
Abstract
Qilian juniper (Sabina przewalskii kom.) is one of the dominant tree species on Qinghai-Tibetan Plateau and has been used in dendroclimatological studies. Here we designed a test to examine whether or not the climate–growth responses in tree rings of Qilian Juniper vary with the change in tree's age. A total number of 135 increment cores were sampled from Qilian Juniper trees at five sites, in which 112 cores were selected and grouped into five 100-year age classes for analysis of age-dependent climate–growth relationships. Chronology statistics, response functions and ANOVA F-test were used to test the consistency of five age-class mean chronologies (AGCs). The results showed that mean sensitivity (MS) and standard deviation (SD) did not change significantly with age. Response function analysis indicated that (a) climate accounts for a high amount of variance in tree-ring widths; (b) tree-ring growth has significant positive correlation with mean monthly air temperature of previous October and November, and with total monthly precipitation of current January and June, while has significant negative correlation with mean monthly air temperature of current May; and (c) AGC-2, AGC-3 and AGC-4 have stronger response to climate change than AGC-1 and AGC-5. The ANOVA F-test showed that generally there are significant differences between the first age class and other four age classes, but among the four classes in which trees are older than 200 years, the differences are usually insignificant. Overall the long-lived Qilian Juniper is still an ideal tree species for dendroclimatic reconstruction.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
It has long been well documented that tree rings are the most widely used proxy in annually resolving reconstructions of climate (Fritts 1976; Bradley and Jones 1992). Dendrochronological techniques are also commonly used to evaluate current climate–growth relationships of trees (D’Arrigo et al. 1992). The physiological mechanisms whereby climatic parameters are translated into radial growth variations are complex, because radial growth in any given year integrates the effects of climate conditions of the plant in prior years, the age and structure of the trees (Fritts 1976). Despite these complexities, most traditional dendroclimatic and dendroecological analyses have assumed that climate–growth relationships are age-independent, once the biological growth trends and the influence of disturbance pulses have been removed (Szeicz and MacDonald 1994). But ecophysiological studies suggested that growth-related environmental signals are likely to be age-dependent, and lower photosynthetic rates in older trees have been measured in various conifer species (Bond 2000).
Uncertainties about the effect of age on radial growth have caused much concern in dendrochronological studies in recent years. Szeicz and MacDonald (1994) suggested that age-dependent relationship should be an essential part of dendroclimatic analysis. Several studies have tested whether climate–growth response is stable across different age classes and the conclusions varied with different species. There were significant differences in growth-climate relationships between old and young trees in some species, such as Picea glauca (Szeicz and MacDonald 1994), Abies lasiocarpa (Ettl and Peterson 1995) and Larix decidua (Carrer and Urbinati 2004). In contrast, there were no significant differences in growth-climate relationships between young and old trees of some species such as Pinus aristata Engelm (Fritts 1976) and Larix lyalii Parl. (Colenutt and Luckman 1995).
As far as Qilian juniper (Sabina przewalskii Kom.) is concerned, there has been still no available information about its age effects on climate–growth response. Qilian juniper is one of the dominant tree species on the Qinghai-Tibetan Plateau, the highest plateau in the world, where the climate change is associated with Asian Monsoon and other global climate systems (Zhang et al. 2003; Zhang and Qiu 2007; Huang and Zhang 2007). Many tree-ring studies have shown that Qilian juniper is sensitive to climate change and has great potential in dendroclimatic study (Zhang and Wu 1997; Liu et al. 2005; Shao et al. 2005). Since the age-dependent climate–growth relationship might potentially affect the validity of results of past climate reconstructions, the test of this relationship is sorely needed. The objective of our study was to test whether or not the Qilian juniper is age invariate after removal of the biological growth function.
Materials and methods
Study area and climate data
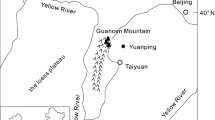
The study sites are located at the upper treeline of the eastern Qilian Mountains where Qilian juniper is the dominant species, with Pinus tabulaeformis, Picea asperata occasionally present. The canopy coverage is 10–40%. The samples were taken from five sites (Fig. 1) and the elevations range from 2,968 to 3,347 m above sea level. The sampled trees were generally growing on slopes facing south with a gradient of 30°–45°. No fire-scarred tree was found, and little is known about any other disturbance history of these sites. One core per tree was extracted from trees at their breast height from the direction parallel to the slope using an increment borer. At three sites (Donggou, Qingyanggou and Mangza), more than 30 undamaged standing trees were sampled for each site. For sites Youhulu-1 and Youhulu-2, increment cores sampled were collected from 35 trees. A total number of 135 increment cores were collected in this study.

Map of sampling sites and meterological stations
There are three meteorological stations very close to the sampling areas and the elevations range from 2271 to 3180 m. All the meteorological stations and sampling sites are located within 100 km. The region is in arid and semi-arid zones of the Northwest China. Mean annual precipitation of the three stations for the last 45 years (1958–2002) is 378 mm, with 91.3% of the precipitation distributed in April–September. Mean annual air temperature is 0.38°C. Mean monthly air temperature and total monthly precipitation both peak in July (Fig. 2).

Climatograph for mean monthly air temperature (°C) and mean total monthly precipitation (mm) of three meteorological stations
Dendrochronological methods
In the laboratory, the increment cores were mounted in slotted wooden holders and polished with sandpaper of progressively finer grit to highlight ring-width patterns. Ring widths were measured with a Velmex Measuring System, which reaches the precision to 0.001 mm. The tree-ring samples were cross-dated by a variety of means according to the standard method introduced by Stokes and Smiley (1968). Firstly the tree rings were visually cross-dated by matching the patterns of narrow and wide rings among samples. Program COFECHA (Holmes 1983) was used to quality check the preliminary results of the cross-dating. The problematic samples were then examined under a microscope to find out where and what the problem was and then corrected them accordingly. In addition, we plotted the sequences for those samples that need more careful cross-dating to find the best matching position of narrow-and-wide ring patterns as well as the pointer years. There are 13 poor quality samples that could not be cross-dated due to too many fragmentation and/or rotten pieces were excluded from further analysis.
For the core samples that did not have pith but were very close to it, the tree’s age was estimated by adding 2–7 rings after comparison of their ring patterns with the cores that had pith (Clack and Hallgren 2004). There were 10 cores sampled far away from the pith were excluded for further study due to the complexity in estimating age. In total, 112 increment cores were used for further analysis. Those cores were grouped into five 100-year age classes based on tree age at breast height in 2005. The 100-year interval was considered as a reasonable compromise between sufficient sample size and sufficient disaggregation of the data (Carrer and Urbinati 2004).
Age-class mean chronologies (AGCs) and individual chronologies were standardized with the program ARSTAN (Holmes 1992) and two standardization approaches were adopted. The first method is negative exponential detrending method which can remove the growth trends from raw ring-width series while retaining most low frequency variations that are potentially climate-driven. Furthermore, this method is always used in climate–growth studies at the treeline or arid and semi-arid area (Jacoby and Cook 1981; Cook 1985; Graumlich 1991). To guarantee the quality of chronologies, we also tried to remove trends with the second method, cubic smoothing spline, in which every tree-ring width series was fitted with a curve, and then each measured value was divided by the correspondent estimated value of the fitted curve. For fitting, a spline function was used with a 50% frequency response of 20 years.
Several fundamental descriptive statistics commonly used in dendrochronology were adopted to assess the high-frequency variations due to climatic fluctuations (Fritts 1976; Rolland 1993). These statistics included mean sensitivity (MS), standard deviation (SD) and signal-to-noise ratios (SNR). Mean ring-width (MRW) was also calculated to test the age related growth trends.
Statistical analysis
Correlations of mean monthly air temperature (Tem) and correlation of total monthly precipitation (Pre) data from three meteorological stations were analyzed. Mean monthly air temperature data were used as they had more significant results than maximum or minimum monthly air temperatures.
Relationships between climate and radial growth were identified by correlation and response function (RF) analysis (Fritts 1976; Briffa and Cook 1990). RF analysis is a liner multiple regression technique that uses the principal components of monthly climatic variables to estimate tree-ring growth, which overcomes the inter-correlation problems among climatic variables. Because the results of RF analysis vary drastically depending on the number of variables used and whether or not previous growth variables are included (Blasing et al. 1984), we also calculated correlation coefficients between ring-width indices (derived from negative exponential detrending method) and climatic variables along with the response function coefficients (Szeicz and MacDonald 1994; Chhin et al. 2004).
The software PRECON (Fritts 1996) was used in the RF analysis and 5,000 bootstrap replications were undertaken. The climate data were 12 mean monthly air temperature (Tem) and 12 total monthly precipitation (Pre) from three meteorological stations. The monthly climatic variables were from October of the year prior to growth (t − 1) and continue to September of the year of growth (t), and one previous growth variable was added for the AGCs’ RF analysis (Szeicz and MacDonald 1995; Wu 1990). The reference period is 1958–2002. Response functions were computed for each AGC and individual tree ring-width index independently.
To test climate influence within the different AGCs more efficiently, ANOVA F-test was applied. In the ANOVA F-test, the response parameter r/s (the partial regression coefficients divided by their standard deviations) values were used. The used r/s values were selected from the results derived from one of the climate–growth relationship analysis methods, in which the results had the clearest overall trend. The used climatic variables were selected by r/s values which should reach the significant difference level at least in one age class. ANOVA analysis was also used to test whether the trees belonging to five sampling sites have different climate–growth response pattern. This test was applied only to the trees belonging to AGC-2 (in which trees scatter at five sample sites) and to the climatic variables which have significant climate–growth response relationships.
The relationships between ages and response function parameters of individual trees were also analyzed. Two main response function variables ( r/s and R 2 (coefficient of determination)) which concisely expressed the amount of climate variability retained in tree-ring width were adopted. Linear regressions were simultaneously performed.
Results
Chronology statistics
Chronology statistics were summarized in Table 1. The mean ring-width (MRW) values show that Qilian juniper has a biological–geometrical trend decreasing with age. Mean sensitivity (MS) does not change greatly for different age-class mean chronologies (AGCs) and different detrending methods. Standard deviation (SD) displays highly homogeneous values in the five age classes, and values derived from negative exponential detrending method are higher than that from cubic spline detrending method. In general, signal-to-noise ratios (SNR) values derived from cubic spline detrending method are much higher than that derived from negative exponential detrending method.
Growth–climate relationships
Mean monthly air temperature and total monthly precipitation for three meteorological stations are all highly correlated. The correlation coefficients for mean monthly air temperatures are all over 99%, while they are more than 86% for total monthly precipitations.
The results of response function analysis (Table 2) indicate that climate accounts for high amount of variance in tree-ring width. Compared with AGCs derived from cubic spline detrending method, AGCs derived from negative exponential detrending method have higher values for variance explanation. Mean V-climates (percentage of the variance in tree-ring width explained by climatic factor) attain 0.538, 0.494 respectively for the AGCs derived from two detrending methods, while the values of mean V-growth (percentage of the variance in tree-ring width explained by previous year growth) are only 0.044, 0.025 respectively, which indicate a very small proportion of the variance in radial growth. The parameters V-climate, V-growth and R 2 (coefficient of determination) have been compared for different age-class mean chronologies and different detrending methods. For negative exponential detrending method, V-climate values increase substantially from AGC-1 to AGC-4 (from 0.327 to 0.677) and decrease in the oldest age class (AGC-5); the value of V-growth is much higher in AGC-1; and the R 2 values change greatly for all AGCs. For cubic spline detrending method, V-climate values do not change greatly over the age classes (from 0.457 to 0.522); the V-growth values are low for all AGCs; and the highest R 2 values were found in AGC-2 and AGC-3.
The results of climate–growth relationship analysis show that throughout most of the life-span, Qilian juniper is always reacting to the same climatic variables. Tree ring growth has significantly positive correlations with mean monthly air temperature of previous October and November, and total monthly precipitation of current January and June, while it has significantly negative correlation with mean monthly air temperature of current May (Fig. 3). Both the simple correlation analysis and response function (RF) analysis for AGCs derived from negative exponential detrending method revealed that AGC-2, AGC-3 and AGC-4 are usually the highest climate sensitive AGCs. In the results of RF analysis for AGCs derived from cubic spline detrending method, the trend is almost the same, but not as clear as the results of negative exponential detrending method.

(a) Age-class response functions for cubic spline detrending method, (b) Age-class response functions for negative exponential detrending method, and(c) Simple correlation. (Notes: r/s, the partial regression coefficients divided by their standard deviations obtained after the 5,000 bootstrap replications; r, correlation coefficient. Climatic variables are coded from 1 (previous October) to 12 (current September))
ANOVA analysis
The results of ANOVA F-test (Table 3) show that generally there are significant differences between the first age class and the other four age classes, but among the four classes in which trees are older than 200 years, the differences are usually insignificant. Three variables (mean monthly air temperature of previous November, total monthly precipitations of current May and August) analyzed display significant differences (P, 0.005) between the AGCs, with the highest F value for the precipitation of May. The result of ANOVA F-test for different sample sites showed that the climate–growth response parameter r/s for the same climatic variable among all sites did not have significant difference except the r/s values of tree-ring growth response to mean monthly air temperature of previous November at Mangza and Qingyanggou (between the two sites; P, 0.05).
The relationships between ages and response function parameters of individual trees
The results of scatter plots (Fig. 4) of tree age versus r/s values showed that the overall trends for total monthly precipitation of current May, mean monthly air temperatures of previous October and November, and mean monthly air temperatures of current June and July are significantly positive, but the overall trends for other climatic variables are negative or very weak, even nonexistent. The results of scatter plots of tree age versus R2 values displayed that the overall trend is significantly positive.

Scatter plot for tree age-response function relationships. (a, b) for response function parameter ( r/s) of selected climatic variables (a) for temperature; (b) for precipitation. (c) for coefficient of determination. (Tem temperature, Pre precipitation. r/s, the partial regression coefficients divided by their standard deviations obtained after the 5,000 bootstrap replications; R 2, coefficient of determination)
Discussion and conclusions
Chronology statistics revealed that the mean ring-width values decreased with age, which might be the reason that the amount of new wood added each year is constant while the circumference increases (Nash et al. 1975), and another reason might be trees have biological growth trend. So selecting a suitable method to detrend the biological growth trend is very important. In our study, results of two detrending methods were compared. MS and SD are similar throughout the different age classes for two detrending method, which are similar to Carrer and Urbinati's findings (2004). However, variance explanations of response function analysis for the five AGCs in two detrending methods are distinct. The AGCs detrended by cubic spline function have lower noise, and the percentage of ring-width variance explained by climate is lower too. The negative exponential detrending method can preserve higher V-climate value for the AGCs, and it may be more suitable for dendroclimatic studies in this area. Considering the heterogeneity of five sample sites, we also take the results of cubic spline detrending method into account.
Site differences can explain why the AGC-2 and AGC-4 have relatively lower signal-to-noise ratios. Trees belonging to AGC-2 scatter at five sample sites, and trees belonging to other AGCs have relatively concentrated distribution. Other studies also pointed out that site differences might modify the manner of age-dependent responses (Fritts 1976; Szeicz and MacDonald 1994). In our study, the chronology characters might be affected by site differences, but the results of ANOVA analysis showed that few of site differences reach the significant level. Even though the diverse chronological structure of the samples might give rise to a weaker age related climate–growth response pattern, it does not change the main conclusions. We think the growth responses of Qilian juniper to climate are related to tree age, though there are heterogeneous components in the general trend.
The results of chronology statistics and response function analyses showed that there are different phases in the long-term dynamics of the climate–growth relationships in Qilian juniper. The radial growths of older trees are more sensitive to climate change than that of young trees. Although the oldest AGC has declined climate sensitivity, ANOVA F-test could not found significant differences among the four classes in which trees are older than 200 years. The relationships between ages and response function parameters of individual trees further confirmed that the climate–growth response of Qilian juniper is age-dependent because r/s and R2 values usually increase with age. Other studies also showed that near the treeline, the sequences include higher climate sensitivity in old stressed trees and that a more prominent growth-climate signal retains in tree rings (Fritts 1976; Travis et al. 1990).
The reason for age-dependent response of radial growth to climate is likely due to physiological changes accompanying with aging. Ryan and Yoder (1997) suggested that water conduction becomes a principal physiological challenge as trees aged. The total resistance of the hydraulic pathway increases as the tree approaches its maximum height under the influences of many factors, such as gravity, a longer and more complex hydraulic path length through stems and branches. Increases in hydraulic resistance can lower the supply of water for transpiration inducing earlier stomatal closure (Ryan et al. 1997; Ryan and Yoder 1997), which in turn limits stomatal conductance and gaseous exchange (Yoder et al. 1994; Hubbard et al. 1999; Kolb and Stone 2000; England and Attiwill 2006). Hydraulic constraints can only partially explain how age may control the climate sensitivity of trees. But the real reason is still not clear since the functional processes linked to tree growth are very complicated. For instance, age-related hydraulic constraints can even lead to genetically based changes in physiology which serve as adaptive response to predictable age-related changes in internal water relations (Day et al. 2002).
In summary, the age-dependent responses are detected between groups of trees, and the climate signal is stronger in older trees. This conclusion may be helpful in understanding the relationship between tree age and climate–growth response in Qilian juniper. Furthermore, compared with some other studied species, such as Larix decidua (Carrer and Urbinati 2004), age-controlled climate response in Qilian juniper is relatively weak,so we still think Qilian juniper is an ideal tree species for dendroclimatic reconstruction. But the ecology of tree-ring growth needs to receive more attention when the tree rings are used in extracting past climate signals (Zhang and Shao 2007). Particularly the study of aging processes in trees is itself at a youthful stage and there still remain uncertainties about the quantitative impacts of tree’s aging on climate–growth relationships (Bond 2000).
References
Blasing MN, Solomon AM, Duvick DN (1984) Response functions revisited. Tree-Ring Bull 44:252–256
Bond BJ (2000) Age-related changes in photosynthesis of woody plants. Trends Plant Sci 5:349–353
Bradley RS, Jones PD (1992) Climate since A.D. 1500. Routledge, London
Briffa K, Cook ER (1990) Methods of response function analysis. In: Cook ER, Kairiukstis LA (eds) Methods of dendrochronology applications in the environmental sciences. Kluwer Academic Publishers, Dordrecht, pp 240–247
Carrer M, Urbinati C (2004) Age-dependent tree-ring growth responses to climate in Larix declidua and Pinus cembra. Ecology 85:730–740
Chhin S, Wang G, Geoff, Tardif J (2004) Dendroclimatic analysis of white spruce at its southern limit of distribution in the spruce woods provincial park. Manitoba, Canada. Tree-Ring Res 60:31–43
Clack SL, Hallgren SW (2004) Age estimation of Quercus marilandica and Quercus stellata: applications for interpreting stand dynamics. Can J Forest Res 34: 1353–1358
Colenutt ME, Luckman BH (1995) The dendrochronological characteristics of alpine larch. Can J Forest Res 25:77–789
Cook ER (1985) A time series approach to tree-ring standardization. Dissertation, University of Arizona, Tucson, Arizona
D’Arrigo RD, Jacoby GC, Free RM (1992) Tree-ring width and maximum latewood density at the Northern American tree line: parameters of climatic change. Can J Forest Res 22:1290–1296
Day ME, Greenwood MS, Diaz-Sala C (2002) Age-and size-related trends in woody plant shoot development; regulatory pathways and evidence for genetic control. Tree Physiol 22:507–513
England JR, Attiwill PM (2006) Changes in leaf morphology and anatomy with tree age and height in the broadleaved evergreen species, Eucalyptus regnans F. Muell. Trees 20:79–90
Ettl GJ, Peterson DL (1995) Extreme climate and variation in tree growth: individualistic response in subalpine fir (Abies lasiocarpa). Global Change Biol 1:231–241
Fritts HC (1976) Tree rings and climate. Academic Press, London
Fritts HC (1996) Quick help for PRECON now called preconk version 5.11
Graumlich LJ (1991) Subalpine tree growth, climate, and increasing CO2: an assessment of recent growth trends. Ecology 72:1–11
Holmes RL (1983) Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bull 43:69–75
Holmes RL (1992) Dendrochronology program library user’s manual. Laboratory of Tree-Ring Research, University of Arizona, Tucson, Arizona
Huang JG, Zhang QB (2007) Tree rings and climate for the last 680 years in Wulan area of northeastern Qinghai-Tibetan Plateau. Climatic Change 80:369–377
Hubbard RM, Bond BJ, Ryan MG (1999) Evidence that hydraulic conductance limits photosynthesis in old Pinus ponderosa trees. Tree Physiol 19:165–172
Jacoby GC, Cook ER (1981) Past temperature variations inferred from a 400-year tree-ring chronology from Yukon Territory. Canada, Arct Antarct Alp Res 13:409–418
Kolb TE, Stone JE (2000) Differences in leaf gas exchange and water relations among species and tree sizes in an Arizona pine–oak forest. Tree Physiol 20:1–12
Liu XH, Qin DH, Shao XM, Chen T, Ren JW (2005) Temperature variations recovered from tree-rings in the middle Qilian Mountain over the last millennium. Sci China Ser D 48:521–529
Nash TH, Fritts HC, Stokes MA (1975) A technique for examing non-climatic variation in widths of annual tree rings with special reference to air pollution. Tree-Ring Bull 35:15–24
Rolland C (1993) Tree-ring and climate relationships for Abies alba in the internal Alps. Tree-Ring Bull 53:1–11
Ryan MG, Binkley D, Fownes JH (1997) Age related decline in forest productivity: pattern and process. Adv Ecol Res 27:213–262
Ryan MG, Yoder BJ (1997) Hydraulic limits to tree height and tree growth. BioScience 47:235–242
Shao XM, Huang L, Liu HB, Liang EY, Fang XQ, Wang LL (2005) Reconstruction of precipitation variation from tree rings in recent 1000 years in Delingha, Qinghai. Sci China Ser D 48:939–949
Stokes MA, Smiley TL (1968) An introduction to tree-ring dating. The University of Chicago Press, Chicago
Szeicz JM, MacDonald GM (1994) Age-dependent tree-ring growth responses of subarctic white spruce to climate. Can J Forest Res 23:120–132
Szeicz JM, MacDonald GM (1995) Dendroclimatic reconstruction of summer temperatures in northwestern Canada since A.D. 1638 based on age-dependent modeling. Quaternary Res 44:257–266
Travis DJ, Meentemeyer V, Belanger RP (1990) Stressed trees produce a better climatic signal than healthy trees. Tree-Ring Bull 50:29–32
Wu XD (1990) Tree-ring and climate change. Meteorology Press, Beijing (in Chinese)
Yoder BJ, Ryan MG, Waring RH, Schoettle AW, Kaufmann MR (1994) Evidence of reduced photosynthetic rates in old trees. Forest Sci 40:513–527
Zhang QB, Cheng GD, Yao TD, Kang XC, Huang JG (2003) A 2326-year tree-ring record of climate variability on the northeastern Qinghai-Tibetan plateau. Geophys Res Lett 30:1739–1742
Zhang QB, Qiu HY (2007) A millennium-long tree-ring chronology of Sabina przewalskii on northeastern Qinghai-Tibetan Plateau. Dendrochronologia 24:91–95
Zhang QB, Shao XM (2007) Tree Rings and Ecology: The 7th International Conference on Dendrochronology. J Integr Plant Biol 49:129–130
Zhang ZH, Wu XD (1997) Reconstruction of past 700 years climate change of the Qilian Mountains region from the tree ring data. Chin Sci Bull 42: 849–851
Acknowledgments
This study was jointly funded by the Chinese Academy of Sciences project KZCX1-10 and the National Natural Science Foundation of China project 40631002 and 90102009. We are grateful to Dr. Qi-Bin Zhang for his help in the research, to Dr. Deliang Zhang for many valuable suggestions on the early version of the manuscript. We also appreciate the constructive comments of two anonymous referees.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by J. Carlson.
Rights and permissions
About this article
Cite this article
Yu, G., Liu, Y., Wang, X. et al. Age-dependent tree-ring growth responses to climate in Qilian juniper (Sabina przewalskii Kom.). Trees 22, 197–204 (2008). https://doi.org/10.1007/s00468-007-0170-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00468-007-0170-y