
Abstract
Key message
Multiple linear regression based on the crown diameter (the most explicative biometric variable for pollarded trees) and diameter at breast height indices produced the more parsimonious model with moderate accuracy to estimate leaf biomass used to feed livestock in strongly seasonal silvopastures.
Abstract
The aim of this study is to develop allometric equation to estimate leaf biomass of pollarded Lebanon oak trees, a species used as fodder tree in western Iran. A pollarded stand (6.6 ha in area including 560 Lebanon oak trees) was selected. Full callipering of all trees (collar diameter ≥5 cm) was conducted: collar diameter and diameter at breast height (DBH) of stem, tree height, trunk height, and crown diameters (CDs) were measured. At the time of pollarding, the fresh weight of leafy branches resulting from pollarding of each tree was measured. Finally, dry weight of leaf biomass for entire crown was determined. Biometric indices as predictor variables for estimating dry weight of leaf biomass were used. Multiple linear regression based on CD and stem DBH produced the more parsimonious model. The results indicated that natural logarithmic transformation of dry weight of leaf biomass improved the model’s goodness of fit. The moderate accuracy of the proposed allometric equation is explained by the scattered distribution of the trees and the strong modification of canopy structure through periodical pollarding. Leaf biomass estimate is used to evaluate the impact of pollarding on nutrient cycling and carbon sequestration in forest.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Biomass of different parts of standing trees, which is dependent on site characteristics and tree attributes, is difficult to predict (Lehtonen 2005). While there is abundant scientific literature on the quick estimation of timber and wood biomass (i.e., the world’s most common forest products), scientific knowledge for the estimation of leaf biomass is still poor. Since direct measurement of leaf biomass is impractical, leaf biomass estimation using non-destructive techniques requires development of allometric equations based on predictor variables such as biometric indices which can be easily measured. Accurate estimates of leaf biomass are essential because it indicates the production capacity of forest per unit area or time and can be used as an indicator of site productivity, both biological and economic. Estimates of leaf biomass allow the quantification of forest resources such as availability of wood fuel and tree fodder. In addition, estimates of stem and crown biomass are essential for studies of carbon stocks (Schmidt et al. 2009).
Allometric equations are the simplest, fastest and most cost-efficient method for estimating biomass in forest ecosystems (Wang 2006). Allometric equations that relate biomass of standing trees with easily measurable biometric indices can be applied to forest inventorying, estimation of tree growth, and estimation of standing volume (Ketterings et al. 2001; Zianis and Mencuccini 2004), including belowground biomass (Peichl and Arain 2007). These equations are very common and there is a strong scientific basis for planted trees and common timber tree species. In recent years, they have also been applied to many native species and natural forests to estimate carbon pools (Chave et al. 2005).
Many studies have been conducted in recent years to develop allometric equations aiming at estimating biomass of different parts of trees. However, there is little information for trees that grow in low density stands, such as wood pastures. Also allometric equations for pruned trees are scarce, especially for trees whose canopy is periodically harvested to feed livestock or other uses. While this is a marginal practice in some areas of the world, it is still used in many other regions, especially in arid and semiarid regions where herbaceous pastures are not present most part of the year and livestock depend on supplementary fodder provided by tree leaves through pruning, pollarding, shredding and/or coppicing. Pollarding and livestock grazing in forest ecosystems are performed in different countries and has a century-long history. This practice is common in semiarid regions, where livestock farming depends on tree foliage to overcome the shortage of herbaceous forages in winter. Thakur and Thakur (2007) reported multiple multi-purpose tree species that are periodically harvested to feed livestock in many Asian agroforestry systems. Willow species are important species in cold desert agroforestry systems in the northwestern Himalaya (Rawat and Everson 2013). Previous studies showed that fodder trees are important feed sources for livestock in a wide range of farming systems in Africa (Alemu et al. 2013; Berhe and Tanga 2013; Franzel et al. 2014; Geta et al. 2014; Guyassa et al. 2014). Peri et al. (2016) reported silvopastoral systems have become an economical, ecological and productive alternative in South America (Argentina, Chile and Southern Brazil). Black locust (Robinia pseudoacacia) was proposed to broaden grazing period in USA (Burner et al. 2006). Either because the water shortage imposed by the climate and/or to maximize leaf production (and also understory pasture) trees usually grow scattered at low density, which likely affects the utility of allometric relationships.
Northern Zagros forests in the west of Iran with an area of 449,000 ha (F.R.W.O. 2015) are very important in providing livelihood needs of local residents and non-timber forest products (NTFPs) such as forage, galls, acorns and mastic resin. Residents of rural areas in northern Zagros are highly dependent on forests due to a common use of animal husbandry (Jazirehi and Ebrahimi Rastaghi 2003). A special form of silvopastoral management has been developed in northern Zagros with the aim of providing required forages for feeding livestock during the cold season. In the traditional forest management, still common in northern Zagros, an area of forest (up to 40 ha) is customarily owned by a household. The forest stand owned by local residents is divided into three sections. In a 3-year process, local residents pollard oak trees (i.e., Lebanon oak: Quercus libani, Gall oak: Q. infectoria and Persian oak: Q. brantii) and each year one section will be pollarded. Pollarding will be done in the section which has oak trees with 3-year-old branches. Leafy branches are cut and stored in situ to be used as winter forage storage for livestock, especially goat and sheep (Ghazanfari et al. 2004; Valipour et al. 2014) (Fig. 1). Lebanon oak is a very important species in the silvopastoral system, common in the northern Zagros, due to higher palatability of its leaves (Ranjbar et al. 2012).

(Photo “a” and “b” by: S. Shahabedini; photo “c” by: L. Khedri and photo “d” by: Z. Shakeri)
Pollarding by local residents (a); stacks of leafy branches resulting from pollarding (b); storing of leafy branches on special trees in forest (c); winter feeding of goat and sheep using tree fodder (d).
In previous studies regarding tree biomass conducted in Iran, equations including crown height (CH), the product of CH and crown diameter (CD) (CH × CD) (R 2 = 0.94) and DBH (R 2 = 0.95) were reported to estimate dry weight of leaf biomass of Quercus brantii and Pistacia atlantica, respectively (Adl 2007); DBH squared and tree height (R 2 = 0.69) were used to estimate leaf biomass of Celtis caucasica (Pourhashemi et al. 2012); CD index was used for estimating crown biomass of Pistacia atlantica var. mutica (R 2 = 0.92) (Sohrabi and Shirvani 2012); and DBH squared and tree height (R 2 = 0.99) were reported as biometric indices to estimate Fagus orientalis trunk biomass (Vahedi et al. 2013). DBH was used to estimate leaf biomass of 16 species in the Brazilian savanna (Salis et al. 2006). In Ireland, several allometric equations were reported to estimate leaf biomass using biometric indices (i.e., DBH, crown density and CH) of Picea sitchensis trees (Tobin et al. 2006). In a study conducted in Burkina Faso on Jatropha curcas, an allometric relationship was reported between leaf biomass and trunk diameter at a height of 20 cm with the coefficient of determination of 0.66 (Bayen et al. 2016). All proposed tree biomass models for the main hardwood forest species in Spain included tree diameter, whereas for the majority of species total height was only considered in the stem biomass models and in some of the branch models (Ruiz-Peinado et al. 2012).
The objectives of this study are: (1) to investigate the relationship between leaf biomass (as the dependent variable) and biometric indices of Lebanon oak trees such as collar diameter, DBH, total height, trunk height, CH, CD, and crown area (CA) (as independent variables) in pollarded trees; (2) to analyze allometric equations and select the most appropriate regression model to estimate the leaf biomass of Lebanon oak trees; and (3) to estimate the tree fodder production in typical stands of pollarded Lebanon oak trees.
Materials and methods
Study site
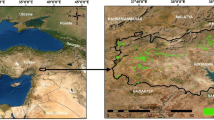
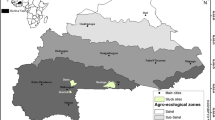
The study area (latitude: 35°45ʹ–36°15ʹN, longitude: 45°50ʹ–46° 15ʹE) with an altitude of 1620 m above sea level is located near Baneh city, Kurdistan province, northern Zagros region, western Iran. The area includes low-lying hills and parent material composed of schist, conglomerate, shale, and metamorphosed limestone generating mainly lithosol (Anonymous 2005). The main tree species of the area are Lebanon oak (Q. libani) and Gall oak (Q. infectoria). Persian oak (Q. brantii), wild pear (Pyrus spp.), hawthorn (Crataegus spp.) are also present at low frequency. Average annual precipitation in the study area is 657 mm, of which 26% occurs in spring, less than 1% in summer, 26% in autumn, and 47% in winter. The average annual temperature is 13.7 °C.
Data collection
To conduct the study, a pollarded stand (6.6 ha in area including 560 Lebanon oak trees, DBH range: 5–33 cm) under traditional silvopastoral management system was selected. This research was conducted in the section where tree branches are 3 years old. In this operation, only young branches (3 or less years old) are harvested. The location and boundary of the selected stand were determined using a GPS device (GARMIN 78 S-accuracy: ±10 m). A full callipering (i.e., a 100 percent inventory) was then conducted and collar diameter (COD) and stem diameter at breast height (DBH) of all trees (collar diameter ≥5 cm) were recorded. In addition, tree height (HTO), and trunk height (HT) were measured using the clinometers (Suunto PM-5) and CD in two perpendicular directions (using measuring tape) were measured before pollarding in summer 2013. At the pollarding time (September 2013) by a customary owner of selected stand (local forester), leafy branches were stacked. Fresh weight of leafy branches was recorded immediately after harvesting using a digital scale (20 g accuracy). For each of the six diameter classes, two trees were randomly selected as a sample to obtain the humidity and coefficient of dry weight for leaves (CDWL), defined as the ratio of the dry weight of leaf to the dry weight of branch and leaf (Eq. 1). For these 12 sample trees, leaves were separated from branches and the fresh weight of each one was recorded separately. The branches and leaves collected from sample trees were transported to the laboratory to determine their humidity and dry weights. Leaves and branches were oven-dried at 80 °C for 48 and 72 h, respectively. A limited number of samples were weighed once every 24 h until the weight became stable. Equation (1) was used to determine CDWL.
where DWL is dry weight of leaves (kg); DWBL is dry weight of branches and leaves (kg); and CDWL is the coefficient of dry weight for leaves (%). These coefficients, together with the humidity content, were then applied to the 560 pollarded trees (stratified by diametric classes) to estimate the leaf biomass from the weighted leafy branches recorded in the field. Dry weight of the leaf biomass was calculated by multiplying dry weight of the stacks of leafy branches by CDWL.
Data analysis
To test for normality of data distributions, we used Kolmogorov–Smirnov test (Lilliefors 1967). The correlation between dry weight of leaf biomass (as dependent variable) and collar diameter, DBH, total height, trunk height, CH, average CD, and CA (as predictors) as well as correlation between biometric indices of 560 Lebanon oak trees were analyzed using Pearson correlation coefficient. Different linear and non-linear models (i.e., logarithmic, inverse, quadratic, cubic, power, compound, exponential) were evaluated through regression analysis. As the non-linear models were not significantly better than the linear ones, the work focused on multiple linear regressions of the form:
where b 0 is the intercept of the regression line with the y-axis; x n are the biometric indices (predictors), b n are the coefficients of predictors, and y is the dry weight of leaf biomass (the dependent variable). Natural logarithmic transformation was applied to data of leaf biomass, which improved significantly goodness of fit of the model.
Three predictors were used, (1) stem diameter, measured as diameter of the collar and DBH, (2) height, measured as tree height, trunk height and CH, and (3) crown size, measured as CD and CA. We compared models with different combinations of predictors (i.e., diameter, height and crown size). Given that variables within each category of predictors were highly correlated (Fig. S1 in the supplementary material) only one variable from each category was included in a given model. Models including three variables were compared with simpler models with one and two variables. Selection of allometric equations requires careful consideration of their fitting (Chave et al. 2005). In this study, a model with the lowest AIC (Akaike Information Criterion), was selected (Akaike 1998). AIC deals with the trade-off between the goodness of fit of the model and the complexity of the model, and it is very useful to select the most parsimonious model. However, AIC does not inform about the quality of the model in an absolute sense. So, we evaluated the validity of models through the adjusted coefficient of determination (R 2adj.), t test of regression coefficients (β parameters) of retained variables, the root mean square error (RMSE) (Eq. 3) and Bias (Eq. 4). Statistical data analysis was performed using SPSS 16.0 and Linear Model Function (lm) in R.
In Equations 3 and 4, n is the number of samples; y i is the measured leaf biomass weight (kg); ŷ i is the leaf biomass weight estimated using the proposed allometric model (kg).
Results
The main characteristics of the studied pollarded stand are presented in Table 1. CDWL for the entire crown was 36.0%.
As expected, most of the biometric variables correlated significantly with each other and with leaf biomass, either for the whole crown (Table 2). Leaf biomass (as the dependent variable) showed the highest correlation with CD (as the predictor). Natural logarithmic transformation (ln) of leaf biomass led to an increase in correlation coefficient (Table 2). The correlation coefficient for the relationship between the logarithm of leaf biomass and CD for entire crown was 0.720 (Table 2).
Among the multiple linear regressions compared, only models that include crown size, either CD or CA, performed well (Table 3; Fig. 2). By including stem diameter the models improved slightly, with the combination of CD with DBH being the most parsimonious model (AIC = 578; R 2 = 0.54; RMSE = 0.4025) (Table 3). Other models that combine crown size (either CD or CA) with stem diameter (either DBH or COD) showed also satisfactory goodness of fit, with DBH performing better than COD, and CD performing better than CA. Height, either of the tree (HTO) or of the crown (HC), gives less significant adjustments. Indeed, adding tree height, either HTO or HC, barely improves the monovariate or bivariate models (only a very slight increase of R 2 and reduction of RMSE), and in all cases, produces less parsimonious models (higher AIC) (Table 3; Fig. 2). HTO and HC are, in fact, not significant in any of the bivariate or trivariate models (Table S1 in the supplementary material). At the end, we proposed an allometric model for leaf biomass of the entire crown (Table 4).

Change of AIC, R 2 and RMSE in bivariate models compared to the most parsimonious monovariate and trivariate models
Figure 3 compares measured leaf biomass versus estimated leaf biomass of total crown (using the proposed, more parsimonious, model). Overall, leaf biomass is slightly overestimated for small trees and sub-estimated in large trees. In addition, the dispersion of data increases with tree size, which can be explained by the reduced sample size for large trees in the studied stand. Indeed, most of the trees are included in the diameter classes of 10–15 and 15–20 cm of DBH; these two intermediate size classes include 70% of the leaf biomass. In total, the studied stand produced 662 kg dry leaves per ha in the cycle of three years, while the modeled estimation reached 616 ± 93 SE kg dry leaves per ha (Table 5).

Measured leaf biomass (points) versus estimated leaf biomass (plan) of Lebanon oak, potted against crown diameter and stem diameter at breast height (DBH)
Discussion
In this study, data were collected from 560 Lebanon oak trees with 5–33 cm DBH to estimate dry weight of leaf biomass (tree fodder) in this commonly pollarded tree species. This allowed us to develop allometric equations that facilitate the quick assessment of tree fodder production in these characteristic wood pastures.
The analysis of the relationship between biometric indices (predictor variables) and leaf biomass (dependent variable) showed that the natural logarithmic transformation of leaf biomass increased the correlation coefficient (Table 2). As tree age increases, woody biomass increases more than leaf biomass. Indeed, leaf biomass tends to stabilize while stem diameter keep growing. Consequently, the relationship among tree size and leaf biomass usually departs from linearity and frequently follow a logarithmic function (Chave et al. 2005).
On the basis of a regression analysis among the studied biometric indices, CD and DBH were the best predictor variables in the suggested equation. Most studies reported DBH as the predictor variable for estimating leaf biomass (Salis et al. 2006; Wang 2006; Calvo-Alvarado et al. 2008; Bayen et al. 2016). In this study, however, DBH did not show a strong correlation with leaf biomass (Table 2). The difference of this research with previous studies in unpollarded stands is that we are working with managed trees whose crown is shaped by periodical pollarding. This makes it more difficult to get robust allometric equations using classical biometric indices. The standardized coefficients (β) for CD (β = 0.664) and DBH (β = 0.161) indicated that the importance of CD was greater than DBH in estimating leaf biomass of Lebanon oak trees (Table 4). As expected in heavily pollarded trees, pruners shape trees to maintain them manageable, and big trees with thick stems have not big (tall) crowns. Due to pollarding in the studied stand, branches and leaves will not have the same chance to grow compared to the diameter; therefore, Lebanon oak trees have young branches and leaves (3–4 years old) despite their old age. The relationships among DBH and crown parameters are also altered in periodically pollarded trees compared to unmanaged trees. In these circumstances, it is not surprising that there was not a very strong correlation between leaf biomass and DBH. In line with the findings of our research, a study on Picea sitchensis in Ireland reported the coefficient of determination resulting from the allometric relationship between DBH and leaf biomass to be 0.72 (Tobin et al. 2006); but with the insertion of CH and crown width variables into the model, the coefficient of determination increased (R 2 = 0.91) while the standard error decreased. In addition, the use of CD index was reported in estimations of leaf biomass of Quercus brantii (Adl 2007), leaf biomass of Pistacia atlantica (Panahi et al. 2014) and crown biomass of Pistacia atlantica var. mutica in Iran (Sohrabi and Taheri Sarteshnizi 2012). Applying equations here presented to data of future forest inventory will inform on the potential of pollarded tree plantations to feed livestock in traditional smallholder farms of Northern Zagros of Iran, and other semiarid regions with oak foliage as the base of winter fodder (Tolunay et al. 2014; Jazib 2015; López-Díaz et al. 2015; Marañón et al. 2016).
In this work, a model to estimate leaf biomass of entire crown was proposed (R 2 = 0.540). The moderate goodness of fit found for the allometric equations of this species is explained by the strong modification of canopy structure through periodical pollarding. In open pollarded stands, tree crowns grow more laterally than in dense forest and vertical distribution of leafy branches are less stratified. Previous studies reported that coefficient of determination for estimating leaf biomass was less than that for the biomass of other tree parts (Bayen et al. 2016). This can be attributed to differences in the development of crown and growth of the leaves of scattered trees which influence shade, microclimate and competition.
In the pollarding process, with the aim of providing leaves (tree fodder), local people cut the branches of Lebanon oak trees, and as a result, CH and crown area are significantly reduced. Although the crown is partially restored one year after pollarding due to resprouting, the branches are harvested again 3 or 4 years later before complete restoration. Consequently, the crowns of Lebanon oak trees in pollarded stands constantly have severe fluctuations without any opportunities for completing growth (Ranjbar et al. 2012). Therefore, crowns of these trees do not grow along with other tree parts. In these circumstances, leaf biomass estimation becomes less accurate than that of other tree components (Kenzo et al. 2009; Sawadogo et al. 2010). Estimation accuracy of forest biomass depends on the diversity and quality of data used to develop allometric equations because the correlation between biomass and biometric indices of trees in different habitat conditions (soil type, species, forest structure, forest density, age and weather conditions) is different (Clark and Clark 2000; Lehtonen 2005). This study illustrates the challenge of estimating leaf biomass and other crown parameters using classical allometric equations. It also shows that leaf biomass estimates may be improved using crown biometric parameters, whose measurement is becoming easier with technology such as LIDAR.
We find that our model overestimated leaf biomass for small trees while fitted much better for bigger trees. Although we have no explanation for this bias, this could be indicating that the smallest trees, likely juvenile trees, are less resilient to the pollarding process. Although moderate defoliation (e.g., by pruning) is common in many fruit and forest species to promote fruit production and knot-free timber, respectively, without important effects on stem growth, a severe pruning can reduce very significantly the tree growth (Neilsen and Pinkard 2003). Resprouting after loss of practically all aboveground biomass consume huge reserves of carbohydrates and nutrients from underground portions of the trunk and/or root system to support expansion of the first leaf of new resprouts (Del Tredici 2001; Zhu et al. 2012). Resprouting usually is more vigorous in juvenile than in mature trees (Bond and Midgley 2001), and young trees should have a greater capacity to rebuild its crown because it has less respiring tissue than an older tree (Prunesim 1991). Among a variety of oak species, sprouting decline with increasing stump diameter and/or age, although sprouting percentages among oaks, even at large diameters, are very high compared with those of other tree species (Del Tredici 2001). However, our results point out that periodical pollarding could suppress the resprouting capacity of some small trees. Hence, for the sake of the sustainability of the systems, we recommend to reduce the intensity of pollarding practice in young trees. Further studies should assess the long-term physiological response and growth of pollarded trees.
Conclusion
Allometric equations are widely used for the quick and indirect estimate of stand aboveground biomass and C pools. They rely on the strong correlations commonly found among different biometric parameters of trees, with stem diameter and tree height being most frequently used. However, allometric equations for pollarded trees are still challenging. Using the allometric equations described in this study, it will be possible to estimate dry weight of leaf biomass of Lebanon oak trees in the pollarding practice to feed livestock in semiarid regions with oak foliage as the base of winter fodder. Having accurate information on dry weight of leaf biomass of oak trees makes it possible to provide management recommendations for pollarded stands, to facilitate harvest planning, to allow accurate evaluation of pollarded stand’s conditions, to enable their monitoring and to ensure the sustainability of this traditional practice. In addition, leaf biomass estimate could be used to evaluate the impact of pollarding on nutrient cycling and carbon sequestration in forest. In this way, one can decide about management plans by determining the amount of organic material and nutrients extracted from the forest.
Finally, working with scattered trees, where lateral branches develop easily (due to no or low competition with neighboring trees), make challenging the use of allometric equations. These two facts differentiate this research with respect to many others conducted in dense wood formations, whose crown growth is driven by biophysical factors (also genetic) and competition among trees.
Author contribution statement
Conceived and designed the experiments: Loghman Ghahramany. Performed the experiments: Loghman Ghahramany, Sima Shahabedini and Sheyda Khosravi. Analyzed the data: Loghman Ghahramany, Sima Shahabedini, Gerardo Moreno and Fernando Pulido. Contributed reagents/materials/analysis tools: Loghman Ghahramany, Sima Shahabedini and Gerardo Moreno. Wrote the paper: Loghman Ghahramany, Sima Shahabedini, Gerardo Moreno, Fernando Pulido and Sheyda Khosravi.
References
Adl HR (2007) Estimation of leaf biomass and leaf area index of two major species in Yasuj forests. Iran J Poplar Res 15:417–426 (Persian)
Akaike H (1998) Information theory and an extension of the maximum likelihood principle. In: Parzen E, Tanabe K, Kitagawa G (eds) Selected papers of Hirotugu Akaike. Springer, New York, pp 199–213
Alemu B, Animut G, Tolera A (2013) Millettia ferruginea: an endemic legume tree as forage for ruminants in southern and northwestern Ethiopia. Livest Res Rural Dev 25:3
Anonymous (2005) Multipurpose forest management plan; emphasis on organizing and management of pollarding in Armardeh, west of Iran. The center of Research and Development on Forest Management in Northern Zagros, University of Kurdistan, Baneh (Persian)
Bayen P, Bognounou F, Lykke AM et al (2016) The use of biomass production and allometric models to estimate carbon sequestration of Jatropha curcas L. plantations in western Burkina Faso. Environ Dev Sustain 18:143–156. doi:10.1007/s10668-015-9631-4
Berhe DH, Tanga AA (2013) Nutritional evaluation of Ficus thonningii Blume leaves as ruminant livestock feed in the Ahferom district of Tigray, Ethiopia. Afr J Range Forage Sci 30:149–154. doi:10.2989/10220119.2013.765505
Bond WJ, Midgley JJ (2001) Ecology of sprouting in woody plants: the persistence niche. Trends Ecol Evol 16:45–51. doi:10.1016/s0169-5347(00)02033-4
Burner DM, Pote DH, Ares A (2006) Foliar and shoot allometry of pollarded black locust, Robinia pseudoacacia L. Agrofor Syst 68:37–42. doi:10.1007/s10457-006-0001-y
Calvo-Alvarado JC, McDowell NG, Waring RH (2008) Allometric relationships predicting foliar biomass and leaf area: sapwood area ratio from tree height in five Costa Rican rain forest species. Tree Physiol 28:1601–1608
Chave J, Andalo C, Brown S et al (2005) Tree allometry and improved estimation of carbon stocks and balance in tropical forests. Oecologia 145:87–99. doi:10.1007/s00442-005-0100-x
Clark DB, Clark DA (2000) Landscape-scale variation in forest structure and biomass in a tropical rain forest. For Ecol Manage 137:185–198. doi:10.1016/S0378-1127(99)00327-8
Del Tredici P (2001) Sprouting in temperate trees: a morphological and ecological review. Bot Rev 67:121–140. doi:10.1007/bf02858075
Forest, Range and Watershed Management Organization of IR of Iran (F.R.W.O.). http://frw.org.ir/00/En/StaticPages/Page.aspx?tid=13237. Accessed Mar 2015 (Persian)
Franzel S, Carsan S, Lukuyu B et al (2014) Fodder trees for improving livestock productivity and smallholder livelihoods in Africa. Curr Opin Environ Sustain 6:98–103. doi:10.1016/j.cosust.2013.11.008
Geta T, Nigatu L, Animut G (2014) Evaluation of potential yield and chemical composition of selected indigenous multi-purpose fodder trees in three districts of Wolayta Zone, Southern Ethiopia. World Appl Sci J 31:399–405
Ghazanfari H, Namiranian M, Sobhani H, Marvi Mohajer MR (2004) Traditional forest management and its application to encourage public participation for sustainable forest management in the northern Zagros mountains of Kurdistan Province, Iran. Scand J For Res 19:65–71. doi:10.1080/14004080410034074
Guyassa E, Raj AJ, Gidey K, Tadesse A (2014) Domestication of indigenous fruit and fodder trees/shrubs in dryland agroforestry and its implication on food security. Int J Ecost 4:83–88
Jazib MJ (2015) Oak: a multipurpose tree species of the Pir Panjal Himalayas facing grave threats. Middle East J Sci Res 23:665–669
Jazirehi MH, Ebrahimi Rastaghi M (2003) Silviculture in Zagros. University of Tehran, Tehran (Persian)
Kenzo T, Furutani R, Hattori D et al (2009) Allometric equations for accurate estimation of above-ground biomass in logged-over tropical rainforests in Sarawak, Malaysia. J For Res 14:365. doi:10.1007/s10310-009-0149-1
Ketterings QM, Coe R, van Noordwijk M et al (2001) Reducing uncertainty in the use of allometric biomass equations for predicting above-ground tree biomass in mixed secondary forests. For Ecol Manag 146:199–209. doi:10.1016/S0378-1127(00)00460-6
Lehtonen A (2005) Estimating foliage biomass in Scots pine (Pinus sylvestris) and Norway spruce (Picea abies) plots. Tree Physiol 25:803–811. doi:10.1093/treephys/25.7.803
Lilliefors HW (1967) On the Kolmogorov–Smirnov test for normality with mean and variance unknown. J Amer Stat Assoc 62:399–402. doi:10.1080/01621459.1967.10482916
López-Díaz ML, Rolo V, Benítez R, Moreno G (2015) Shrub encroachment of Iberian dehesas: implications on total forage productivity. Agrofor Syst 89:587–598. doi:10.1007/s10457-015-9793-y
Marañón T, Ibáñez B, Anaya-Romero M et al (2016) Oak trees and woodlands providing ecosystem services in Southern Spain. In: Trees beyond the wood conference proceedings. pp 291–299
Neilsen WA, Pinkard EA (2003) Effects of green pruning on growth of Pinus radiata. Can J For Res 33:2067–2073. doi:10.1139/x03-131
Panahi P, Pourhashemi M, Hasaninejad M (2014) Allometric equations of leaf biomass and carbon stocks of oaks in National Botanical Garden of Iran. J Plant Res 27:12–22 (Persian)
Peichl M, Arain MA (2007) Allometry and partitioning of above- and below-ground tree biomass in an age-sequence of white pine forests. For Ecol Manag 253:68–80. doi:10.1016/j.foreco.2007.07.003
Peri PL, Bahamonde HA, Lencinas MV et al (2016) A review of silvopastoral systems in native forests of Nothofagus antarctica in southern Patagonia, Argentina. Agrofor Syst 90:933–960. doi:10.1007/s10457-016-9890-6
Pourhashemi M, Eskandari S, Dehghani M et al (2012) Biomass and leaf area index of Caucasian Hackberry (Celtis caucasica Willd.) in Taileh urban forest, Sanandaj, Iran. Iran J For Poplar Res 19:609–620 (Persian)
Prunesim F (1991) A biological justification for pruning in coastal Douglas-fir stands. Weed Sci 32:413–417
Ranjbar A, Ghahramany L, Pourhashemi M (2012) Impact assessment of pollarding on biometrical indices of Lebanon oak (Quercus libani Oliv.) in Belake Forests, Baneh. Iran J For Poplar Res 20:578–594 (Persian)
Rawat YS, Everson CS (2013) Availability and use of willow species in representative cold desert areas of northwestern himalaya, India. J Mt Sci 10:472–481. doi:10.1007/s11629-013-2687-4
Ruiz-Peinado R, Montero G, del Rio M (2012) Biomass models to estimate carbon stocks for hardwood tree species. For Systems 21:42–52. doi:10.5424/fs/2112211-02193
Salis SM, Assis MA, Mattos PP, Pião ACS (2006) Estimating the aboveground biomass and wood volume of savanna woodlands in Brazil’s Pantanal wetlands based on allometric correlations. For Ecol Manag 228:61–68. doi:10.1016/j.foreco.2006.02.025
Sawadogo L, Savadogo P, Tiveau D et al (2010) Allometric prediction of above-ground biomass of eleven woody tree species in the Sudanian savanna-woodland of West Africa. J For Res 21:475–481. doi:10.1007/s11676-010-0101-4
Schmidt A, Poulain M, Klein D et al (2009) Allometric above-belowground biomass equations for Nothofagus pumilio (Poepp. & Endl.) natural regeneration in the Chilean Patagonia. Ann For Sci 66:513. doi:10.1051/forest/2009030
Sohrabi H, Shirvani A (2012) Allometric equations for estimating standing biomass of Atlantic Pistache (Pistacia atlantica var. mutica) in Khojir National Park. Iran J For 4:55–64 (Persian)
Sohrabi H, Taheri Sarteshnizi MJ (2012) Fitting probability distribution functions for modeling diameter distribution of oak species in pollarded northern Zagros forests (Case study: Armardeh-Baneh). Iran J For 4:333–343 (Persian)
Thakur PS, Thakur A (2007) Canopy management in agroforestry-options and strategies. In: Puri S, Panwar P (eds) Agroforestry systems and practices. New India Publishing Agency, New Delhi, pp 533–545
Tobin B, Black K, Osborne B et al (2006) Assessment of allometric algorithms for estimating leaf biomass, leaf area index and litter fall in different-aged Sitka spruce forests. Forestry 79:453–465. doi: 10.1093/forestry/cpl030
Tolunay A, Adıyaman E, Akyol A et al (2014) An investigation on forage yield capacity of Kermes Oak (Quercus coccifera L.) and grazing planning of Mediterranean maquis scrublands for traditional goat farming. Sci World J 2014:9
Vahedi A, Mataji M, Babayi-Kafaki S et al (2013) Modeling the bole mass of beech (Fagus Orientalis Lipsky) through allometric equations within Hyrcanian forests. Iran J For 5:309–322 (Persian)
Valipour A, Plieninger T, Shakeri Z et al (2014) Traditional silvopastoral management and its effects on forest stand structure in northern Zagros, Iran. For Ecol Manag 327:221–230
Wang C (2006) Biomass allometric equations for 10 co-occurring tree species in Chinese temperate forests. For Ecol Manag 222:9–16. doi:10.1016/j.foreco.2005.10.074
Zhu W-Z, Xiang J-S, Wang S-G, Li M-H (2012) Resprouting ability and mobile carbohydrate reserves in an oak shrubland decline with increasing elevation on the eastern edge of the Qinghai–Tibet Plateau. For Ecol Manag 278:118–126. doi:10.1016/j.foreco.2012.04.032
Zianis D, Mencuccini M (2004) On simplifying allometric analyses of forest biomass. For Ecol Manag 187:311–332. doi:10.1016/j.foreco.2003.07.007
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Funding
This study was funded by the University of Kurdistan, Sanandaj, Iran.
Additional information
Communicated by E. Priesack.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Shahabedini, S., Ghahramany, L., Pulido, F. et al. Estimating leaf biomass of pollarded lebanon oak in open silvopastoral systems using allometric equations. Trees 32, 99–108 (2018). https://doi.org/10.1007/s00468-017-1614-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00468-017-1614-7