditions of nutrition were ensured. We recorded 62 summer days and 14 tropical days during HT. Sixty-six days with a value above 72.0 of the temperature–humidity index (THI) and 26 with values greater than 78.0 were found. No significant differences were found in red blood cells with the exception of hemoglobin between MT1 and MT2 (p = 0.031) during the 8th week. Significant differences were observed among treatment groups in the percentage of eosinophiles during the 6th week (p = 0.044). The HT calves had the least body weight from the first week to weaning (p = 0.053) and the highest water consumption (p = 0.042). The results emphasize the ability of dairy calves to maintain homeostasis during prolonged periods of heat stress. The exposure to high temperatures resulted only in a significant decrease of starter concentrate consumption and reduced growth.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Heat stress generally causes a decrease in feed intake. Therefore, high temperatures inhibit growth and mass increase in cattle, depending on genotype, age, and adaptability. Not only performance is reduced, as heat stressed cattle can increase water intake, stand rather than lie down, and increase respiration rate and body temperature, as well as, in the most severe cases, die, thus resulting in economic loss in revenue to the cattle industry (Blackshaw and Blackshaw 1994; Nardone et al. 2006). Because the main benefit from dairy cattle is milk production, protecting dairy cows against heat stress is preferred and amply citated in scientific literature. However, temperature stress is generally forgotten and disregarded in calves, almost all the papers focussing on dairy cows or feedlot cattle (Mitloehner et al. 2002; West 2003; Mader and Davis 2004; Mader et al. 2006). The most difficult situation occurs in outdoor-rearing of calves. Rearing calves in individual outdoor calf hutches has been promoted throughout Slovakia since the early 1980s (Broucek et al. 1988), primarily because individual outdoor housing reduces transmission of diseases. Coincident with the development of commercial plastic hutches that retain solar radiation, the popularity of calf hutches has also increased in cooler northern climates.
The main advantage of the hutches for calves is the minimized risk of disease transfer from calf to calf; however, calves in hutches apparently do not have the possibility of protection against high temperatures. They cannot emit heat by convection or conduction, as that is possible only when air temperatures are lower than the temperature of their skin, and they cannot seek out shade (Soch 2005; Schäffer and von Borell 2008). Calves in hutches with a yard are free to move around and seek shelter in shade inside the hutch; however, a yard is generally too small and the hot roof of the hutch does not guarantee much respite from extreme heat. However, the accumulation of solar heat inside plastic hutches could be a distinct disadvantage in warmer southern climates (Broucek et al. 1990b; Spain and Spiers 1996; Coleman et al. 1996; Schäffer and von Borell 2008). Radiant heat load on calf hutches may add to heat stress by increasing interior hutch temperature (Carter et al. 2008). The thought of calves staying in hutches with a roof baked by the sun also causes stress in humans.
Hutches placed in direct sunlight showed a higher internal ambient temperature due to a transfer of heat from the outer surface. This resulted in higher skin temperatures and respiration rates (Spain and Spiers 1994). These clinical signs of high temperature stress included open-mouthed panting, drooling, reluctance or inability to rise, increased licking of coat, and general dullness including neurological signs with staring and glazed eyes. The calf under shade is protected from the direct rays of the sun; however, it is still exposed to large amounts of diffuse solar energy (Blackshaw and Blackshaw 1994). Covering hutches with reflective insulation reduced interior temperatures by 1.4°C at a high temperature–humidity index (THI), but increased interior temperatures by 0.58°C at moderate THI (Carter et al. 2008). During summer, ventilation was improved markedly by removing the back wall. Painting plastic calf hutches white and improving ventilation by opening the back reduced heat stress (Hoshiba et al. 1985). Shaded areas should be provided for the summer time if hutches are used (Schäffer and von Borell 2008). According to Bray et al. (1997), calves housed in calf hutches in the summer exhibited elevated body temperatures similar to those shown by calves housed in crates under a roof. Calves in the hutches had greater mortality and lower serum IgG than calves housed under corrugated metal shades with no side walls. Hutches were hotter and resulted in significantly greater stress and death losses of calves (Stott et al. 1976, cited in West 2003).
Hoshiba et al. (1988) indicated that the calf hutch especially has the role of protection from rain. The role of protection from heat stress in a hot climate was only estimated in the white-painted plywood calf hutches. The average inside temperature was 2.7°C lower than the average outside temperature. In a comparison of different types of hutches, Broucek et al. (1990b) found that the highest temperatures inside hutches (35°C) were in gray-colored fiberglass hutches (at the height of the wither of a standing calf) and 41°C under the roof when the outside temperature was 31°C. Broucek et al. (1990a) reported that the average temperature and relative humidity were, respectively, 24.4°C and 65.9% in wooden hutches, 23.3°C and 69.2% in calf barns, and 22.9°C and 65.8% outside.
In zones with hot temperatures, young cattle grow more in spring and autumn seasons than in summer. Temperatures between 15 and 29°C do not seem to exert any influence on growth performance. Above 30°C, adverse effects are recorded in daily weight gain (Nardone et al. 2006). The measurement of clinical and hematological parameters could be used to evaluate welfare in cattle. During hypothermic stress, metabolic intermediate products accumulate in blood and hemoglobin-concentration rises with a plasma volume decrease. Heat exposure by direct solar radiation for 6 h during which the environmental temperature rose from 32°C to 52°C resulted in a significant increase of neutrophiles and decrease of eosinophiles (May et al. 1977).
Diseases of newborn and postnatal mortality are a major cause of economical losses in dairy production. Thus, specific hematological reference ranges are needed for each species to help promote the ability of scientists and veterinarians to make more accurate interpretation of clinical pathology data (Seifi et al. 2006).
The aim of this study was to investigate effects of temperatures on the hematological parameters, health, and performance of calves kept in individual outdoor hutches. The hypothesis that the observed parameters would be influenced by the season at birth and gender was tested.
Materials and methods
Sixty-three (30 females and 33 males) Holstein calves which showed high vitality at birth (respiration immediately after birth, standing up and suckling soon after) were analyzed. They were reared in fiber-glass laminated calf hutches (white color) from the second day of life to weaning at the age of 8 weeks. Each hutch (1.8 × 1.2 m) had an outside fenced yard of 1.8 × 1.2 m and was bedded with straw. Hutches were arranged in rows, 0.8 m apart, oriented east to west to minimize exposure to solar heating. Hutches remained in the same location for the duration of the study. Cold drafts from blowing winter winds were blocked by the walls.
After 24 h spent with their mothers, the calves received mother’s milk ad libitum three times a day from a bucket with a nipple from the second to fourth day. From the fifth day, they received 6 kg of MR (milk replacer) per day divided into 2 portions at 12-h intervals. From the second day until weaning, the calves could eat starter mixture and alfalfa hay ad libitum. Feed and water refusals were removed and weighed each morning before feeding throughout the experiment. Observation of the appetite was measured twice daily during each feeding.
Air temperature and relative humidity were accurately recorded by measuring equipment during the entire 24-h period. The equipment was placed in the center of the calf hutches area at the same height as the withers of a standing calf.
All calves were divided into three groups according to the temperature period at birth. The periods were determined as follows: moderate temperature period 1 (MT1) from 4 April to 11 June (69 days, n = 18, 12 males and 6 females), hot temperature period (HT) from 12 June to 14 September (95 days, n = 30, 15 males and 15 females), moderate temperature period 2 (MT2) from 15 September to 24 November (70 days, n = 15, 6 males and 9 females). The periods were designated according to the daily maximum temperature; the limit was 25.0°C. The average maximum temperatures and THI were, respectively, 19.5°C and 65.2 (MT1), 26.5°C and 74.8 (HT), and 15.7°C and 59.7(MT2).
We determined, according to the maximum temperature, the number of summer days (maximum temperature above 25.0°C) and tropical days (maximum temperature above 30.0°C) from 24-h records (Soch 2005). The THI was calculated as proposed by Nienaber et al. (1999) by combining maximum temperature (in °C) and average relative humidity (%) per day using the following expression: THI = (0.8 × Tmax) + (% average RH/100) × (Tmax – 14.4) + 46.4.
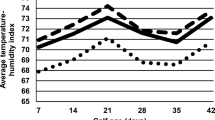
We recorded 62 summer days and 14 tropical days for the HT period. Sixty-six days with the value of THI above 72.0, which is already a stressor, were found during the period. The total number of days with values higher than 78.0, which was a substantial stress, was 26 for the whole HT period (Fig. 1).

The course of temperatures and the temperature–humidity index during the experiment. MT1 Moderate temperature period 1 from 4 April to 11 June (69 days), HT hot temperature period from 12 June to 14 September (95 days), MT2 moderate temperature period 2 from 15 September to 24 November
Blood samples for analysis of white and red blood compounds were taken every second week. Samples were obtained between 0800 and 0900 hours, 2 h after the morning feed. Blood was taken via jugular venipuncture in 10-ml tubes containing disodium-EDTA. All tubes were placed immediately in an ice portable refrigerator and transferred to the laboratory. Hemoglobin concentrations were analyzed spectrophotometrically using a standard cyanmethemoglobin method (van Kampen solution) at a wavelength of 540 nm. Hematocrit values (packed cell volume, PCV) were determined by means of the centrifuge microhematocrit technique (MPW-310). Differential white blood cell count was performed microscopically with the use of an immersion lens. Proportions of individual leukocyte types (in 200 cells) were computed from blood smears panoptically stained according to Pappenheim. The blood smears which were air-dried and stained with the Pappenheim method (May-Grünwald + Giemsa-Romanowski) (Broucek and Kovalcik 1989). The total erythrocyte and leukocyte counts were determined by means of the flask method of dilution and corpuscles counted using the Bürker chamber (Jagos et al. 1985).
Methods of measuring and describing the health condition vary widely and in many cases are incomplete. In the present study, the classical method of Larson et al. (1977) for the evaluation and expression of diarrhea was used. Color (yellow, code 0; green, code 1; white, code 2; red, code 3; pink, code 4; brown, code 5; dark or black, code 6) and consistency (liquid, code 0; normal, code 1; firm, code 2) of feces and health condition was evaluated by the code system twice a day together with rectal temperature measurements at the time of feeding.
Respiratory condition (Pneumonia) was assessed by the types of symptom (normal, runny nose, heavy breathing, and cough – moist or dry). Other frequency of cough (possible respiratory disorder) as occasional, intermittent, or persistent. Operators observed the condition of hair and eyes (dullness and brightness) and signs of dehydration (sunken eyes, inelastic skin, and prostration) (Larson et al. 1977).
The data were analyzed using a General linear model ANOVA (two-way with the interactions) by the statistical package STATISTIX, Version 8.0 (Anonymous 2001). The dependent variables were hematological parameters, body weight, average daily gains, consumption of feeds, and weeks of observation, and the independent variables were the temperature period and gender. Significant differences among means were tested by Bonferroni’s test. We did not find statistically significant differences in the evaluation of gender and weeks of observation.
The normal distribution of data was evaluated by Wilk-Shapiro/Rankin Plot procedure. We found that, in the case of the color and consistency of feces assessment, one or a few points departed from the linear trend of the plot indicating non-normality. Approximate Wilk-Shapiro normality statistics were also lower. For this reason, the Kruskal-Wallis ANOVA procedure was more suitable for the comparison of groups. Significant differences between groups were tested by Comparisons of Mean Ranks. All values are reported as means ± standard error of the mean (Anonymous 2001).
Results
Results of blood values from four sampling periods during the milk feeding period are shown in Tables 1 and 2. There were no temperature period and gender effects (p > 0.05) for blood measurements of red blood cells with the exception of hemoglobin during the 8th week, when the difference between MT1 and MT2 was significant (p = 0.031) (Table 1).
We did not find any significant differences among observed factors in the number of leukocytes or percentage of basophiles, monocytes, and neutrophiles. Significant interactions (p < 0.05) between treatment group and gender were recorded in leukocytes during the 6th week (Table 2).
Significant differences were found among treatment groups in percent of eosinophiles during the 6th week (MT1 4.24 ± 0.24%; HT 5.02 ± 0.21%; MT2 4.19 ± 0.31%; p = 0.044) (Table 2).
There were no differences among temperature period and gender in the color and consistency of feces, although diarrhea is important in evaluating the response of health of calves to experimental conditions. Color showed a steadily trend from yellow to green and consistency changed smoothly from liquid to normal.
Calves born in the HT had the highest water consumption during all experiment (p = 0.042) (Table 3). We found significant differences among groups in weeks 2 (p = 0.022), 3 (p = 0.019), 4 (p = 0.033), 5 (p = 0.045), and 6 (p = 0.021).
The highest consumption of the starter mixture was found in the group MT2 in the all-milk drinking period (p = 0.038) (Table 3). In each week, the calves born during the HT period received the least mixture. The greatest differences were obtained between temperature period groups from the third to fifth weeks (p < 0.001) and then in the first and sixth weeks (p < 0.05).
Calves born in the MT1 period received the least alfalfa hay to weaning on the 56th day (18.21 ± 1.44 kg) and calves born in the MT2 the most (24.63 ± 1.60 kg; p = 0.045); only minimal difference was found between MT2 and HT (Table 3). However, we obtained the highest consumption of hay in group HT in the seventh (MT1 4.24 ± 0.43 kg, HT 6.43 ± 0.34 kg, MT2 6.02 ± 0.48 kg, p = 0.037) and eighth (MT1 4.97 ± 0.42 kg, HT 6.77 ± 0.34 kg, MT2 5.69 ± 0.47 kg, p = 0.022) weeks. Because the feeding was the same throughout the experiment, the effects associated with this phenomenon are not based on food differences.
The calves born during the HT period had the least body weight from the first week to weaning (p = 0.053). The highest growth was recorded in calves born in the MT2 period. A significant difference in the average daily gains was found in the first week (MT1 0.55 ± 0.04 kg; HT 0.46 ± 0.04; MT2 0.69 ± 0.71 kg; p = 0.029) and for the whole period from birth to weaning (MT1 0.44 ± 0.02 kg; HT 0.39 ± 0.02; MT2 0.48 ± 0.2 kg; p = 0.048).
Discussion
Reducing heat stress in calves is not the first concern during summer days, because dairy cows have a priority. However, the fact is that the effects of hot temperature stress often go unnoticed in calves. They may be suffering, but the cows need more attention.
We did not record such high temperatures in our experiment as they do in the southern states of Europe, but we found temperatures above the threshold of thermoneutrality; that is, mean THI values higher than 72 and even higher than 78 (Mader and Davis 2004; West 2003). Although knowledge of THI alone is beneficial in determining the potential for heat stress, wind speed and solar radiation adjustments to the THI more accurately assess animal discomfort (Huber 1996; Mader et al. 2006). The average total number of days with high values of THI and duration of events is also important for calf management decisions. As indicated by Hubbard et al. (1999), young cattle can withstand short periods of high THI values when they have adequate recovery time between episodes; it depends on the length of the period of exposure to elevated temperatures which can be 1 or 2 weeks (acute thermal conditions) or 3–8 weeks or more (chronic) (Nienaber et al. 1999).
A study by Brown-Brandl et al. (2003) determined that body temperature, breathing rate, and metabolic rate were affected by air temperature. Given the potential problems in temperature effects, calves housed outside could be exposed to significant stress (Spain and Spiers 1996). To cope with a hot environment, cattle use a variety of strategies: behavior (shade-seeking, orientation to the sun, increased water intake); increases in tissue conductance to facilitate heat transfer from the body core to the skin, in skin temperature to raise heat loss by convection and radiation, in sweating rate to increase skin evaporative loss, and in respiratory volume to increase respiratory evaporative heat loss; and a fall in metabolic rate (longer term). If these strategies fail and the animal is unable to regulate its body temperature, the heat load exceeds the body’s capacity to eliminate it, the body temperature will rise, and the animal may suffer heat stroke and death (Blackshaw and Blackshaw 1994; Soch 2005). Hot temperatures aggravated by high humidity play a cardinal role in animal morbidity and mortality. Sickness rate increases at THI values in the emergency category (Du Prezz et al. 1990). Other metabolic and endocrine responses to heat stress have been reviewed by Davis and Drackley (1998) and Soch et al. (1999).
During the present study, the parameters of red and white blood counts corresponded to the reference range previously reported (Meyer and Harvey 2004; Mohri et al. 2007). Heat stress can result in sick animals which require prolonged care. However, no significant differences were found in health: no calf died, nor was culled because of bad health. Rectal temperature measurements did not record any signs of health problems. Similarly, a distinct loss of appetite was not observed.
However, during the hottest period, blood parameters remained unchanged. Our results suggest that calves with the experience of physiological changes during exposure to prolonged and continuous high heat and humidity showed little reaction. High vitality new-born calves are able to face adverse climatic conditions (Jagos et al. 1985; Broucek et al. 1987; Beatty et al. 2006). In contrast, low vitality calves, born under unfavorable conditions (dystocia, premature), are handicapped by hypoxia, acidosis, and hyperlactataemia as well as by a smaller mobilization of body lipids (Illek et al. 2007). We did not observe such effects in calves, probably because, unlike previous studies, our animals had nocturnal respite from the heat. Young calves have a well-developed power of acclimation to high temperatures. In the study of Singh and Newton (1978), calves were exposed to 40.5°C and 50% relative humidity in a climatic chamber during 12 h each day for 2 weeks. Their physiological parameters rose sharply on the first day of exposure and then gradually declined. However, most of the acclimation of physiologic measurements was completed in the first 9–10 days of exposure.
Heat stress is responsible for alterations of the immune response which changes depending on the type of immune response and the length of time that calves are exposed to the stressor (Broucek and Kovalcik 1989). Conversely, different studies, carried out either under field conditions or in a controlled environment, documented a higher mortality rate or impairment of passive immunization in calves born during the summer (Lacetera et al. 2005; Illek et al. 2007). There is no relationship between the decline of average daily gains and the occurrence of clinical signs of heat stress in animals. Results from our study would indicate that moderate heat stress does not modify the protective effect of colostrums significantly, as evaluated by determining the concentration of Ig fractions. Cows born in the summer showed lower milk yield and FCM yield. The most productive were cows born in the winter and spring (Broucek et al. 2006). Nardone et al. (1997) reported lower immunoglobulin content in the colostrum of primiparous cows exposed to heat stress prepartum.
Hot environments are also detrimental to the productivity of animals. The upper limit of the zone of thermoneutrality for young calves is at an effective environmental temperature of about 26°C (Davis and Drackley 1998). As the environmental temperature increases beyond this point, termed the point of hyperthermal rise or upper critical temperature, the animal must expend energy to dissipate heat from its body. This occurs primarily through increased respiration (panting) in an effort to increase evaporative losses of heat.
The heat stress problem is acutely felt in southern European countries. The weather is characterized by high summer temperatures coupled with high humidity levels. Calves born in the HT period had the highest water intake. Drinking water may have a direct comfort effect by cooling the reticulum as well as reducing the thermal load (Blackshaw and Blackshaw 1994).
Maximum water intake during the HT period almost doubled the water intake of other periods, corresponding with previous studies cited by Beatty et al. (2006). However, this increase could be related to a higher consumption of dry-matter, because a significant increase in consumption of alfalfa hay was recorded compared to MT1; thus, it was mainly caused by an increased intake of rough feed. The mechanism for the increase in water consumption may, like feed intake, involve direct effects of the heat, because directly warming the preoptic area and rostral hypothalamus of the ruminants caused a large increase in water consumption (Soch et al. 1999; Robertshaw 2006). Although depleted water stores due to evaporative demands could induce increased water intake via hypovolemia or hyperosmolarity, neither of these changes occurred in this study, suggesting the direct effect of heat. There were some indications for an increase of total blood volume as indicated by a decrease of hematocrit.
The cumulative effect of prolonged heat load was also evident on feed intake. Cattle reduce feed intake in response to heat stress. A reduction in feed intake is followed by a decline in metabolic rate and, therefore, reduced heat production, which helps to maintain heat balance (Mitloehner et al. 2002). Although knowledge of THI alone is beneficial in determining the potential for heat stress, wind speed and solar radiation adjustments to the THI more accurately assess animal discomfort (Mader et al. 1999). The etiology of the decreased starter mixture and increased alfalfa hay intake for the HT treatment group is unknown, but it was significantly associated with the elevation of temperature. Whether it was a direct effect of heat or hormonally mediated cannot be determined from these experiments.
Calves born in summer showed a tendency for decreased average daily gain and starter consumption. The reason that the HT period displayed more negative effects is not obvious. Minimizing heat loads of outside-kept calves during hot weather is important from both production and animal welfare standpoints. In the experimental facility, hutches were localized in the same area, arranged in rows, and oriented east to west to minimize exposure to solar heating. On the other hand, calves born in spring or fall would be expected to have access to the best forage. However, this should not be true provided the milk replacer, starter mixture, and alfalfa hay were the same and stable throughout the year. Effects of heat stress on performance are mainly a result of a decrease in daily matter intake; however, consumption of afalfa hay differed only in comparison to calves born during the fall period (from 15 September to 24 November). Hyperthermic stress was alleviated by depression of the appetite, but receiving of bulk feed was not reduced.
Values of THI recorded during the HT period indicated that summer-born calves were exposed during the daytime to conditions capable of inducing moderate heat stress. Furthermore, it must be noted that values of THI recorded during nights were not markedly below 72, which is considered the upper critical THI for dairy cows (Johnson 1987). However, this result would suggest that during the summer period the extent of night-time THI decline allows some slight alleviation of stress associated with daytime heat and humidity. Hahn and Mader (1997) also indicated that several hours with THI above 84 and limited or no night-time recovery periods with THI under 74 can result in death of vulnerable animals unless immediate action is taken to limit excessive heat loads. Their results emphasize the importance of recovery periods, as well as heat intensity. Calves can adjust to higher temperatures, but that does not mean they do not suffer long-term effects, like reduced growth rates. Especially, heat waves are a prime example of weather events which strongly impact the dynamic response of cattle (Brown-Brandl et al. 2004). Hahn (1995, cited in Hahn and Mader 1997) has reported that cattle require about 3-4 days to balance heat production, resulting from metabolizing feed, with their heat dissipation capabilities.
Conclusion
Results of this study indicated that hyperthermic stress is a negative factor of calves' environment. The calves born during the period with higher temperatures had the lowest average daily gains of live body weight, drank the most of water to weaning, and received the least starter mixture from the first week to weaning. We can conclude that heat stress has no negative effect on the hematological parameters and health condition in calves kept in hutches.
References
Anonymous (2001) Statistix for Windows. Version 8.0. User’s Manual. Analytical Software, P.O.Box 12185, Tallahassee, FL 32317-2185, USA
Blackshaw JK, Blackshaw AW (1994) Heat stress in cattle and the effect of shade on production and behaviour. Aust J Exp Agric 34:285–295. doi:10.1071/EA9940285
Beatty DT, Barnes A, Taylor E, Pethick D, McCarthy M, Maloney SK (2006) Physiological responses of Bos taurus and Bos indicus cattle to prolonged, continuous heat and humidity. J Anim Sci 84:972–985
Bray DR, Bucklin RA, Shearer JK, Montoya R, Giesy R (1997) Reduction of environmental stress in adult and young dairy cattle in hot, humid climates. Proceedings of the Fifth International Symposium, Bloomington, Minnesota, May 29–31, Vol I, pp 672–679
Broucek J, Kovalcik K, Gajdosik D (1987) The effect of extreme ambient temperatures on the haematological and biochemical parameters of heifers. Vet Med 32:259–268
Broucek J, Novak L, Kovalcik K (1988) The effect of lower temperatures on growth and health condition calves housed in outdoor hutches and calf barn. Czech J Anim Sci 33:27–36
Broucek J, Kovalcik K (1989) Einfluss der übermässigen künstlichen UV - Strahlung auf die Messgrossen des Blutbildes und auf die Phagozytose bei Kälbern. Dtsch Tierarztl Wschr 96:318–320
Broucek J, Kovalcik K, Letkovicova M, Novak L (1990a) Evaluation of ethological parameters in calves kept in outdoor hutches. Czech J Anim Sci 35:301–310
Broucek J, Kovalcik K, Novak L (1990b) Evaluation of different types of hutches for calves on the basis of the ethological studies. Polnohospodarstvo 36:543–552
Broucek J, Arave CW, Kisac P, Mihina S, Flak P, Uhrincat M, Hanus A (2006) Effects of some management factors on milk production in first-calf heifers. Asian-australas J Anim Sci 19:672–678
Brown-Brandl TM, Nienaber JA, Eigenberg RA, Freetly HC, Hahn GL (2003) Thermoregulatory responses of feeder cattle. J Therm Biol 28:149–157. doi:10.1016/S0306-4565(02)00052-9
Brown-Brandl TM, Eigenberg RA, Nienaber JA, Hahn GL (2004) Indicators of heat stress in shaded and non-shaded feedlot cattle. ASAE/CSAE Annual International Meeting, Ottawa, Ontario, Canada, Paper No 044037
Carter BH, Friend TH, Matis JH, Sawyer JE, Tomaszewski MA (2008) Effect of reflective insulation on calves in polyethylene hutches. J Anim Sci Vol 86, E-Suppl 2/J Dairy Sci Vol 91. E-Suppl 1:327.
Coleman DA, Moss BR, McCaskey TA (1996) Supplemental shade for dairy calves reared in commercial calf hutches in a southern climate. J Dairy Sci 79:2038–2043
Davis CL, Drackley JK (1998) The development, nutrition, and management of the young calf. Iowa State University Press, Ames, Iowa
Du Prezz JH, Giesecke WH, Hattingh PJ (1990) Heat stress in dairy cattle and other livestock under southern African conditions. III. Monthly temperature-humidity index mean values and their significance in the performance of dairy cattle. Onderstepoort J Vet Res 57:243–248
Hahn GL, Mader TL (1997) Heat waves in relation to thermoregulation, feeding behavior and mortality of feedlot cattle. Proceedings of the Fifth International Symposium, Bloomington, Minnesota, May 29–31, Vol I, pp 563–571
Hoshiba S, Sato Y, Isobe S, Sone A, Okamoto M, Dohkoshi J (1985) Thermal environment in calf hutches. Japn J Livest Manage 20:101–107
Hoshiba S, Isobe S, Sato Y, Dohkoshi J, Sone A, Okamoto M (1988) Behavior of calves housed in calf hutches in summer of Hokkaido as influenced by the meteorological environment. Japn J Livest Manage 23:73–80
Hubbard KG, Stooksbury DE, Hahn GL, Mader TL (1999) A climatologic perspective on feedlot cattle performance and mortality related to the temperature-humidity index. J Prod Agric 12:650–653
Huber JT (1996) Amelioration of heat stress in dairy cattle. In: Philips CJC (ed) Progress in dairy Science. CAB International, Oxon, UK, pp 211–243
Illek J, Kudrna V, Matejicek M, Novak P, Slavik P (2007) Heat stress in dairy cows – health, production and reproduction. Our Herd Prague 67:63–65
Jagos P, Illek J, Doubek J (1985) Metabolic profile in beef bulls under conditions of industrial technologies. Acta Vet (Brno) 54:41–51
Johnson HD (1987) Bioclimate effects on growth, reproduction and milk production. In: Johnson HD (ed) Bioclimatology and the adaptation of livestock. World Animal Science, B5. Elsevier, Amsterdam, The Nederlands, pp 35–57
Larson LL, Owen FG, Albright JL (1977) Guidelines toward more uniformity in measuring and reporting calf experimental data. J Dairy Sci 60:989–991
Lacetera N, Bernabucci U, Scalia D, Ronchi B, Kuzminsky G, Nardone A (2005) Lymphocyte functions in dairy cows under hot environment. Int J Biometeorol 50:105–110. doi:10.1007/s00484-005-0273-3
Mader TL, Davis MS (2004) Effect of management strategies on reducing heat stress of feedlot cattle: feed and water intake. J Anim Sci 82:3077–3087
Mader TL, Dahlquist JM, Hahn GL, Gaughan JB (1999) Shade and wind barrier effects on summertime feedlot cattle performance. J Anim Sci 77:2065–2072
Mader TL, Davis MS, Brown-Brandl TM (2006) Environmental factors influencing heat stress in feedlot cattle. J Anim Sci 84:712–719
May J, Mannoiu I, Donta C (1977) Untersuchungen über die Wärmebelastung beim Kalb. Zbl Veter Med Reihe A 24:153–159
Meyer DJ, Harvey JW (2004) Veterinary laboratory medicine: iInterpretation and diagnosis, 3rd edn. Saunders, St. Louis
Mitloehner FM, Galyean ML, McGlone JJ (2002) Shade effects on performance, carcass traits, physiology, and behavior of heat-stressed feedlot heifers. J Anim Sci 80:2043–2050
Mohri M, Sharifi K, Eidi S (2007) Hematology and serum biochemistry of Holstein dairy calves: age related changes and comparison with blood composition in adults. Res Vet Sci 83:30–39. doi:10.1016/j.rvsc.2006.10.017
Nardone A, Lacetera N, Bernabucci U, Ronchi B (1997) Composition of colostrum from dairy heifers exposed to high air temperatures during late pregnancy and the early postpartum period. J Dairy Sci 80:838–844
Nardone A, Ronchi B, Lacetera N, Bernabucci U (2006) Climatic effects on productive traits in livestock. Vet Res Commun 30(Suppl. 1):75–81. doi:10.1007/s11259-006-0016-x
Nienaber JA, Hahn GL, Eigenberg RA (1999) Quantifying livestock responses for heat stress management: a review. Int J Biometeorol 42:183–188. doi:10.1007/s004840050103
Robertshaw D (2006) Mechanisms for the control of respiratory evaporative heat loss in panting animals. J Appl Physiol 101:664–668. doi:10.1152/japplphysiol.01380.2005
Schäffer D, von Borell E (2008) Kritische Kontrollpunkte (CCP) in der Außenhaltung von Kälbern. Züchtungskunde 80:291–302
Seifi HA, Mohri M, Shoorei E, Farzaneh N (2006) Using haematological and serum biochemical findings as prognostic indicators in calf diarrhoea. Comp Clin Pathol 15:143–147. doi:10.1007/s00580-006-0620-8
Singh SP, Newton WM (1978) Acclimation of young calves to high temperatures: physiologic responses. Am J Vet Res 39:795–797
Soch M, Novak P, Kratochvil P, Travnicek J (1999) Losses of water vapour in the relation to the stable climate factors in calves. Acta Hortic Regiotecturae Nitra Slovakia 2:179–181
Soch M (2005) Effect of environment on selected indices of cattle welfare. University of South Bohemia, Ceske Budejovice
Spain JN, Spiers DE (1994) Physiologival responses of young calves housed in hutches with or without supplemental shade during summer. Dairy Systems for the 21st Century. Proceedings of the Third International Dairy Housing Conference, 2–5 February 1994, Orlando, Florida, pp 781–789
Spain JN, Spiers DE (1996) Effects of supplemental shade of calves to heat challenge in on thermoregulatory response a hutch environment. J Dairy Sci 79:639–646
West JW (2003) Effects of heat-stress on production in dairy cattle. J Dairy Sci 86:2131–2144
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Broucek, J., Kisac, P. & Uhrincat, M. Effect of hot temperatures on the hematological parameters, health and performance of calves. Int J Biometeorol 53, 201–208 (2009). https://doi.org/10.1007/s00484-008-0204-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00484-008-0204-1