
Abstract
Some of the factors that could influence changes in phenology of the song thrush (Turdus philomelos) during spring migration have been analyzed in relation to the timing of their return to the south-east Baltic region over the last 40 years. These include wind direction and velocity, temperature and precipitation in the wintering areas and along the migratory route. In March, a significant correlation was found between the timing of passage in the Baltic region and both the air temperatures in wintering area and winds over the migratory route. In April, when mass migration of thrushes occurs, the timing of passage was correlated with temperature, winds and, partly, precipitation over the route of migration. In this month, the frequency of tailwinds increased significantly along the route of migration from south-west France to the Baltic region over the last 40 years. The tailwind frequency was correlated with the beginning, middle and end of spring passage, accounting for 51% of the variance of median dates of the passage. The higher ambient temperatures over the migratory route explain nearly 20% of the variance in the timing of passage. Our results suggest that the advance in the timing of spring migration is primarily due to (1) the increased frequency of tailwinds favorable for migratory flights over the migratory route, and (2) the earlier emergence of the conditions enhancing migration due to higher spring temperatures in the winter quarters and on the migratory route.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
A large amount of information concerning global climate changes and their impact on ecosystems has recently been published (Post et al. 2001; Møller 2002; Nott et al. 2002; Stenseth et al. 2002; Walther et al. 2002; Hüppop and Hüppop 2003). A positive NAO (North Atlantic Oscillation) index trend reflects an intensification of redistribution of the atmospheric mass across the North Atlantic (Hurrell et al. 2001; Nott et al. 2002) which, in turn, causes an increase in the frequency of warm and wet SW and W winds from the Atlantic across northern and central Europe in late winter and early spring (Hurrell 1995; Visbeck et al. 2001). Winter and early spring air temperatures in Europe have increased mainly because of this phenomenon (Sparks and Carey 1995; Post et al. 2001).
Climate changes are reflected in changes in bird phenology. Many authors have revealed a close relationship between higher spring air temperatures and the earlier arrival dates of both short- and long-distance migrants (Mason 1995; Sokolov et al. 1998; Sparks 1999; Barrett 2002; Sparks et al. 2003). This has been confirmed by monitoring observational and trapping records of birds in Europe since the middle of the twentieth century (Sokolov et al. 1998; Tryjanowski et al. 2002; Hubálek 2003).
Wind condition correlated with the temperature regime in Europe in early spring may also strongly influence the arrival of birds. For many species of short- and long-distance migrants passing in spring across the Baltic region, a significant relationship has been established not only with air temperature but also with precipitation and wind characteristics (Žalakevičius 2005).
Wind is believed to be the major meteorological factor governing migratory flight activity of birds (Lack 1960b; Richardson 1978, 1990; Alerstam 1990). Visual and radar observations have shown that the numbers of passerine nocturnal migrants in particular are strongly correlated with the occurrence of tailwinds which are favorable from energetic and orientational considerations (Alerstam 1976, 1990; Alerstam and Lindström 1990; Åkesson and Hedenström 2000). Migrants select for favorable wind conditions both for initiating departure and for flight (Liechti 2006) preferring nights with tailwinds, or choosing an altitude range with appropriate following winds (Liechti and Bruderer 1998). Those birds that have tailwinds generally migrate for longer distances per night than birds that have headwinds or strong side winds (Cochran and Wikelski 2005). In some years, the passage of thrushes may occur during a few “good” nights with tailwinds (Alerstam 1990).
The song thrush (Turdus philomelos) is a typical and numerous nocturnal middle-distance migrant overwintering in SW Europe. The study of the phenology of this species during 40 years of monitoring trapping shows that the following occurs:
-
1.
The median date of spring passage in the south-east Baltic region (SE Baltic) has significantly advanced by 10 days (0.25 ± 0.16 days per year, P = 0.009).
-
2.
The dates of the commencement of spring passage are correlated with the March NAO index values.
-
3.
There is no significant relationship between the timing of spring passage and either March or April air temperatures in the SE Baltic region (Sinelschikova and Sokolov 2004).
As a consequence, it is suggested that the significant advance in timing of spring migration in birds may not be due entirely to the higher spring temperatures per se. Another factor causing faster migration may be the changing wind situation in Europe over the migratory route, due to the increased number of nights with tail winds and/or to increased wind speed.
In the present study, we investigated (1) the change of wind situation in Europe in spring (increase in frequency of tailwind for birds wintering in SW Europe and/or its speed) over the last 40 years, and (2) the specific advantages that wind change en route from SW Europe to the SE Baltic give to the thrushes with respect to temperature and precipitation.
Material and methods
Trapping data
Data were analyzed for 1,324 song thrushes captured in the spring by Rybachy-type traps at Fringilla field station (55°05′N, 20°44′E) situated on the Courish Spit on the Baltic Sea coast, for the years 1961 to 2003. Each year, two stationary traps were operated simultaneously 24 h a day from the end of March until November. The main spring migratory period of the song thrush recorded by moon-watching occurs, on average, between 28 March and 29 April (Bolshakov et al. 2002). The years when the trapping began later than 29 March or the number of thrushes caught in spring was fewer than 20 birds (n = 7 years) were excluded from the analysis. In the course of 40 years of trapping no trends were found in the annual total of the migratory population of the song thrush (F 1,38 = 2.55, P = 0.120).
The analysis period of spring migration was from the onset of trapping in the end of March to 20 May. All thrushes trapped until 1 May were used. Birds caught during 2 h after sunrise between 1 and 20 May were also included because nocturnal migratory flights recorded by calls may continue until the end of May (Bolshakov et al. 2002). Birds caught in May during the day were excluded as local ones had probably begun nesting. The calendar dates for capturing 5, 20, 50, 80 and 95% of all birds were taken as temporal metrics for analyzing spring migration. In some years, nocturnal passage of thrushes may be recorded by calls as early as 10 March (Bolshakov et al. 2002). Therefore, the capture dates of 5% of birds do not entirely reflect the onset of migration, but they do indicate trends (Sinelschikova and Sokolov 2004).
Recovery data
The influence of climatic factors on the timing of the spring passage of the song thrushes in the SW Baltic area was considered in relation to the distribution of the birds within their wintering grounds and on their migratory route. Long-distance recovery data was extracted from the database of the Biological Station “Rybachy” and the Bird Ringing Centre (Moscow), which also provided the recovery data of the birds that had been ringed in the neighboring SE Baltic countries of Latvia and Lithuania in 1961–1998. A total of 134 long-distance recoveries between February and May in 1962–2002 was included.
Wind, temperature, precipitation
The meteorological situation was assessed both in the wintering areas and along the migratory route for the populations of the song thrushes passing through the Baltic. This is the wide area extending from S Spain, Portugal, Mediterranean islands, S Italy to the SE Baltic (Payevsky et al. 2004).
The weather data for 17 sites in Europe from February to May were downloaded from the Internet: wind - http://www.dss.ucar.edu/datasets/ds195.1; temperature - http://www.ncdc.noaa.gov/; and precipitation - http://www.cru.uea.ac.uk/~mikeh/datasets/global/. All parameters were presented as 5 × 5 degree grid. The coordinates for wind data and precipitation coincided, but for temperature they differed by 2.5 degrees relative to the winds or precipitation. To match the coordinates we have interpolated the temperature grid data on the same coordinates as wind and precipitation. Data on speed and direction of wind corresponding to 850 Mb (1500 m asl) at midnight 00Z (Greenwich) were used. The studies of nocturnal migration of European birds have shown that in spring the most intensive passage occurs around midnight (Alerstam 1976), and although passerine birds can fly at heights up to 4 km agl about 90% of them fly within the height range of 100–2,000 m agl (Liechti et al. 1995; Bolshakov et al. 2002; Liechti 2006, and references therein).
The wind most appropriate for assisting migratory flight was investigated. The mean azimuth of winter recoveries for the song thrushes banded on the Courish Spit was 223° (Payevsky et al. 2004). A previous study by moon-watching showed that the general direction of spring nocturnal migration of thrushes in the SE Baltic is 044° (Bolshakov et al. 2002). Assuming the fly direction the strict tailwind appears to be 224°. In spring, some 78% of thrushes migrate in winds with a tailwind component. More than half of birds migrate when the wind does not deviate from the strict tailwind by more than 39° (Baushev and Sinelschikova 2007). The most intensive nocturnal passage in spring was observed when the wind deviated from the strict tailwind by 1–2° and wind speed varied between 2–12 m/s (Sinelschikova et al., in preparation).
The frequency of tailwinds (i.e. the proportion of nights with tailwinds as % per month) and their mean speed were calculated for each of the 17 points throughout Europe, and for each month.
The wind was considered to be a tailwind if it fits the following assumption. When one expands the vector of wind speed on the general migratory direction and the orthogonal the values of the corresponding projections are V×Cosφ and V×Sinφ, where V is the wind speed and φ the angle between wind direction and general direction of migration. The wind becomes a constituent of the ground speed of the bird, if inequality V×Sinφ ≤ V×Cosφ is performed. This inequality is performed when 0 ≤ tg φ ≤ 1, hence 0° ≤ φ ≤ 45°. Therefore, the SW winds within 90° sector ( 224 ± 45°) were accepted as the tailwinds.
To reveal long-term trends in wind condition a regression analysis was applied. Correlation analysis was used to find the relationships between the timing of migration and temperature regime, precipitation and tailwind frequency. To confirm that the resulting relationships did not just reflect the trend correlation we additionally applied correlation of regression residuals. We used multiple linear regression models to explain the variation in spring passage of thrushes in the Baltic in terms of weather factors in their winter quarters and on their migratory route.
Results
The distribution of the song thrushes in their wintering area and on their migratory route
Song thrushes arrive at their most distant wintering areas in the Mediterranean from January to the first half of February, as shown by banding recoveries (Payevsky et al. 2004).
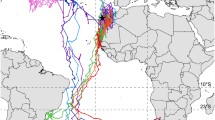
In March, the birds begin their return journey in a northeasterly direction when distributed within southwest and central Europe (Fig. 1). For the purpose of further analysis we defined the line between wintering and migratory areas. It lies from central Italy to SW France. Although in some years early song thrushes flying at night are recorded at the Courish Spit by the second 10-day period of March (Bolshakov et al. 2002), recoveries at that time show a distribution between the southern parts of the wintering area to the Baltic part of their migratory route.

Distribution of the song thrushes (Turdus philomelos) banded in the SE Baltic and found in their winter quarters and migratory route. Symbols: birds found in, ● February, ▲ March, ● April, ■ May; 0 ringing region; ↗ general direction of spring migration
In April, birds have left the southern parts of their wintering grounds and are distributed mainly along their migratory route. The median and mean date for spring passage of the song thrush at the Courish Spit is 17 April. This date is the same for both diurnal trapping at stopovers (Sinelschikova and Sokolov 2004) and nocturnal moon-watching observation (Bolshakov et al. 2002).
The spring passage at the Courish Spit ends, on average, by mid-May, although the last birds migrating at night may be recorded by calls until the beginning of June (Bolshakov et al. 2002). May recoveries are mainly concentrated north of the Courish Spit.
Wind situation in Europe
The monthly trend of wind changes in Europe over the last 40 years was evaluated for assisting spring migratory flight of the song thrush wintering in SW Europe; that is in SW sector 224° ± 45°.
In February, in the winter quarters of the song thrush in Spain, N Italy and the islands in the Mediterranean Sea, it was shown by regression analysis that the frequency of tailwinds reduced significantly over the period under consideration (Fig. 2). The mean speed of the tailwinds did not change and even decreased in E Spain. In the other parts of their wintering area the wind situation did not change.

Trends of the wind situation in Europe in the sector of SW tailwinds (224 ± 45°) during 1961–2002. Results of regression analysis (the values of coefficient F). Each pair of columns is referred to the intersection of the 5° grid lines. In each pair of columns: first column - frequency of the tailwinds, second column - mean speed of the tailwinds. 1st line - February, 2nd line - March, 3rd line - April, 4th line - May. Bold - significant values of F, where: *P < 0.05, **P < 0.005. (-) F- when B < 0 (the frequency of tailwinds or their speed decreased)
In March, the situation on the wintering grounds was very similar to February. No change was found in tailwind frequency along the migratory route except for a significant increase in the tailwind speed observed only in Germany.
In April, when the mass migration of thrushes occurs in the Baltic, the frequency of tailwinds increased significantly along the whole western and central part of the route of migration from SW France to the S Baltic (Fig. 2). However, the mean speed of the tailwinds over the migratory route did not change in April over the 40 years.
In May, the wind condition in Europe did not change at any of the 17 sites under consideration.
The relationship between the timing of spring migration and wind
Evaluation of the possible impact of wind conditions in Europe on the timing of spring migration of thrushes in the SE Baltic region gave the following results:
No relationship was found between the timing of migration and February winds. All the values of Spearman correlation coefficients corresponded to P > 0.05. The frequency of tailwinds in March did not increase in Europe, nevertheless a significant negative correlation was revealed between the winds over the route of migration and in NW areas of wintering grounds on the one hand, and the timing of the first half and middle of migration in the Baltic (dates of passage of 20% and 50%) on the other (Fig. 3a). Over the migratory route, a significant correlation was found between the timing of migration and both frequency of tailwind and its speed over the vast extent of the north-western part of the route along the coasts of the North and the Baltic Seas. When we ranged the frequency of tailwind by its speed in 5 m/s steps, it became obvious that the strongest correlation existed with tailwinds in the range of 5–10 m/s (r from −0.54 to −0.51, P from 0.006 to 0.009). In the range of 0–5 m/s, correlation was found in only two sites en route (P = 0.029 and 0.043). In the other 5 sites, it was close to significant (P between 0.062 and 0.097). Practically the same was found for the tailwind range of 10–15 m/s (P between 0.057 and 0.123). It is possible that the speed of tailwinds along the eastern part of the migratory routes influences the timing of the passage in the Baltic region. Wind conditions in April may have the strongest impact on the timing of migration and consequently on the speed of migration since mass migration of thrushes in general occurs in this month. In April, almost all the periods of passage of the song thrushes (20, 50, 80 and 95%) in the Baltic are significantly correlated with the frequency of tailwinds over the western part of the migratory route and NW of the wintering areas, where, judging by banding recoveries, some thrushes may still remain (Fig. 3b). The highest values of the coefficient of Spearman correlation (r from −0.65 to −0.71, P < 0.001) were for the passage of 50 and 80% of birds over the stretch from France to the S Baltic. The strongest effect of the tailwind velocity on the timing of the first half of passage occurs along the western part of the migratory route.
In April, as in March, the frequency of tailwinds increased predominantly in the speed range of 10–15 m/s (F 1,32 between 8.99-6.04, P 0.006-0.03). But the most significant correlation was found with tailwinds within the range of 5–10 m/s (r from −0.63 to −0.79, P < 0.001). Correlation with the frequency of tailwinds within the ranges of 0–5 and 10–15 m/s was also significant in most points (r from −0.57 to −0.44, P < 0.05). Strong winds over 20 m/s which might impede the migration (Bolshakov et al. 2002) occurred rarely and not every year. These results show that more often tailwinds occur and that the higher their corresponding velocity along the western part of the migratory route the earlier spring migration of thrushes occurs in the SE Baltic overall.
A significant negative correlation was found between the end of migration (dates of 80 and 95%) and the frequency of tailwinds only in the S and SE Baltic in May.
The correlation analysis of regression residuals have not changed the presented results. The revealed relationships were not a result of a coincidence of trends but of the congruence of variations. This confirms that the trends of times and variations of dates of the spring passage of the song thrush in the SE Baltic over the last 40 years are significantly related to the general trends in wind situation and its variations in Europe.

Correlation between the dates of capture of 5, 20, 50, 80 and 95% of the song thrushes in the SE Baltic and meteorological factors in Europe (Spearman’s correlation analysis). a March, b April. Each array of signs is referred to the intersection of the 5° grid lines. In each array: 1st line - correlation of meteorological factors with 5% of captured birds, 2nd line - correlation of meteorological factors with 20% of captured birds, 3rd line - correlation of meteorological factors with 50% of captured birds, 4th line - correlation of meteorological factors with 80% of captured birds, 5th line - correlation of meteorological factors with 95% of captured birds. Symbols: open - the values are nonsignificant, filled - significant; □ - correlation of dates of capture with the frequency of the SW tailwinds, ■ - correlation is significant with r < 0, P < 0.05; ✼ - correlation of dates of capture with the mean speed of the SW tailwinds, ✼ - correlation is significant with r < 0, P < 0.05. ○ - correlation of dates of capture with monthly air temperature, ● - correlation is significant with r < 0, P < 0.05., △ - correlation of dates of capture with monthly precipitation (r > 0), ▽ - correlation of dates of capture with monthly precipitation (r < 0), ▼ - correlation is significant with ( r < 0, P < 0.05)
The relationship between the timing of spring migration and temperature
In February, no influence of monthly ambient temperature on the timing of passage of the song thrush in the Baltic was found.
In March, there was a significant negative correlation found between the temperatures in western and southern wintering areas (Spain, S France, S Italy and islands of Mediterranean Sea) and timing of spring migration in the Baltic (Fig. 3a). The March temperature influences mainly the middle of the passage. The March temperatures in most parts of the wintering area were correlated with winds, but over the migratory route there was no significant relationship between the March temperature and timing of migration.
In April, a significant correlation between passage and European temperatures was found at only three sites along the migratory route. At these sites, the relationship with wind was also established (Fig. 3b). Note, that unlike the end of winter or early spring when the temperature regime in Europe is strongly governed by the incursion of the Atlantic cyclones, in April, sun radiation intensity starts to play an important role (Weisberg 1976; Barinova 2002). It seems to be the main reason why along most parts of the migratory route a correlation between wind and temperature in April was absent. Only at one site in Germany was the temperature correlated with tailwind frequency in April.
In May, no significant correlation was found between the temperature over the route and the dates of the end of spring migration.
Relationship between timing of spring migration and precipitation
In February, the monthly precipitation rates have not changed over 38 years with available data (1961–1998) at any examined sites in Europe. According to regression analysis, no value of F exceeded the means when P > 0.170. A significant correlation between the timing of passage and precipitation was not established. In March, in the wintering areas (Spain and NW Mediterranean), the monthly precipitation rate decreased significantly (F from 13.14 to 4.21, with P from 0.001 to 0.047). In the other regions of Western Europe, there were no significant changes. Within the wintering area, the precipitation rates were positively correlated with the timing of spring passage, whereas in the S Baltic the correlation was negative (Fig. 3a). In April, monthly precipitation rate did not change in any site in Western Europe. No value of F was significant (P > 0.220). No correlation was found with the timing of migration of thrushes (Fig. 3b). In May, no trends in precipitation rate were found along the migratory route and no influence on the timing of migration was apparent.
Models
In order to assess the quantitative range of relationships between the annual song thrush monitoring data in the SE Baltic and climatic factors in the wintering areas and en route, and consequently to explain the shift in bird arrival dates by climate change, we applied multiple linear regression models. Similarity of the type of climate changes within the Mediterranean (wintering area) as well as in the NW part of Western Europe (migratory route) permitted grouping the sites and averaging data. The sites over the SE part of the Alps and Western Carpathian Mountains seemed to be of less significance, since the density of birds flux over the mountain area was about 3-fold less than to the north of Alps (Liechti et al. 1996), and no correlation was found between the temporal metrics of the migration and winds or temperature there. For each group of sites where the climate data showed significant correlation with the timing of spring passage for each year, the mean values of temperature and frequency and speed of tailwinds were calculated. The monthly climate data relating to each grouping of sites were also regressed with each other to assess their independence. After this procedure precipitation was excluded due to its correlation with the temperature or wind. This resulted in 6 independent variables (judged by 5% significance level) which were included in the general model of multiple linear regression as the predictors:
-
1.
Mean March temperature in the wintering area (4 sites averaged),
-
2.
Mean April temperature over the migratory route (3 sites),
-
3.
Frequency of the tailwinds in March over the migratory route (4 sites),
-
4.
Frequency of tailwinds in April over the migratory route (4 sites),
-
5.
Mean speed of tail winds in March over the migratory route (4 sites),
-
6.
Mean speed of tail winds in April over the migratory route (4sites).
The median (Me) date of the spring passage at the Courish Spit was taken as a dependent variable. The previous study showed that the Me date was significantly correlated with the dates of beginning and end of the migration of the thrushes (Sinelschikova and Sokolov 2004).
In order to make the variables dimensionless they have been standardized. Thus, the general model was as follows:
where Δt′=(t−tmean)/SD(t), Δf’=(f−fmean)/SD(f),), Δv′=(v−vmean)/SD(v), t = temperature, f = frequency of tailwind and v = tailwind speed.
In order to reveal which predictors have the greatest effect (Chatterjee and Hadi 1986) and to estimate the amount of explained variance we examined 6 models with each of the independent predictors (Table 1).
Our results, as shown in Table 1, reveal that more than 50% of variance of median dates of spring passage could be explained by the frequency of the tailwinds in April along the migratory route. Additionally, we tested two models with two predictors: (1) frequency of tailwinds in March and April over the route of migration and (2) temperature in March on the wintering grounds and in April over the route. The model including the two predictors—frequency of tailwinds in March and April—explains nearly 60% of the variance. The temperature regime both in the wintering areas and en route governs the timing of spring migration, as most researchers have concluded, but explains only about 30% of variance.
Discussion
Climate change is likely to have caused the shift in the timing of spring migration in many species of long-distance and especially short-distance migrant birds. The main questions of whether birds leave their wintering areas earlier in recent decades and whether this fact is the cause of their advance in the timing of migration or acceleration of migration occurs over the migratory route is poorly investigated. The study of the wintering behavior in common European bird species has shown that, since the 1960s, some of them have moved their wintering areas further north (Clark et al. 2005) where a different photoperiod and other environmental factors play a role in modifying departure schedules (Berthold 1993). It is possible that the spring departure schedule may vary in birds. It was shown that in the white stork (Ciconia ciconia), even in the same individual bird marked by a transmitter, the beginning of spring migration might vary within a month in different years (Kaatz 2004). Other investigations suggest that it is unlikely that winter departure time would change to adjust to short-term changes in climate on northern breeding areas (Marra et al. 2005, and references therein). Delayed arrivals in Spain of some species of trans-Saharan migrants, which arrive in northern European regions earlier than before, may be the evidence of acceleration along the European part of their migratory route (Gordo et al. 2005). Unfortunately, in this study, lack of recovery data did not permit us to undertake spatial analysis of distance and pattern of distribution of the song thrushes in their winter quarters during the very beginning of migration in the years of their late and early arrival at the Courish Spit. Thus, it is still unknown whether thrushes shifted their departure schedule in the recent decades of early arrival or not (Sinelschikova and Sokolov 2004). It should be noted that, in February and March, during the beginning of spring movements, the frequency of tailwinds has significantly declined in the Mediterranean area over the last 40 years. Therefore, wind conditions in this region in early spring were not more favorable for earlier migratory nocturnal flights.
We have revealed relationships of arrival of thrushes with temperature, precipitation and wind condition. Significant correlation was found between the timing of spring passage in the Baltic and both temperature and precipitation on the winter grounds in March. It has been shown that in the Mediterranean area the average annual temperatures have increased by 1.4°C over the period of 1952–2000, and in recent years leaves unfold, on average, 16 days earlier, and plants flower, on average, 6 days earlier than in 1952 (Penuelas et al. 2002). According to the results of our study the Mediterranean temperature in March explains about 20% of variance of median dates of spring passage. Therefore, the warmer and drier an early spring is in this region, the earlier the most birds migrate to the Baltic. The overwhelming majority of research workers explain the earlier migrant arrival, including by the song thrush, by the increase in monthly air temperature (Mason 1995; Sparks and Mason 2001; Forchhammer et al. 2002). Some of them suggest that the higher spring temperature encourages food growth (Sokolov 2000; Both and Visser 2001; Forchhammer et al. 2002; Sparks et al. 2003). Ambient temperature controls food abundance and availability during the pre-migratory period providing the physiological support for migration. A shift in migratory schedule may be due to climate change allowing faster migration through improving food supply at stopovers. Our results show that the ambient temperature in April over the migratory route explains nearly 20% of the variance in the timing of passage of the song thrushes.
Climate changes over the European part of the migratory route of many species concern not only the temperature regime. Forchhammer et al. (2002) analyzed and contrasted the effects of climate (NAO) and temporal dependence on the long-term (1928–1977) dynamics of springtime arrival in short- and long-distance migratory bird species at breeding locations across Norway. They showed that the above effect on short-distance migrants may not be improved food availability alone, but weather conditions during their northward spring migration may also be involved. Increasing NAO index reflects increasing strength and frequency of warm westerly winds from the Atlantic which, in turn, cause high winter and spring temperatures in Europe (Visbeck et al. 2001). Consequently, in Western Europe not only the air temperature (the increase of which is believed to be the main cause of the phenological change in birds) but also the frequency of westerly and south-westerly tail winds has increased in spring in recent decades (Sparks et al. 2002). Our analysis is absolutely in agreement with this conclusion.
Our investigation over the 40-year period from 1961 to 2003 has shown that, in March and especially in April, during mass migration of thrushes a significant relationship exists between the timing of migration and the frequency and speed of the tailwinds in the areas along the migratory route from France to the Baltic countries. The strongest relationship is revealed in the north-western part of the migratory route where along the coasts of the North Sea and the Baltic Sea a significant increase in the frequency of SW tailwinds was recorded in April. In March, the winds over the route were mainly correlated with the first half and the middle of spring passage and explain nearly 20% of the variance of the median of spring migration. In April, the tailwind frequency in speed range of 1–15 m/s was correlated with the beginning, middle and end of spring passage over the western and central part of the migratory route, accounting for 51% of the variance of spring passage in the song thrush in the Baltic region. Our results show that the increase in frequency of tailwinds in April over the migratory route provided the major influence on the advance of spring migration of this species in Western Europe over this 40-year period. The advantages of tailwinds during migration are mentioned above. During spring migration, birds take account of the winds more selectively than in autumn (Sinelschikova et al. 2003). The wind conditions evident in Europe along the migratory route in recent decades provide a wider opportunity for birds to select nights appropriate for migration.
Our results also confirm the previous proposition (Sinelschikova and Sokolov 2004) that a significant advance in the timing of spring migration recorded in the last 40 years is primarily due to (1) the increased frequency of tailwinds favorable for migratory flights over the western and central parts of the migratory route, and also to (2) the earlier emergence of the conditions enhancing migration due to higher spring temperatures (favoring food and fuel deposition) in the winter quarters and on the migratory route.
References
Åkesson S, Hedenström A (2000) Wind selectivity of migratory flight departures in birds. Behav Ecol Sociobiol 47:140–144
Alerstam T (1976) Nocturnal migration of thrushes (Turdus spp.) in southern Sweden. Oikos 27:457–475
Alerstam T (1990) Bird migration. Cambridge University Press, Cambridge
Alerstam T, Lindström Å (1990) In: Gwinner E (ed) Optimal bird migration: the relative importance of time, energy and safety. Springer, Berlin Heidelberg New York, pp 331–359
Barinova GM (2002) Kaliningrad region. Climate. FGUIPP ”Iantarny Skaz” (in Russian)
Barrett RT (2002) The phenology of spring bird migration to north Norway. Bird Study 49:270–277
Baushev AN, Sinelschikova A (2007) On a probabilistic model for the numerical estimation of nocturnal migration of birds. Math Biosci 205:44–58
Berthold P (1993) Bird migration. A general survey. Oxford University Press, Oxford
Bolshakov C, Žalakevičius M, Švaas S (2002) Nocturnal migration of thrushes in the Eastern Baltic region. Vilnius
Both C, Visser ME (2001) Adjustment to climate change is constrained by arrival date in long-distance migrant birds. Nature 411:296–298
Chatterjee S, Hadi AS (1986) Influental Observations, High Leverage Points, and Others in Linear Regression. Stat Sci 379–416
Clark J, Soutullo A, Gill J, Robinson R, Reynolds J (2005) Climate change and shifts in winter distribution of European breeding birds. ESF Optimality in bird migration, final conference “Migration in the life-history of birds”. 16–20 Feb 2005, Wilhelmshaven, Germany, p 10
Cochran WW, Wikelski M (2005) Individual migratory tactics of new world Catharus Thrushes. In: Greenberg R, Marra PP (eds) Birds of two worlds: the ecology and evolution of migration. John Hopkins University Press, Baltimore, pp 274–289
Forchhammer MC, Post E, Stenseth NC (2002) North Atlantic Oscillation timing of long-and short-distance migration. J Anim Ecol 71:1002–1014
Gordo O, Brotons L, Ferrer X, Comas P (2005) Do changes in climate patterns in wintering areas affect the timing of the spring arrival of trans-Saharan migrant birds? Global Change Biol 11:12–21
Hubálek Z (2003) Spring migration of birds in relation to North Atlantic Oscillation. Folia Zool 52:287–298
Hüppop O, Hüppop K (2003) North Atlantic Oscillation and timing of spring migration in birds. Proc R Soc Lond B 270:233–240
Hurrell JW (1995) Decadal trends in the North Atlantic Oscillation: regional temperatures and precipitation. Science 269:676–679
Hurrell JW, Kushnir Y, Visbeck M (2001) The North Atlantic Oscillation. Science 291:603–605
Kaatz M (2004) Der Zug des Weiβstorchs Ciconia ciconia auf der europäischen Ostroute über den Nahen Osten nach Afrika. Dissertation (Dr.agr.). In: Mit prinzenβchen unterwegs. Abenteuer, Wissenschaft und Storchensommer. Halle/Saale
Lack D (1960b) Migration across the North Sea studied by radar. Part 2. The spring departure 1956-59. Ibis 102:26–57
Liechti F (2006) Birds: blowin’ by the wind? J Ornithol 147:202–211
Liechti F, Bruderer B (1998) The relevance of wind for optimal migration theory. J Avian Biol 29:561–568
Liechti F, Bruderer B, Paproth H (1995) Quantification of nocturnal bird migration by moonwatching: comparison with radar and infrared observations. J Field Ornithol 66:457–468
Liechti F, Peter D, Lardelli R, Bruderer B (1996) Topography and wind affect the course of migration. Ornithol Beob 93:131–152
Marra P, Francis C, Mulvihill R, Moore F (2005) The influence of climate on the timing and rate of spring bird migration. Oecologia 142:307–315
Mason CF (1995) Long-term trends in the arrival dates of spring migrants. Bird Study 42:182–189
Møller AP (2002) North Atlantic Oscillation (NAO) effects of climate on the relative importance of first and second clutches in a migratory passerine bird. J Anim Ecol 71:201–210
Nott MP, Desante DF, Siegel RB, Pyle P (2002) Influences of the El Nińo/Southern Oscillation and the North Atlantic Oscillation on avian productivity in forests of the Pacific Northwest of North America. Global Ecol Biogeogr 11:333–342
Payevsky VA, Vysotsky VG, Shapoval AP (2004) Demography and spatial distribution during migrations of hunting thrushes. Zool Journ 83:342–354, (in Russian with resume in English)
Penuelas J, Filella I, Comas P (2002) Changed plant and animal life cycles from 1952 to 2000 in the Mediterranean region. Global Change Biol 8:531–544
Post E, Forchhammer MC, Stenseth NC, Callaghan TV (2001) The timing of life-history events in a changing climate. Proc R Soc Lond B 268:15–23
Richardson WJ (1978) Timing and amount of bird migration in relation to weather: a review. Oikos 30:224–272
Richardson WJ (1990) Timing of bird migration in relation to weather: update review. In: Gwinner E (ed) Bird migration. Springer, Berlin Heidelberg New York, pp 78–101
Sinelschikova A, Sokolov LV (2004) Long-term monitoring of the timing of migration in thrushes (Turdus philomelos, T. iliacus) in the Eastern Baltic. Avian Ecol Behav 12:11–30
Sinelschikova A, Bolshakov C, Bulyuk V (2003) Nocturnal migration of thrushes (Turdus spp.): numbers aloft and the wind. Die Vogelwarte, 4th Conf EOU: 100
Sokolov LV (2000) Spring ambient temperature as an important factor controlling timing of arrival, breeding, post-fledging dispersal and breeding success of Pied Flycatchers Ficedula hypoleuca in Eastern Baltic. Avian Ecol Behav 5:79–104
Sokolov LV, Markovets MY, Shapoval AP, Morozov YG (1998) Long-term trends in the timing of spring migration of passerines on the Courish Spit of the Baltic Sea. Avian Ecol Behav 1:1–21
Sparks TH (1999) Phenology and the changing pattern of bird migration in Britain. Int J Biometeorol 42:134–138
Sparks TH, Carey PD (1995) The responses of species to climate over two centuries: an analysis of the Marsham phenological record, 1736–1947. J Ecol 83:321–329
Sparks TH, Mason CF (2001) Dates of arrivals and departures of spring migrants taken from Essex Bird Reports 1950–1998. Essex Bird Reports 1999:154–164
Sparks TH, Roberts DR, Crick HQP (2002) What is the value of first arrival dates of spring migration in phenology? Long-term dynamics of bird and mammal populations and global climatic changes. Kazan, pp 39–46
Sparks TH, Crick HQP, Dunn PO, Sokolov LV (2003) Ch.3.6 Birds. In: Schwartz (ed) Phenology: an integrative environmental science, pp 421–436
Stenseth NC, Mysterud A, Ottersen G, Hurrell JW, Chan K-S, Lima M (2002) Ecological effects of climate fluctuations. Science 297:1292–1296
Tryjanowski P, Kuzniak S, Sparks T (2002) Earlier arrival of some farmland migrants in western Poland. Ibis 144:62–68
Visbeck MH, Hurrell JW, Polvani L, Cullen HM (2001) The North Atlantic Oscillation: Past, present and future. PNAS 98(23):12876–12877
Walther GR, Post E, Convey P, Menzel A, Parmesan C, Beebee TJC, Fromentin JM, Hoegh-Guldberg O, Bairlein F (2002) Ecological responses to recent climate change. Nature 416:389–395
Weisberg JS (1976) Meteorology of the Earth and its Weather. Houghton Mifflin
Žalakevičius M (2005) Spring arrival response in birds to climate change in Eastern Europe: a case study. ESF Optimality in bird migration, final conference “Migration in the life-history of birds”, Wilhelmshaven, Germany, p 11
Acknowledgements
This study was supported by OMPO (France) and the Russian Foundation for Basic Research (grant to Victor Bulyuk no. 04-04-49161 and grant to Leonid Sokolov no. 06-04-48774). The authors are grateful to their colleagues and ringers who have participated in the long-term trapping and ringing of thrushes on the Courish Spit. We thank Prof. Dr. Martin Griffiths for his helpful suggestions, Dr. Andrey Sharkov for MS improvement and two reviewers for their valuable comments.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Sinelschikova, A., Kosarev, V., Panov, I. et al. The influence of wind conditions in Europe on the advance in timing of the spring migration of the song thrush (Turdus philomelos) in the south-east Baltic region. Int J Biometeorol 51, 431–440 (2007). https://doi.org/10.1007/s00484-006-0077-0
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00484-006-0077-0