
Abstract
Holm oak (Quercus ilex ssp. ballota [Desf.] Samp) is one of the most important species in forest communities of the western Mediterranean Basin, but is very vulnerable to environmental stress during the first years of its life. In particular, summer drought and winter frosts limit the distribution, survival, and growth of Holm oak. These two stress factors can lead to plasmolysis and/or oxidative stress. We hypothesized that autumn fertilization with nitrogen (N) can improve plant tolerance to these abiotic stress factors and improve plant quality and therefore reforestation success. A 12-week autumn application of 30 or 60 mg N (plus 70 mg N applied to both treatments during the previous 28 weeks in the nursery, i.e. 100 and 130 mg N in total, respectively) improved overall growth, root growth capacity, frost tolerance, and nutritional status of plants relative to plants given 0.0 and 1.5 mg autumnal N, and had no negative effect on seedling response to water stress. A very small increment in N doses during the autumn (1.5 vs. 0.0 mg N) improved some morphological parameters, such as stem diameter (D) and shoot dry weight, and physiological parameters, such as total antioxidant activity. The highest autumnal N dose (60 mg) increased leaf nutrient content without decreasing the concentration, but had a tendency to reduce frost tolerance relative to seedlings given a lower autumnal N dose (30 mg).
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Holm oak (Quercus ilex ssp. ballota [Desf.] Samp) is one of the most important species in forest communities of the western Mediterranean Basin (Palacios et al. 2009). This evergreen sclerophyllous species has been increasingly used for reforestation (MAPA 2006). However, Holm oak usually has poor early out-planting performance compared to other Mediterranean species, particularly in sites with unfavorable climatic conditions (Pausas et al. 2004). This poor performance has been attributed to water stress (Villar-Salvador et al. 2004a, b), low site fertility (Pardos et al. 2005; Sanz-Pérez et al. 2007; Valdecantos et al. 2006), and poor seedling quality (del Campo et al. 2010).
Drought and frosts are the main factors that limit the distribution, abundance, survival, and growth of this species in Mediterranean areas (Larcher 2000), and Holm oak is very vulnerable to stress conditions during early life (Ruiz de la Torre 2006). Afforestation projects conducted with this species are usually subjected to long periods of low temperatures and frosts and extreme summer drought, especially in continental areas (Aranda et al. 2005; Corcuera et al. 2005; García-Plazaola et al. 1999; Gimeno et al. 2009), limiting the survival and growth after field planting. The exposure to suboptimal non-lethal growth temperatures causes a depression of photosynthesis and photosystem II efficiency (Strand and Öquist 1985), exacerbating the formation of reactive oxygen species (ROS) in chloroplasts and oxidative stress (Mittler 2002). Oxidative stress damages cell membranes by inducing lipid peroxidation, protein oxidation, enzyme inhibition, and DNA damage (Briat 2002; Clijsters et al. 1999). Detoxification of ROS is carried out by enzymatic mechanisms and by a system of physiological antioxidant molecules, which can be lipophilic (tocopherols, carotenoids) or hydrophilic (ascorbate, glutathione) (García-Plazaola et al. 1999; Márquez-García et al. 2009; Márquez-García and Córdoba 2010). Cold-tolerant plants respond to low temperatures by increasing the levels of these antioxidant compounds and the activity of some antioxidative enzymes (García-Plazaola et al. 1999). Therefore, the maintenance of adequate levels of antioxidants in plants could improve the physiological status and, consequently, field performance of Holm oak. As far as we know, there are not studies on oxidative stress applied to nursery plants subjected to sublethal low temperatures.
Nursery practices, environmental conditions, and genetic factors affect the functional characteristics of seedlings and field performance after transplantation (Birchler et al. 1998). Manipulation of nutrient availability in the nursery can modify morphophysiological characteristics of seedlings (Puttonen 1997), although the relationship with field performance is uncertain for species of Quercus (Broncano et al. 1998; Oliet et al. 2009; Trubat et al. 2010; Villar-Salvador et al. 2004a, 2005). During the hardening phase in the nursery, numerous environmental factors (including chill temperature, short photoperiod, irrigation, and fertilization) can induce plant dormancy and improve hardiness (Colombo et al. 2003; Fernández et al. 2008). N fertilization can improve photosynthetic rate, root growth, and water-use efficiency (Fernández et al. 2006; Lambers et al. 1990). A previous study with Holm oak has reported that N has a greater effect than P or K on morphological and nutritional status and improvement of seedling frost tolerance (Andivia et al. 2011). However, the latter assay only studied two levels of N fertilization and the nursery fertilization programs for this species require additional refinement (Jacobs et al. 2009).
In order to contribute to the knowledge of the Holm oak nursery fertilization program, we hypothesized that autumnal N fertilization can affect growth and minerals content but also the cellular response to a sublethal dosage of an abiotic stress-like frosts. Therefore, the main objectives of this study were: (i) to evaluate the effect of four different doses of autumn fertilization with N on morpho-physiological characteristics of seedlings and (ii) to explore the oxidative stress reaction to a non-lethal frost tolerance test of nursery plants subjected to different autumnal N doses.
Materials and methods
Plant material and growth conditions
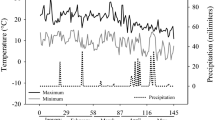
During autumn 2008, acorns of Q. ilex ssp. ballota (Desf.) Samp. were collected from the Spanish provenance region of La Alcarria y Serranía de Cuenca (39°25′–41°20′N, 1°40′–3°15′W). At the beginning of 2009, 600 acorns were pre-germinated in a growth chamber at 20°C on wet perlite to reduce germination time and intra-population variability. On the third week of February 2009, 360 healthy germinated acorns were randomly selected and sown in 300 cm3 Plasnor® cells (45 cells per tray, 8 trays) that contained sphagnum peat Kekkilä® B0 (pH corrected to 6.0 using Dolokal®). Seedlings were grown in a nursery under white shade-cloth that reduced radiation by ~50%. Daily nursery temperatures are shown in Fig. 1. All trays were well watered, moved weekly, and rotated to eliminate microsite effects during the entire study period.

Daily minimum (T Min) and maximum (T Max) temperature (°C) at the nursery during the study period (February 2009–2010)
During 28 weeks (third week of March to the end of September), a constant fertilization regimen was applied using a water-soluble fertilizer (Peters professional® 20-20-20) at a rate of 125 ppm N, 54 ppm P, and 104 ppm K. Each seedling received a weekly dose of 2.5 mg N (3.94% ammoniacal N, 6.05% nitrate N, 10.01% urea N, all w/w), 1.088 mg P, and 2.075 mg K (monoammonium phosphate, monopotassium phosphate, and potassium nitrate). These weekly doses were called N 1, P 1, and K 1 and corresponded to 70.0 mg N, 30.5 mg P, and 58.1 mg K over the entire 28-week period. The fertilizer was added to each seedling once per week in distilled water; plants were watered as needed with tap water between fertilizer applications. At the end of the first 28 weeks, seedlings had a mean ± SD height of 9.37 ± 0.74 cm, stem diameter of 2.51 ± 0.11 mm, shoot dry weight (ShDW) of 1.21 ± 0.09 g, root dry weight of 2.53 ± 0.34 g, shoot-to-root dry weight ratio of 0.59 ± 0.03, and 1.01% N, 0.09% P, 0.61% K in the leaves.
After the 28 weeks period 270 healthy seedlings were randomly chosen (33–34 plant per tray) and distributed in 6 trays (45 cells per tray). For the next 12 weeks (first week of October to third week of December, the hardening phase), four different dose of N were used (N 0, N 1/20, N 1, N 2). Two different dose of N per tray were tested (each applied to half of the plants) so that in the six trays all fertilization treatments were combined (N 0–N 1/20, N 0–N 1, N 0–N 2, N 1/20–N 1, N 1/20–N 2, N 1–N 2) with 67–68 seedlings given each treatment and distributed in three trays. Autumn fertilization was applied using a nutrient solution composed at the laboratory, using calcium nitrate and ammonium sulfate as nitrogen sources. The doses of N were weekly applied in a single weekly dose. Over the entire 12-week period, each plant received 0.0 mg N (N 0), 1.5 mg N (N 1/20), 30.0 mg N (N 1), or 60.0 mg N (N 2). The doses of P and K were constant and similar as in the previous 28 weeks, and each seedling received during the 12 weeks of the hardening phase 13.1 mg P and 24.9 mg K. Plants developed new sprouts from late summer to the end of October, and there were no significant differences in the number of sprouted plants among treatments. In early November, no plants had visual signs of shoot elongation. The weekly doses of N 1, P 1, and K 1 were the same as in the previous 28 weeks, and close to those typically recommended in forest nurseries for Holm oak during the growing phase (Navarro-Cerillo et al. 2009). A group with low N dose (N 1/20) was used to avoid total nutrient deprivation (Boivin et al. 2002). Total N deprivation was tested by dose N 0, and N 2 was used to test a high dose of N, but was still within the optimal range for this species (Jacobs et al. 2009).
Morphological and nutritional status
At the end of December 2009, six plants per treatment (2 seedlings per tray) were randomly chosen for assessment of morphological and nutritional status. Shoots were cut at the cotyledon insertion point, leaves and stems were separated, and roots were cleaned. Shoot height was measured and stem diameter was measured 0.5 cm above the cotyledon insertion point. All samples were washed with distilled water, oven-dried at 65°C until no further weight reduction occurred, and the dry weights of leaves (LDW), stems (StDW), and roots (RDW) were measured. The shoot dry weight (ShDW = LDW + StDW) and shoot-to-root dry weight ratio (SRDW = ShDW/RDW) were calculated.
Mineral nutrients (N, P, K) were only determined in leaves because Andivia et al. (2011) reported that differences in nutrient concentration and content of plants given different N fertilization treatments were much greater in leaves than in other tissues. To obtain leaf samples sufficiently large for nutrient analysis, three groups per dose of N were formed. Each group was formed with two plants belonging to a tray. For N analysis, an elemental analyzer (Termo Finnigan 1112 Series EA, Milan, Italy) was used. For P and K analyses, dry samples were treated at 550°C for approximately 7 h, subjected to acid digestion in 5 M HCl, and analyzed by ICP-OES Jobin Yvon Ultima 2 (Tokyo, Japan). Non-structural carbohydrates (NSC) were also determined. Hydroalcoholic extraction and anthrone colorimetry (using a spectrophotometer; UV-1601, Shimadzu®, Tokyo, Japan) was used to determine soluble sugars (SS) (Spiro 1966) and acid hydrolysis followed by anthrone colorimetry was used to determine starch (St) (Rose et al. 1991). Nutrient and NSC levels (in mg) were calculated as the multiplication of concentration (%) and leaf dry mass (g).
Frost tolerance
Cold hardiness was assessed by a detached leaf test, as described for Eucalyptus globulus L. (Fernández et al. 2007) and previously tested on Holm oak (Andivia et al. 2011). A temperature programmable freezer (West® 4400; ISE Inc., Cleveland, OH) with two internal fans to remove air was used. The test began at 12°C, and the temperature was reduced by 3°C h−1 to the minimum. The temperature was maintained at the minimum for 3 h, and was then increased by 5°C h−1 to 12°C. A complete cycle lasted 14–16 h.
The detached leaf tests were performed in the third week of December 2009. Our previous experience (Andivia et al. 2011; Fernández et al. 2008) and the preliminary checking of the plants guaranteed that the effect of autumn fertilization was evident in that time. Two fully expanded leaves per temperature and plant (one from the medium-higher part of the shoot and other from the medium-lower part) and four plants per treatment (1–2 per tray randomly selected) were sampled and tested at minimum temperatures of −8, −10, −12, and −14°C. There were 239 h of total chilling time (≤8°C) accumulated in the nursery at the time of the freezing tests. Selected leaves were inserted into test tubes (2.2 cm diameter, 15 cm length), and placed in the freezer. After completion of temperature regimen, leaves were removed from the freezer, distilled water (≤2 mL) was added until the cross-section of the short petiole was immersed, and samples were maintained in a growth chamber (25°C day/17°C night; photoperiod of 12 h; ≥70% relative humidity; 350 μmol m−2 s−1 photosynthetic photon flux). Damage was assessed at 24, 48, 72, and 96 h after the end of the freezing test by measurement of the leaf percentage that had visual damage (VD), defined as dimming and/or browning of the leaf due to cell lysis. To estimate visual damage, each leaf lamina was observed by dividing it into two longitudinal parts considering each part as the 50% of the leaf surface (we did not account the central vein), and we estimated visually the surface of each part that was dimming and/or browning. The VD final data was the average of the damage estimated percentages of each part.
At the end of January 2010, six plants per treatment (2 per tray) were randomly selected (treatments N 0, N 1/20, N 1) to assess frost-induced oxidative stress. At this time, plants had reached maximum cold hardiness in this nursery (Andivia et al. 2011), so it was possible to apply a very low (−12°C) but non-lethal minimum temperature to induce a plant reaction. Plants given treatment N 2 were not tested because our previous results (December, 2009) indicated no significant differences in cold tolerance for plants given doses N 1 and N 2. There were 493 h of total chilling time (≤8°C) accumulated in the nursery at the time of the freezing test. Twelve of the 18 plants selected (4 per treatment) were subjected to a freezing cycle, as previously described for detached leaf tests, with a minimum temperature of −12°C. Selected seedlings were placed in Plasnor® trays of 300 cm3 and put into a cardboard box filled up to the cotyledon insertion point with polystyrene to prevent the freezing of roots. Other six plants (2 per treatment) were used as controls and maintained in a growth chamber under the same conditions as in the detached leaf tests. After the freezing cycle, plants were also kept in the growth chamber with control plants for 24 h. Leaves were cut, the central nerve was removed, and they were immediately frozen in liquid nitrogen and stored at −80°C. Frost-induced oxidative stress was determined by measurement of lipid peroxidation, level of ascorbic acid (AsA), and total antioxidant activity (AAT).
The extent of lipid peroxidation was estimated by measurement of the level of malondialdehyde (MDA), as described by Hodges et al. (1999). Frozen leaves were ground in a pre-cooled mortar and pestle with liquid N2 and homogenized in 80% (v/v) ethanol. The extract was centrifuged at 3,000g for 10 min and the supernatant was used for the assay. This method corrects for interference caused by anthocyanins. A 1 mL aliquot of a diluted sample was added to a test tube with 1 mL of either: (i) TBA solution, containing 20% (w/v) trichloroacetic acid and 0.01% butylated hydroxytoluene, or (ii) +TBA solution, containing the above plus 0.65% of TBA. Samples were mixed and heated at 95°C for 25 min and then cooled and centrifuged at 3,000g for 10 min. Absorbance was read at 440, 532, and 600 nm using a spectrophotometer (Beckman Coulter DU 800) and the concentration of MDA equivalents (nmol mL−1) was calculated by the following equations:
Reduced AsA was quantified by the bipyridyl method (Knörzer et al. 1996). For extraction, leaves stored at −80°C were weighed and macerated with liquid N2 in porcelain mortars using metaphosphoric acid at 5% (w/v). The homogenate was centrifuged at 10,000g for 30 min and the absorbance was measured at 525 nm. Total ascorbate was determined after incubation with dithiothreitol (DTT), and oxidized ascorbate (DHA) was calculated as the difference between total and reduced levels of ascorbate.
The AAT of the extracts was determined using 2,2-diphenyl-1-picryl-hydrazyl radical (DPPH·) method as described by Brand-Williams et al. (1995), with some modifications (Leong and Shui 2002; Wong et al. 2006). An aliquot fraction (40 μL) of a methanolic plant extract was added to 3 mL of 0.1 mM DPPH in 30% methanol, vortexed, and then left to stand at room temperature for 30 min. The antioxidant capacity based on DPPH· scavenging activity was expressed as μmol trolox equivalents (TE) per mg plant dry weight (DW).
Root growth capacity and drought resistance
Root growth capacity (RGC) was assessed in December 2009 using six plants per treatment (2 plants per tray) randomly selected. Seedlings were carefully removed from tray cells, the white root tips were cut, and seedlings were planted with undamaged plugs in 2.5 L pots with wet perlite. Pots were randomly distributed on a heating table (20°C) inside a greenhouse for 4 weeks. Seedlings were watered every 2–3 days and given no fertilization. After 4 weeks, seedlings were cleaned and the total new roots dry mass (TRW) was measured.
To evaluate the resistance to water stress, eight plants per treatment (2–3 plants per tray) were randomly taken and randomly distributed between two polystyrene boxes (16 plants per box). The boxes (50 L volume) were filled with a mixture of perlite and vermiculite (1:1) and the plants were transplanted on 30 March 2010. Then, the boxes were placed inside a greenhouse and not watered during the test. At time of transplantation, the height and number of leaves of each plant were determined. At the beginning of this test plants had started the new spring growth and morphological parameters were still significantly different between N treatments. From the second week (April 9), predawn water potential (pressure chamber Model 1000, PMS Instruments, Corvallis, OR, USA) of these plants was measured every 2–4 days until 29 April 2010. One leaf per plant from the central part of the stem was used for these measurements.
Data analysis
The effect of autumn fertilization on seedling morphology, nutritional status, and RGC was assessed using a general linear model (GLM):
where y ij is the value of the dependent variable in plant j, or group of plant j in the case of nutritional status, fertilized at dose i of N (i = 1, 2, 3, 4); μ is the overall mean; N is the fixed effect of N fertilization, and e is the error term, with the hypothesis e ij ~ N(0, σ 2e ). When the dependent variable was a percentage, the transformation \( \arcsin \sqrt {{{\text{var} (\% )} \mathord{\left/ {\vphantom {{\text{var} (\% )} {100}}} \right. \kern-\nulldelimiterspace} {100}}} \) was used. Significant differences between effects were assessed by Tukey’s HSD test, with α = 0.05.
The influence of fertilization on VD was estimated by a mixed model:
where y ijklm is \( \arcsin \sqrt {{{VD(\% )} \mathord{\left/ {\vphantom {{VD(\% )} {100}}} \right. \kern-\nulldelimiterspace} {100}}} \) of leaf i of plant j, tested at temperature k, measured after l hours (24, 48, 72, and 96 h), at dose m of N (m = 1, 2, 3, 4); α is a random plant effect with hypothesis α ~ N(0, σ 2p ); t is the temperature fixed effect; h is the measurement time fixed effect; N is the fixed effect of N fertilization; and e is the error term under the hypothesis of Normal distribution. The other terms describe interactions of the main effects. The analysis considered that (1) observations belonging to the same plant can be correlated and (2) that observations taken in the same leaf at different measurement times (24, 48, 72, and 96 h) can also be correlated and have heterogeneous variance. To test (1) we tested the significance of the plant random effect with a likelihood ratio test, as the reduction of the statistic −2 × log likelihood (−2LL), after introducing the plant random effect follows a χ2 distribution with 1 degree of freedom. For testing (2) we tested different variance–covariance structures for the submatrix corresponding to the observations taken at the same leaf at the four different times. The structures tested were (Wolfinger 1996) for a description of the variance–covariance structures): diagonal with homogeneous variance, unstructured with 1, 2, 3, and 4 bands, toeplitz with 1, 2, 3, and 4 bands, heterogeneous toeplitz with 1, 2, 3, and 4 bands, autoregressive heterogeneous and first-order ante-dependence. Comparison of different structures have been done with Akaike’s information criteria (AIC, the smaller the better). Variance components for each structure were estimated by restricted maximum likelihood (REML).
The effect of autumn N fertilization on oxidative stress (MDA, AAT, and AsA) was assessed using a GLM:
where y ijk is the value of the dependent variable in plant k fertilized at dose i of N (i = 1, 2, 3) and subjected to the frost cycle j (j = 1, 2); μ is the overall mean; N is the fixed effect of the N fertilization; T is the effect of being subjected or not to a frost cycle, and e is the error term for the hypothesis e ijk ~ N(0, σ 2e ). Significant differences between effects were assessed by Tukey’s HSD test, with α = 0.05.
The influence of N fertilization on water stress resistance was estimated by a mixed model:
where y ijk is the predawn water potential of plant i, evaluated at date j, fertilized with a dose k of N (n = 1, 2, 3, 4); α is the random plant effect with hypothesis α~ N(0, σ 2p ); H is the effect of the covariable height; m is the effect of the covariable number of leaves; N is the fixed effect of the N fertilization; F is the fixed effect of the measurement date; and e is the error term with the hypothesis of Normal distribution. The double term (FN) describes the interaction between the dose of N and the measurement date. We have considered first the seedling as random effect, hence considering that observations taken in the same plant are correlated with covariance equal to σ 2p and that covariance of observations taken in different seedlings is null. We have compared that model structure with more complex ones that consider different structures of the variance–covariance matrix of the observations taken in the same seedling. We tested the same structures as in the visual damage (VD) analysis.
Results
Morphological and nutritional status of seedlings
Autumn N fertilization had a significant effect (p < 0.001) on all measured morphological parameters except in shoot-to-root dry weight ratio (SRDW) (p = 0.838) (Table 1). Seedlings that were fertilized with high doses of N (N 1 and N 2) had greater height (H), diameter (D), ShDW, and root dry weight (RDW) than seedlings fertilized with low doses (N 0 and N 1/20). There were also significant differences between N 0 and N 1/20 for D (p = 0.049) and between N 1 and N 2 for ShDW (p = 0.027), with higher doses associated with greater D and ShDW.
Table 2 shows that autumn N fertilization significantly increased leaf N and P concentrations (p < 0.010) and leaf N, P, and K contents (p < 0.001), with higher doses having a greater effect. There were no significant differences between N 0 and N 1/20 doses (p > 0.900). Leaf nutrient contents were higher in seedlings fertilized with N 2 than in those fertilized with N 1 (p < 0.010), although significant differences between N 1 and N 2 doses were not found for leaf N concentration (p = 0.217) and for leaf P concentration (p = 1.000).
There were no significant effects of autumn N fertilization on leaf soluble sugars (SS) (p = 0.831) and starch (St) (p = 0.796) concentrations (4.45 and 27.87%, respectively). However, seedlings fertilized with N 2 had higher leaf SS contents (72.03 vs. 36.92 mg) (p < 0.001) and St (512.60 vs. 212.74 mg) (p < 0.050) than seedlings fertilized with N 0 and N 1/20. Leaf SS and St contents of seedlings fertilized with N 1 were between these values, with no significant differences (p > 0.200).
Frost tolerance
The selected model indicated a significant plant random effect (p = 0.007). The selected variance–covariance matrix of observations from the same leaf measured at different times was the first-order ante-dependence, hence indicating correlation between measurements from the same leaf and heterogeneous variance at different measurement times. Our results indicated significant differences in visual damage (VD) between all tested temperatures (p < 0.001) except −10 and −12°C. VD significantly (p < 0.001) increased with time since the freezing test (24, 48, 72, and 96 h). Autumn N fertilization also significantly affected VD (p < 0.001). Seedlings given lower doses of N (N 0 and N 1/20) had greater VD than those given higher doses (N 1 and N 2) at −8 and −10°C (Fig. 2): at −12°C significant differences were only found between N 1 dose and N 0 and N 1/20 dose whereas at −14°C significant differences were found between N 1 and N 2 doses and N 0 dose. Seedlings given N 1 dose showed lower VD than those given N 2 dose for all temperature tested, although differences were not significant (p > 0.245).

Least squares means estimates of visual damage (VD) for the combination of N doses and temperatures. Different letters indicate significant differences between N doses (p < 0.05) for each tested temperature
In general, different doses of N had little or no significant effect on oxidative stress parameters or on controls and seedlings subjected to low temperature. The lipid peroxidation values ranged from 13.00 to 10.41 nmol MDA g−1 (fresh weight), without significant differences. The AsA levels ranged from 112 to 134 μmol reduced AsA g−1 (fresh weight) and 119 to 140 μmol total AsA g−1 (fresh weight). The percentage of reduced AsA was about 95% in all treatments, with no significant differences (data not shown). However, there were significant differences in total AAT in control and treated plants (Fig. 3). In particular, seedlings subjected to the freezing cycle had higher AAT levels than controls. In addition, seedlings fertilized with N 0 had significantly lower AAT than those fertilized with N 1/20 and N 1.

Total antioxidant activity (AAT, μmoles trolox equivalents [TE] per g leaf dry weight) of seedlings fertilized with different N doses (N 0, N 1/20 and N 1) and subjected to or not subjected to a frost cycle (frost vs. control). Different letters indicate significant differences (p < 0.05) between N doses (lowercase letters) and between frost cycle (uppercase letters). 18 seedlings were sampled, 6 per dose of N and 4 of them were subjected to the frost cycle, the other 2 were used as controls
RGC and drought resistance
Autumn N fertilization had a significant effect on RGC (p = 0.004). In particular, seedlings fertilized with N 1 and N 2 developed more new roots than seedlings fertilized with N 0 and N 1/20 (12.19 mg TRW [N 0], 24.31 mg TRW [N 1/20], 86.75 mg TRW [N 1], 78.30 mg TRW [N 2]).
Our analysis of water stress indicated no significant differences in water potential in plants given different doses of N (p = 0.082), with all N treatments leading to a similar decline of water potential, despite having the same soil moisture. The selected variance covariance matrix for observations from the same plant was the first-order ante-dependence. Covariates, height and number of leaves, were not significant (p = 0.158 and p = 0.821, respectively). At the beginning of this test seedlings given N 1 and N 2 were significantly taller (p < 0.050) than seedlings given N 0 and N 1/20 doses. Mean water potential at the beginning of the test was −0.36 MPa and at the end of the test was −3.70 MPa with no significant differences for the interaction between the measurement time and the dose of N (p = 0.457).
Discussion
Nitrogen fertilization usually increases morphological parameters and nutritional status of Holm oak nursery seedlings (Broncano et al. 1998; Oliet et al. 2009; Sanz-Pérez et al. 2007; Villar-Salvador et al. 2004a). In agreement to this, our results and those obtained by Andivia et al. (2011), indicated that autumn N fertilization also increases the H, D, ShDW and RDW of Holm oak seedlings. This increment took place during a three months period, and could be enough for increasing plant quality (Andivia et al. 2011). The differences in morphology observed in the present study indicated that some morphological parameters, such as diameter and shoot mass, are very sensitive to small changes in N fertilization during autumnal hardening (0.0 vs. 1.5 mg), at least for a nursery located in a warm temperate zone. However, Oliet et al. (2011) did not find differences in biomass increment due to fall fertilization, although fall fertilization enhanced root P concentration, root growth potential and field performance; and some previous studies did not report an increase in RDW by N fertilization (Oliet et al. 2009; Villar-Salvador et al. 2004a). The differences between these studies and the present study may be due to the mild autumn temperatures of our nursery (in Huelva) and/or the addition of fertilizer from early October until mid-December rather than early autumn or a shorter period during autumn. In any case, the increment in growth and nutritional status could enhance plant quality and field response after transplanting by improving root growth potential, frost tolerance, field growth and a phenologically earlier development (Cuesta et al. 2010; Oliet et al. 2011).
Our results indicate that N fertilization during hardening phase increased the nutrient levels of seedlings. The N, P, K content of the leaves of seedlings fertilized with highest dose (N 2) were higher than those fertilized with N 1, probably due to the greater biomass in N 2 seedlings, as indicated by the greater leaf mass. Jacobs et al. (2009) also reported higher N, P and K content in fertilized plants and showed that doses of 200 mg N per plant led to no toxicity. However, parameters such as root volume, which may predict a good post-transplant response, decreased at N doses higher than 100 mg. In our study, N 2 (130 mg N) dose increased shoot mass and the accumulation of nutrients, did not diminish the response to water and cold stress, but tended to reduce RGC and tolerance to frost relative to N 1, although this was not statistically significant. Thus, it could be recommended doses of about 100 mg N fertilization during Holm oak cultivation in nurseries. However, we must take into account that applying 70 mg of N during growing stage (before hardening occurs) plus 30 mg in autumn could lead to smaller plants than applying 100 mg during growing phase. In addition, the dose adjustment may be needed depending on local weather conditions and the ultimate destination of seedlings (Cuesta et al. 2010; Trubat et al. 2011), an issue which should be addressed in future studies. The autumnal N doses studied had no significant effect on non-structural carbohydrates concentration. Our use of autumn N fertilization prevented the growth-mediated dilution of leaf carbohydrates concentration, which would have been significant if we used only spring fertilization (Fernández et al. 2008; Palacio et al. 2008; Sanz-Pérez et al. 2007). Furthermore, our N 2 seedlings had higher leaf carbohydrates contents than the N 0 and N1/20 seedlings, because, despite having N 2 seedlings higher values of biomass, there was no significant decrease in leaf carbohydrates concentrations.
Previous studies have reported different effects of N fertilization on cold hardiness (literature cited in Fernández 2008; Vilagrosa et al. 2006). Our leaf detached test data indicate that autumn N fertilization improved frost tolerance of Holm oak seedlings. Several studies reported a positive association between soluble sugars concentration and frost tolerance in Quercus species (Fernández et al. 2008; Mollá et al. 2006; Morin et al. 2007). However, our data showed similar soluble sugars concentration and content in plants given different doses of N. Although the accumulation of soluble sugars may improve cold hardiness and osmotic cryogenic properties, elevated levels of other molecules and solutes, such as lipids, amino acids, proteins (those with apoplastic antifreeze activity, cryoprotective proteins, dehydrins, etc.), organic acids, glycosides and organic salts can also improve cold tolerance (Larcher 2000; Zwiazek et al. 2001). We also found that N fertilization increased N and P leaf concentration and leaf content in seedlings. P is an important constituent of the phospholipids of cell membranes (Marschner 1995), and frost-induced damage first occurs in cell membranes (Heino and Palva 2003; Kaminska-Rozek and Pukacki 2005). The higher N and P leaf concentration might explain the positive effect of N fertilization on cold hardiness, in fact seedlings given N 1 and N 2 doses showed similar cold tolerance, probably due to the similar values of N and P leaf concentration as well as carbohydrates concentrations.
We found no difference in the lipid peroxidation and AsA content of plants subjected to frost treatment and controls, presumably because the test we used (−12°C) was not cold enough to cause serious damage to plants. Lipid peroxidation reactions lead to loss of membrane stability, and can be stopped if the plant has sufficient levels of antioxidants (Blokhina et al. 2003; Briat 2002; Shaw et al. 2004). AsA is well known to play an important role in the prevention of oxidative stress (Foyer 1993; Smirnoff 1998, 2000). The concentration of oxidized AsA increases as oxidative stress increases; however, in healthy plants, the reduced form constitutes approximately 90% of the total AsA (Foyer 1993; Smirnoff 2000). In our study, the reduced form of AsA was approximately 95% of the total which, together with our lipid peroxidation data, indicates that our plants were not significantly stressed by our freezing test. However, seedlings subjected to a freezing cycle had higher total AAT values than the controls, indicating that our treatment led to some oxidative stress that was presumably tolerated due to other detoxification mechanisms, possibly hydrophilic (glutathione, AsA) or lipophilic (tocopherols and carotenoids) antioxidant molecules (Walker and McKersie 1993; Wise and Naylor 1987). For example, in a study of Q. ilex ssp. ballota, García-Plazaola et al. (1999) reported an increase of α-tocopherol, β-carotene, and AsA during the cold months.
Our data indicate that seedlings deprived of N during the autumn hardening phase (N 0) had lower antioxidant capacity than seedlings fertilized with N (N 1/20 and N 1). This suggests that the addition of a very small amount of N during the cold hardening phase (e.g. N 1/20) might be sufficient to prevent frost-induced oxidative stress. Besides, it would have been very interesting to check the response of N 2 treatment, as there was a pattern of higher VD for N 2 than for N 1, VD was conducted on detached leaves rather than whole plant and N 1/20, that experienced the same VD as N 0, had a higher AAT than N 0. Nevertheless, our results are preliminary and further studies are needed to assess the effect of lower temperatures and the role of other antioxidants involved in alleviating oxidative stress. It should be also necessary to conduct frost tests at higher photon flux because the combination of low temperatures and high photon flux would be expected to increase the production of reactive oxidative species in chloroplasts (Foyer et al. 1994).
Autumn N fertilization increased RGC, in agreement with other studies (Mollá et al. 2006; Oliet et al. 2011). In particular, seedlings fertilized with high dose N (N 1 and N 2) had more new roots than seedlings fertilized with low dose N (N 0 and N 1/20). RGC has been used as a predictor of transplantation performance of forest species, and is correlated with cold hardiness in many species (Simpson and Ritchie 1996).
We found no significant differences in the water potential of plants given different N doses. Harvey and van den Driessche (1999) concluded that highly fertilized plants are more vulnerable to water stress because they have a greater increase in aerial parts and this leads to increased transpiration. Although our fertilized plants were larger, and may have experienced a greater loss of water via transpiration (Hernández et al. 2009), we found no significant differences in the shoot-to-root dry weight ratio, leading to a similar balance between absorbed and transpired water (Lloret et al. 1999).
In conclusion, autumn N fertilization of Holm oak seedlings increased their growth, nutrient content and concentration, cold hardiness, and RGC. However, seedlings fertilized with different N doses had similar responses to water stress. Our results indicate that some morphological parameters, such as diameter and shoot mass, are sensitive to small increases of N and that seedlings deprived of N during the autumn hardening phase had lower antioxidant capacity, suggesting that the addition of a small amount of N during the cold hardening phase might be sufficient to prevent frost-induced oxidative stress in mild winter locations. Our data also confirm that seedlings fertilized with doses of 100 mg N (30 mg in autumn) and seedlings fertilized with 130 mg N (60 mg N in the fall) had similar values of plant quality parameters, although the effect of these doses of N in post-transplanting response should be corroborated, especially in areas with unfavorable climatic conditions.
References
Andivia E, Fernández M, Vázquez-Piqué J (2011) Autumn fertilization of Quercus ilex ssp. ballota (Desf.) Samp. nursery seedlings: effects on morpho-physiology and field performance. Ann For Sci 68:543–553
Aranda I, Castro L, Alía L, Pardos JA, Gil L (2005) Low temperature during winter elicits differential responses among populations of the Mediterranean evergreen cork oak (Quercus suber). Tree Physiol 25:1085–1090
Birchler T, Rose RW, Royo A, Pardos M (1998) La planta ideal: revisión del concepto, parámetros definitorios e implementación práctica. Inv Agrar-Sist Rec F 7:109–121
Blokhina O, Virolainen E, Fagerstedt KV (2003) Antioxidants, oxidative damage and oxygen deprivation stress: a review. Ann Bot Lond 91:179–194
Boivin JR, Miller BD, Timmer VR (2002) Late-season fertilization of Picea mariana seedlings under greenhouse culture: biomass and nutrient dynamics. Ann For Sci 59:255–264
Brand-Williams W, Cuvelier ME, Berset C (1995) Use of a free radical method to evaluate antioxidant activity. Food Sci Technol LEB 28:25–30
Briat JF (2002) Metal ion-activated oxidative stress and its control. In: Inzé D, Montagu M (eds) Oxidative stress in plants. Taylor and Francis, London, pp 171–189
Broncano MJ, Riba M, Retana J (1998) Seed germination and seedling performance of two Mediterranean tree species. Holm oak (Quercus ilex L.) and Aleppo pine (Pinus halepensis Mill.): a multifactor experimental approach. Plant Ecol 138:17–26
del Campo AD, Navarro-Cerillo RM, Ceacero CJ (2010) Seedling quality and field performance of commercial stocklots of containerized Holm oak (Quercus ilex) in Mediterranean Spain: an approach for establishing a quality standard. New For 39:19–37
Clijsters H, Cuypers A, Vangronsveld J (1999) Physiological responses to heavy metals in higher plants: defence against oxidative stress. Z Naturforsch C 54:730–734
Colombo SJ, Glerum C, Webb DP (2003) Daylength, temperature and fertilization effects on desiccation resistance, cold hardiness and root growth potential of Picea mariana seedlings. Ann For Sci 60:307–317
Corcuera L, Morales F, Abadia A, Gil-Pelegrin E (2005) Seasonal changes in photosynthesis and photoprotection in a Quercus ilex subsp. ballota woodland located in its upper altitudinal extreme in the Iberian Peninsula. Tree Physiol 25:599–608
Cuesta B, Villar-Salvador P, Puértolas J, Jacobs DF, Rey Benayas JM (2010) Why do large, nitrogen rich seedlings better resist stressful transplanting conditions? A physiological analysis in two functionally contrasting Mediterranean forest species. For Ecol Manage 260:71–78
Fernández M (2008) Endurecimiento en vivero de especies leñosas mediterráneas destinadas a plantación forestal. Cuad Soc Esp Cienc For 28:13–24
Fernández M, Alejano R, Domínguez L, Tapias R (2008) Temperature controls cold hardening more effectively than photoperiod in four Mediterranean broadleaf evergreen species. Tree For Sci Biotech 2:43–49
Fernández M, Marcos C, Tapias R, Ruiz F, López G (2007) Nursery fertilisation affects the frost-tolerance and plant quality of Eucalyptus globulus Labill. cuttings. Ann For Sci 68:865–873
Fernández M, Novillo C, Pardos JA (2006) Effects of water and nutrient availability in Pinus pinaster Ait. open pollinated families at an early age: growth, gas exchange and water relations. New For 31:321–342
Foyer CH (1993) Ascorbic acid. In: Alscher EG, Hess JL (eds) Antioxidants in higher plants. CRC Press, Florida, pp 32–54
Foyer CH, Lelandais M, Kunert KJ (1994) Photooxidative stress in plants. Physiol Plantarum 92:696–717
García-Plazaola JI, Artetxe U, Becerril JM (1999) Diurnal changes in antioxidant and carotenoid composition in the Mediterranean sclerophyll tree Quercus ilex (L.) during winter. Plant Sci 143:125–133
Gimeno TE, Pías B, Lemos-Filhos JP, Valladares F (2009) Plasticity and stress tolerance override local adaptation in the response of Mediterranean Holm oak seedlings to drought and cold. Tree Physiol 29:87–98
Harvey HP, Van den Driessche (1999) Nitrogen and potassium effects on xylem cavitation and water-use efficiency in poplars. Tree Physiol 19:943–950
Heino P, Palva ET (2003) Signal transduction in plant cold acclimation. In: Hirt H, Shinozaki K (eds) Plant responses to abiotic stress. Top Curr Gen Vol 4. Springer, Berlin, pp 151–185
Hernández EI, Vilagrosa A, Luis VC, Llorca M, Chirino E, Vallejo VR (2009) Root hydraulic conductance, gas exchange and leaf water potential in seedling of Pistacea lentiscus L. and Quercus suber L. grown under different fertilization and light regimes. Environ Exp Bot 67:269–276
Hodges DM, DeLong JM, Forney CF, Prange RK (1999) Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 207:604–611
Jacobs DF, Salifu KF, Oliet JA (2009) Sobrecarga exponencial de nutrientes para la optimización de la fertilización en vivero de plantas de Quercus ilex L. In: Sociedad Española de Ciencias Forestales, Junta de Castilla y León (eds) Proceedings V Congreso Forestal Español, vol 1. Ávila, Spain, pp 11
Kaminska-Rozek E, Pukacki PM (2005) Effect of freezing desiccation on cold hardiness, ROS, membrane lipid levels antioxidant status in spruce seedling. Acta Soc Bot Pol 3:219–228
Knörzer OC, Durner J, Boger P (1996) Alterations in the antioxidative system of suspension cultured soybean cells (Glycine max) induced by oxidative stress. Physiol Plantarum 97:388–396
Lambers H, Cambridge ML, Konings H, Pons TL (1990) Causes and consequences of variation in growth rate and productivity of higher plant. SPB Academic Publishing, Amsterdam, p 363
Larcher W (2000) Temperature stress and survival ability of Mediterranean sclerophyllous plants. Plant Biosyst 134:279–295
Leong LP, Shui G (2002) An investigation of antioxidant capacity of fruits in Singapore markets. Food Chem 76:69–75
Lloret F, Casanovas C, Peñuelas J (1999) Seedlings survival of Mediterranean shrubland species in relation to root: shoot ratio, seed size and water and nitrogen use. Funct Ecol 13:210–216
MAPA (2006) Forestación de tierras agrícolas. Ministerio de Agricultura y Pesca, Madrid, p 373
Márquez-García B, Córdoba F (2010) Antioxidative system in wild populations of Erica andevalensis. Environ Exp Bot 68:58–65
Márquez-García B, Fernández MA, Córdoba F (2009) Phenolics composition in Erica sp. differentially exponed to metal pollution in the Iberian Southwestern Pyritic Belt. Bioresour Technol 100:446–451
Marschner H (1995) Mineral nutrition of higher plants. Academic Press, London, p 889
Mittler R (2002) Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci 7:405–410
Mollá S, Villar-Salvador P, García-Fayos P, Peñuelas-Rubira JL (2006) Physiological and transplanting performance of Quercus ilex L. (holm oak) seedlings grown in nurseries with different winter conditions. For Ecol Manage 237:218–226
Morin X, Ameglio T, Ahas R, Kurz-Besson C, Lanta V, Lebourgeois F, Miglietta F, Chuine I (2007) Variation in cold hardiness and carbohydrates concentration from dormancy induction to bud burst among provenances of three European oak species. Tree Physiol 27:817–825
Navarro-Cerillo RM, Pemán J, del Campo A, Moreno J, Lara MA, Díaz JL, Pousa F, Piñón FM (2009) Manual de especies para la forestación de tierras agrarias en Andalucía. Junta de Andalucía, Consejería de Agricultura y Pesca, Sevilla, p 395
Oliet JA, Tejada M, Salifu KF, Collazos A, Jacobs DF (2009) Performance and nutrient dynamics of Holm oak (Quercus ilex L.) seedlings in relation to nursery nutrient loading and post-transplant fertility. Eur J Forest Res 128:253–263
Oliet JA, Salazar JM, Villar R, Robredo E, Valladares F (2011) Fall fertilization of Holm oak affects N and P dynamics, root growth potential, and post-planting phenology and growth. Ann For Sci 68:647–656
Palacio S, Milla R, Albuixech J, Pérez-Rontomé C, Camarero JJ, Maestro M, Montserrat-Martí G (2008) Seasonal variability of dry matter content and its relationship with shoot growth and non-structural carbohydrates. New Phytol 180:133–142
Palacios G, Navarro-Cerrillo RM, del Campo A, Toral M (2009) Site preparation, stock quality and planning date effect on early establishment of Holm oak (Quercus ilex L.) seedlings. Ecol Eng 35:38–46
Pardos M, Royo A, Pardos JA (2005) Growth, nutrient, water relations, and gas exchange in Holm oak plantation in response to irrigation and fertilization. New For 30:75–94
Pausas JG, Blade C, Valdecantos A, Seva JP, Fuentes D, Alloza JA, Milagrosa A, Bautista S, Cortina J, Vallejo R (2004) Pines and oaks in the restoration of Mediterranean landscapes of Spain: new perspectives for an old practice—a review. Plant Ecol 171:209–220
Puttonen P (1997) Looking for the “silver-bullet”—can one test do it? New For 13:9–27
Rose R, Rose CL, Omi SK, Forry KR, Durral DM, Bigg WL (1991) Starch determination by perchloric acid vs enzymes: evaluating the accuracy and precision of six colorimetric methods. J Agric Food Chem 39:2–11
Ruiz de la Torre J (2006) Flora mayor. Ministerio de Medio Ambiente. Organismo Autónomo Parques Nacionales, Madrid
Sanz-Pérez V, Castro-Díez P, Valladares F (2007) Growth versus storage: response of Mediterranean oak seedlings to changes in nutrient and water availabilities. Ann For Sci 64:201–210
Shaw BP, Sahu SK, Mishra RK (2004) Heavy metal induced oxidative damage in terrestrial plants. In: Prassad MNV (ed) Heavy metal stress in plants: from biomolecules to ecosystem. Springer, Berlín, pp 84–126
Simpson DG, Ritchie GA (1996) Does RGP predict field performance? A debate. New For 13:249–273
Smirnoff N (1998) Plant resistance to environmental stress. Curr Opin Biotech 9:214–219
Smirnoff N (2000) Ascorbic acid: metabolism and functions of a multi-facceted molecule. Curr Opin Plant Biol 3:229–235
Spiro RG (1966) Analysis of sugars found in glycoproteins. In: Neufeld EF, Ginsburg V (eds) Methods in ezymology. Vol. VIII, Complex carbohydrates. Academic Press, New York, pp 3–26
Strand M, Öquist G (1985) Inhibition of photosynthesis by freezing temperatures and high light levels in cold-acclimated seedlings of Scots pine (Pinus sylvestris). I. Effects on the light-limited and light-saturated rates of CO2 assimilation. Physiol Plant 64:425–530
Trubat R, Cortina J, Vilagrosa A (2010) Nursery fertilization affects seedling traits but not field performance in Quercus suber L. J Arid Environ 74:491–497
Trubat R, Cortina J, Vilagrosa A (2011) Nutrient deprivation improves field performance of woody seedlings in a degraded semi-arid shrubland. Ecol Eng 37:1164–1173
Valdecantos A, Cortina J, Vallejo VR (2006) Nutrient status and field performance of tree seedlings planted in Mediterranean degraded areas. Ann For Sci 63:249–256
Vilagrosa A, Villar-Salvador P, Puértolas J (2006) El endurecimiento en vivero de especies forestales mediterráneas. In: Cortina J, Peñuelas JL, Puértolas J, Savé R, Vilagrosa A (eds) Calidad de planta forestal para la restauración en ambientes mediterráneos: estado actual de conocimientos. MMA, Organismo Autónomo de Parques Nacionales, Madrid, pp 119–140
Villar-Salvador P, Planelles R, Enriquez E, Peñuelas-Rubira JL (2004a) a Nursery cultivation regimes, plant functional attributes, and field performance relationships in the Mediterranean oak Quercus ilex L. For Ecol Manag 196:257–266
Villar-Salvador P, Planelles R, Oliet JA, Peñuelas-Rubira JL, Jacobs DF, González M (2004b) b Drought tolerance and transplanting performance of Holm oak (Quercus ilex L.) seedlings after drought hardening in the nursery. Tree Physiol 24:1147–1155
Villar-Salvador P, Puértolas J, Peñuelas JL, Planelles R (2005) Effect of nitrogen fertilization in the nursery on the drought and frost resistance of Mediterranean forest species. Invest Agrar Sist Recur For 14:408–418
Walker MA, McKersie BD (1993) Role of the ascorbate–glutathione antioxidant system in chilling resistance of tomato. J Plant Physiol 141:234–239
Wise RR, Naylor AW (1987) Chilling-enhanced photooxidation: evidence for the role of singlet oxygen and superoxide in the breakdown of pigment and endogenous antioxidants. Plant Physiol 83:278–282
Wolfinger RD (1996) Heterogeneous variance-covariance structures for repeated measures. J Agric Biol Environ Stat 1:205–230
Wong SP, Leong LP, Koh JHW (2006) Antioxidant activities of aqueous extracts of selected plants. Food Chem 99:775–783
Zwiazek J, Renault S, Croser C, Hansen J, Beck E (2001) Biochemical and biophysical changes in relation to cold hardiness. In: Bigras FJ, Colombo SJ (eds) Conifer cold hardiness. Kluwer Academic Publishers, Dordrecht, pp 165–186
Acknowledgments
This work has been financed by MEC of Spain (ref. AGL2006-12609-C02-01/FOR). In addition, the work was partially supported by University of Huelva and Andalucía Research Programs to promote research groups activity. First author is benefiting from a doctoral grant from the Ministry of Education of Spain. We gratefully acknowledge the support of the laboratory of research and Agrifood control (LICAH) of the University of Huelva for carrying out chemical analysis.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by R. Hampp.
Rights and permissions
About this article
Cite this article
Andivia, E., Márquez-García, B., Vázquez-Piqué, J. et al. Autumn fertilization with nitrogen improves nutritional status, cold hardiness and the oxidative stress response of Holm oak (Quercus ilex ssp. ballota [Desf.] Samp) nursery seedlings. Trees 26, 311–320 (2012). https://doi.org/10.1007/s00468-011-0593-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00468-011-0593-3