
Abstract
Moisture availability has the potential to affect tropical forest productivity at scales ranging from leaf to ecosystem. We compared data for leaf photosynthetic, chemical and structural traits of canopy trees, litterfall production and seasonal availability of soil water at four sites across a precipitation gradient (1,800–3,500 mm year−1) in lowland Panamanian forest to determine how productivity at leaf and ecosystem scales may be related. We found stronger seasonality in soil water potential at drier sites. Values were close to zero at all sites during the wet season and varied between a minimum of −2.5 MPa and −0.3 MPa at the driest and wettest sites, respectively, during the dry season. Leaf photosynthesis and nitrogen concentration decreased with increasing precipitation, whereas leaf thickness increased with increasing precipitation. Leaf toughness and fiber/N ratios increased with increasing precipitation indicating reduced nutritional content and palatability with precipitation. Seasonality of litter production and quality decreased with increasing precipitation, but the amount of litterfall produced was not substantially different among sites. It appears that in Neotropical forest, moisture availability is associated with leaf photosynthetic and defensive traits that influence litterfall timing and quality. Therefore, variation in leaf physiological traits has the potential to influence decomposition and nutrient cycling through effects on litter quality.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
One of the ways to understand responses of tropical plants to water availability is to investigate patterns of leaf physiology and productivity along natural precipitation gradients. Water availability can have substantial effects on the type of leaves that species produce. In the lowland tropics, canopies of seasonally dry forest tend to be dominated by dry-season deciduous species, whereas canopies of wet forest are usually dominated by evergreen species (Santiago et al. 2004 Vázquez and Givnish 1998). A major proportion of matter and energy flowing through terrestrial ecosystems is cycled by the production, maturation, senescence and eventual abscission of leaves. Therefore, patterns of canopy leaf physiology and chemistry may be predictive of processes occurring on the ecosystem scale, such as litterfall production and the decomposition characteristics of litter. The purpose of this synthesis is to investigate whether leaf physiological traits of canopy trees along a precipitation gradient in lowland Panama are related to litter production and quality and whether this relationship has the potential to shape nutrient cycling dynamics.
Water availability has the potential to strongly affect productivity at both leaf and ecosystem scales in tropical forest. At the leaf scale, seasonal water deficit may restrict carbon gain in dry forest, favoring drought deciduous species (Eamus 1999; Sobrado 1991). At the ecosystem scale, recent studies of variation in aboveground net primary productivity (ANPP) with mean annual precipitation suggest that although ANPP increases with precipitation up to 2,500 mm year−1, at sites with higher rainfall, water availability begins to exceed biological demand and ANPP levels off or declines (Clark et al. 2001; Schuur 2003; Schuur and Matson 2001). The potential mechanisms for this pattern include increased rates of nutrient leaching, reduced oxygen availability through soil waterlogging and reduced light availability associated with clouds (Graham et al. 2003; Santiago et al. 2000; Schuur and Matson 2001). Litter decomposability and quality also appear to decline with increasing precipitation in tropical ecosystems (Austin and Vitousek 2000; Schuur 2001; Vitousek et al. 1994). Therefore, litter may be an important link between canopy leaves and soil-mediated ecosystem processes.
Understanding how leaf photosynthetic characteristics vary with precipitation and how those characteristics are related to litter quality may help place decomposition in an evolutionary context. For example, species from nutrient-poor habitats tend to produce leaves with low photosynthetic rates that are slow to decompose (Chapin 1980). There is also evidence that litter quality is related to variation in chemical and structural leaf traits (Cornelissen and Thompson 1997; Grime et al. 1996); litter decomposability can often be predicted by lignin concentration, which slows decomposition and the release of mineral nutrients (Melillo et al. 1982). However, little is known about how leaf physiological traits are related to litter quality and ecosystem processes (Santiago 2002). We propose that understanding the evolutionary trade-offs that lead to selection for deciduous canopy species in seasonally dry tropical forest and evergreen canopy species in wet tropical forest may allow prediction of decomposition dynamics and nutrient cycling based on physiological characteristics that govern plant carbon gain.
This study addresses how canopy tree species in lowland forest respond physiologically and biochemically to water availability, and how these characteristics are related to patterns of litterfall production and litter quality. Our main objectives were to: (1) determine patterns of leaf photosynthetic capacity and structural defenses along a precipitation gradient; (2) investigate whether photosynthetic capacity is related to litterfall production; and (3) explore the consequences and implications of photosynthetic allocation patterns for litterfall productivity and litter quality.
Study site and water availability
The study was conducted along a precipitation gradient extending across the Isthmus of Panama in protected forest surrounding the Panama Canal (Fig. 1). The precipitation gradient extends from the relatively dry Pacific coast, which receives 1,800 mm of precipitation annually to the Caribbean coast, where annual precipitation is >4,000 mm in some areas. Four 1-ha forest dynamics plots maintained by the Center for Tropical Forest Science (CTFS) were used to cover a range of mean annual precipitation from 1,800 mm to 3,500 mm (Table 1). All sites have a mean monthly precipitation >100 mm during the wet season between May and December, but dry season length varies twofold between the driest and wettest sites (Condit 1998) (Table 1). In addition to variation in amount of precipitation, there is also variation in the length and intensity of the dry season, which can be illustrated by variation in soil water availability at the four sites (Santiago et al. 2004; Fig. 2). During the wet season, when all sites receive abundant precipitation, soil water potential is near zero at all sites along the gradient. However, during the dry season, sites receiving less precipitation exhibit lower soil water potential, suggesting that minimum soil water potential plays a role in distinguishing among sites in the context of canopy phenology, leaf chemistry and photosynthesis. Other climatic factors such as relative humidity and light may also influence leaf physiological traits and are related to precipitation inputs (Rand and Rand 1982). All sites are lowland forest with altitude <400 m and mean annual temperatures of approximately 26°C (Dietrich et al. 1982; Santiago 2003).

The location of study sites along the Panama Canal, Republic of Panama, showing isohyets (mm) of mean annual precipitation

Time series of soil water potential (Ψsoil) from four lowland forest study plots in the Panama canal zone determined between 15 cm and 20 cm depth (n=6). J F M A M J J A S O N D J F January February March April May June July August September October November December January February
Soils in the Panama Canal forests are rich in clay and nitrogen and poor in phosphorus and potassium (Dietrich et al. 1982; Yavitt et al. 1993). Soils in this area tend to be well-drained (Kursar et al. 1995), suggesting that low redox potential is not a strong factor in reducing plant productivity. A heterogeneous mosaic of geological formations constitutes the parent material of the Panama Canal watershed. The 3,100-mm site is located on sedimentary parent material, whereas the other three sites in this study lie on volcanic parent material (Pyke et al. 2001; Woodring et al. 1980).
Species composition and phenology change rapidly across this gradient; there is a decrease in the proportion of dry season deciduous species with increasing precipitation (Table 2). Species richness of trees with a stem diameter ≥10 cm increases with precipitation, from 36 species at the 1,800-mm site to 162 species at the 3,500-mm site (Pyke et al. 2001; Santiago et al. 2004). The increase in species richness with increasing precipitation in the tropics is a well-documented pattern (Gentry 1988); recent analyses from the Guiana Shield and the Amazon Basin suggest that increasing dry season length limits the number of species that can survive under a given climatic regime (ter Steege et al. 2000). All study plots are located in mature forest (>200 year), except the 1,800-mm site, which is a forest of mixed age (70–100 year). Measurements on the largest canopy trees at the 1,800-mm site are comparable to other sites even though this forest is younger, because canopy composition was representative of mature seasonally dry forest (Croat 1978).
Canopy photosynthetic characteristics
Photosynthesis was measured on canopy sun leaves throughout the wet seasons of 2000 and 2001. The eight most common canopy species at each site were determined by contribution to stand basal area and used for measurements of photosynthesis. Two to four individuals of each species (15 leaves per species at a site) were measured with an oxygen electrode (Björkman and Demmig 1987; Delieu and Walker 1981). Maximum photosynthetic oxygen evolution rates per unit mass decreased with increasing precipitation (Fig. 3a). Area-based maximum photosynthetic rates also significantly decreased with increasing precipitation, but the relationship was not as strong as mass-based measurements (r2=0.13; P=0.04; Santiago et al. 2004). Leaf nitrogen concentration per unit mass decreased with increasing precipitation (Fig. 3b) and was a good predictor of mass-based photosynthetic rate (r2=0.78; P<0.0001; Santiago et al. 2004). Decreasing leaf nitrogen per unit mass with increasing precipitation has also been reported along precipitation gradients in Australia and is thought to promote higher water use efficiency in drier sites (Wright et al. 2001; Wright and Westoby 2002).

Relationship between mean annual precipitation and maximum net photosynthetic rate per unit mass (a) and leaf nitrogen concentration (b) for the eight most abundant canopy tree species in four Panamanian lowland forests
Variation in canopy photosynthesis and leaf nitrogen concentration has the potential to influence litter quality and nutrient cycling following senescence. Nitrogen retranslocation efficiency (proportion of leaf nitrogen resorbed) varied from 34% to 68% with a mean of 50% for 12 canopy species at the 3,100-mm site (Santiago 2003). Investigations from Australia found that nitrogen resorption did not differ with site rainfall at a given level of soil nutrient availability and support the concept that selection to minimize nutrient losses drives nutrient concentrations of senesced leaves, rather than proportional resorption (Killingbeck 1996; Wright and Westoby 2003). Therefore, it is likely that retranslocation efficiency falls within similar ranges across this precipitation gradient, and that variation in photosynthesis and leaf nitrogen translate into litter quality by determining litter nitrogen concentration and potentially decomposition.
Minimum soil water potential explained 39% of variation in maximum photosynthetic rate per unit mass (Fig. 4a). Photosynthetic rates decreased from a range of 130.8–289.8 nmol g−1 s−1 at the site with the largest seasonal water deficit to 58.1–161.5 nmol g−1 s−1 at the site with the most aseasonal water regime. Mean dry season length explained 30% of variation in mass-based maximum photosynthesis (Fig. 4b), illustrating the relationship between seasonality and maximum photosynthetic rate. Because seasonally dry tropical forest is characterized by deciduous vegetation in contrast to evergreen species in wet forest, leaf life span is expected to be shorter and photosynthetic rate higher in seasonally dry forest than in wet forest (Reich et al. 1997) resulting in a shift in the carbon fixation strategy of plants with precipitation (Wright et al. 2001). Although shifts in leaf photosynthetic traits along tropical precipitation gradients may have been predicted based on existing theory (Kikuzawa 1991), to our knowledge this data set represents the first examination of photosynthesis of tropical tree species along a broad precipitation gradient.

Relationship between maximum net photosynthetic rate and minimum average soil water potential (a) and mean dry season length (b) for the eight most abundant canopy tree species in four Panamanian forests on a precipitation gradient
Canopy leaf structure and chemistry
Leaf structural and chemical traits were evaluated on from two to four individuals (15 fresh leaves per species at a site) of the eight most common canopy species at each site to determine whether variation along a range in precipitation is predictive of litter quality. Lamina thickness increased with increasing precipitation (Table 3; r2=0.30; P=0.001; Santiago et al. 2004), and in part explains why mass-based photosynthetic rates and nitrogen concentrations of canopy leaves decrease with increasing precipitation more strongly than area-based photosynthetic rates and nitrogen concentrations (Santiago et al. 2004). Leaf toughness also increased with increasing precipitation (Table 3; r2=0.29; P<0.005; Santiago et al. 2004) suggesting that evergreen species in wet forest allocate more to structural leaf defenses. Several explanations for increases in plant diversity with increasing precipitation in lowland tropical forest include increased herbivore and pathogen pressure with increasing precipitation, since aseasonally humid environments favor plant pests (Givnish 1999; Wolda 1988; Wright 1992). Therefore, increasing leaf toughness of canopy species along this precipitation gradient may reflect variation in herbivore pressure with increasing precipitation in addition to the expected increase in structural defense associated with evergreenness (Coley et al. 1985). Increasing leaf toughness with increasing precipitation may also indicate that litter produced by evergreen species in wet forest may be more structurally resistant to decomposition than litter derived from relatively thin leaves of dry forest species.
Canopy leaves are an important food source for animal communities in tropical forest (Milton 1982), thus chemical and structural leaf adaptations to various precipitation regimes may also be indicative of food quality. We observed higher fiber/nitrogen ratios from canopy species at the two wetter sites (Table 3). High leaf fiber concentrations per unit nitrogen reflect relatively low nutritional content because fiber is undigestible by most insects (Hochuli 1996) and is negatively correlated with herbivory on mature leaves in Panamanian lowland forest (Coley 1983). Therefore, decreasing food quality of canopy leaves with increasing precipitation supports the prediction that herbivore and pathogen pressure is higher in wetter tropical forest. Currently, few studies address how leaf chemistry influences energy flow to herbivores and decomposers simultaneously. However, several reports suggest that antiherbivore defenses may continue to function against decomposers during decomposition (Grime et al. 1996; Wardle et al. 2002). Patterns of leaf physiology, chemistry and structural defenses identified across this precipitation gradient suggest that lower food quality and higher allocation to structural defenses in wetter forest may contribute to reduced litter quality.
Litter production and quality
Aboveground production of leaf litter accounts for approximately 25–35% of annual net primary productivity in forest ecosystems (Schlesinger 1991), and represents an important pathway connecting canopy leaves to ecosystem processes such as decomposition and nutrient cycling. We measured litter production using 10- 0.25-m2 litter traps randomly distributed in each 1-ha study plot. Litter was collected 9–11 times at intervals ranging from 1 week to 12 weeks for the period between February 2001 and February 2002. Litter used in chemical analyses was collected at intervals of 7–10 days in February–June 2001, October 2001, and February 2002. Litter collected at intervals >2 weeks were adjusted for mass loss within traps using decomposition data from the 3,100-mm site (Santiago 2003). Leaf litterfall accounted for 59–76% of total litterfall in this study (Table 4), and therefore connects canopy leaves to soil nutrient pools. Although we observed no significant variation in total litterfall production among sites in this study (F=1.96; P>0.1), we found that dry-season deciduous species influenced the seasonality of litter production (Fig. 5). At the drier sites, we observed a pulse of leaf drop during the February dry season peak, and a gradual decline in leaf litter production rates during the wet season (May–November). However, at the wettest site, there was little annual variation in litterfall production (Fig. 5) as evergreen species dominate and produce litter more evenly throughout the year.

Time series of seasonality in leaf litterfall rate on four 1-ha lowland forest sites in Panama that vary in mean annual precipitation (n=10). yr Year, wk week
Nitrogen fluxes in leaf litterfall highlight the importance of this nutrient pathway. Between 7.30 g m−2 year−1 and 9.07 g m−2 year−1 of nitrogen was returned to the forest floor through leaf litter (Table 4). These values fall in a similar range to previously reported nitrogen fluxes from reviews of tropical forest nutrient cycling (Leigh and Windsor 1982; Vitousek and Sanford 1986). Based on the review of Leigh and Windsor (1982), leaf litter contains approximately 75% of nitrogen in total litterfall. Therefore, leaf allocation patterns of canopy trees have a substantial bearing on how nitrogen is returned to the soil pool. For example, if leaf litter is highly lignified (low litter quality), then leaf litter nitrogen will enter the relatively recalcitrant pool of soil organic matter and will become available to plants at lower rates (Vitousek et al. 1994). Alternatively, if leaf litter is composed of relatively labile carbon, nitrogen in leaf litter will be released to the plant available pool at higher rates.
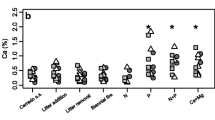
The cumulative effects of allocation to leaf physiological and structural traits on decomposition and nutrient cycling of senesced leaves is reflected by litter quality. We observed significantly higher lignin concentration of leaf litter at wetter sites (F=11.76; P<0.001; Fig. 6), suggesting that interactions between climate, species composition and physiology result in the production of poorer quality litter with increasing precipitation. Litter quality is presumed to be the strongest determinant of decomposition in tropical forests since these systems are less likely to have decomposition rate limited by temperature and moisture, relative to other biomes (Meentemeyer 1978). In addition, studies of Metrosideros polymorpha, the dominant woody species in Hawaii, suggest that litter quality can be at least as important as site characteristics in determining leaf decomposition rates along precipitation gradients (Austin and Vitousek 2000; Schuur 2001; Vitousek et al. 1994). The results presented along this range of precipitation in lowland tropical forest suggest that interactions between climate and species composition influence the timing and quality of leaf litter production, but not necessarily total litterfall production. Further studies need to be initiated to determine the relative importance of litter quality and site characteristics in determining decomposition in lowland tropical forest (Vitousek et al. 1994).

Leaf litter lignin concentration of composite samples from four sites along a precipitation gradient in lowland tropical forest. (n=4)
Synthesis
Our data indicate that variation in photosynthetic, chemical and structural leaf traits of canopy tree species is related to amount and seasonality of precipitation in Central Panamanian lowland forest. In seasonally dry forest, drought deciduousness is more prevalent and species produce thin leaves with high photosynthetic potential, in contrast to sites with shorter, less intense dry seasons, which are dominated by relatively thick-leaved evergreen species. Leaf photosynthetic and defensive traits were also related to litter quality supporting the idea that leaf physiology and defense in tropical forest may vary along an axis that is also predictive of litter decomposition rate and mineral nutrient release. Therefore, precipitation may regulate plant community composition and productivity directly through water availability, and indirectly through effects on litter quality. The connection between photosynthetic capacity and litter quality under natural conditions in lowland tropical forest suggest that integration of plant physiological ecology and ecosystem science may prove fruitful in augmenting our understanding of the effects of plant species and communities on ecosystem processes.
References
Austin AT, Vitousek PM (2000) Precipitation, decomposition and litter decomposability of Metrosideros polymorpha in native forests on Hawai’i. J Ecol 88:129–138
Björkman O, Demmig B (1987) Photon yield of O2 evolution and chlorophyll fluorescence characteristics at 77 K among vascular plants of diverse origins. Planta 170:489–504
Chapin FS III (1980) The mineral nutrition of wild plants. Annu Rev Ecol Syst 11:233–260
Clark DA, Brown S, Kicklighter DW, Chambers JQ, Thomlinson JR, Ni J, Holland EA (2001) Net primary production in tropical forests: an evaluation and synthesis of existing field data. Ecol Appl 11:371–384
Coley PD (1983) Herbivory and defensive characteristics of tree species in a lowland tropical forest. Ecol Monogr 53:209–233
Coley PD, Bryant JP, Chapin FS III (1985) Resource availability and plant anti-herbivore defense. Science 230:895–899
Condit R (1998) Ecological implications of changes in drought patterns: shifts in forest composition in Panama. Climate Change 39:413–427
Cornelissen JHC, Thompson K (1997) Functional leaf attributes predict litter decomposition rate in herbaceous plants. New Phytol 135:109–114
Croat TB (1978) Flora of Barro Colorado Island. Stanford University Press, Stanford, Calif.
Delieu T, Walker D (1981) Polarographic measurement of photosynthetic oxygen evolution by leaf-disks. New Phytol 89:165–178
Dietrich WE, Windsor DM, Dunne T (1982) Geology, climate, and hydrology of Barro Colorado Island. In: Leigh EG, Rand AS, Windsor DM (eds) The ecology of a tropical forest. Smithsonian Institution Press, Washington, D.C., pp 21–46
Eamus D (1999) Ecophysiological traits of deciduous and evergreen woody species in the seasonally dry tropics. Trends Ecol Evol 14:11–16
Gentry AH (1988) Changes in plant community diversity and floristic composition on environmental and geographical gradients. Ann Mo Bot Gard 75:1–34
Givnish TJ (1999) On the causes of gradients in tropical tree diversity. J Ecol 87:193–210
Graham EA, Mulkey SS, Kitajima K, Phillips NG, Wright SJ (2003) Cloud cover limits net CO2 uptake and growth of a rainforest tree during tropical rainy seasons. Proc Natl Acad Sci U S A 100:572–576
Grime JP, Cornelissen JHC, Thompson K, Hodgson JG (1996) Evidence of a causal connection between anti-herbivore defence and the decomposition rate of leaves. Oikos 77:489–494
Hochuli DF (1996) The ecology of plant/insect interactions—implications of digestive strategy for phytophagous insects. Oikos 75:133–141
Kikuzawa K (1991) A cost–benefit-analysis of leaf habit and leaf longevity of trees and their geographic pattern. Am Nat 138:1250–1263
Killingbeck KT (1996) Nutrients in senesced leaves: keys to the search for potential resorption and resorption proficiency. Ecology 77:1716–1727
Kursar TA, Wright SJ, Radulovich R (1995) The effects of the rainy season and irrigation on soil water and oxygen in a seasonal forest in Panama. J Trop Ecol 11:497–516
Leigh EG, Windsor DM (1982) Forest production and regulation of primary consumers on Barro Colorado Island. In: Leigh EG, Rand AS, Windsor DM (eds) The ecology of a tropical forest. Smithsonian Institution Press, Washington, D.C., pp 111–122
Meentemeyer V (1978) Microclimate and lignin control of litter decomposition rates. Ecology 59:465–472
Melillo JM, Aber JD, Muratore JF (1982) Nitrogen and lignin control of hardwood leaf litter decomposition dynamics. Ecology 63:621–626
Milton K (1982) Dietary quality and demographic regulation in a howler monkey population. In: Leigh EG, Rand AS, Windsor DM (eds) The ecology of a tropical forest. Smithsonian Institution Press, Washington, D.C., pp 273–290
Pyke CR, Condit R, Aguilar S, Lao S (2001) Floristic composition across a climatic gradient in a neotropical lowland forest. J Veg Sci 12:553–566
Rand AS, Rand WM (1982) Variation in rainfall on Barro Colorado Island. In: Windsor DM (ed) The ecology of a tropical forest. Smithsonian Institution Press, Washington, D.C., pp 47–59
Reich PB, Walters MB, Ellsworth DS (1997) From tropics to tundra: global convergence in plant functioning. Proc Natl Acad Sci U S A 94:13730–13734
Santiago LS (2002) Leaf photosynthesis and chemistry predict leaf decomposition rate in the lowland wet forest of Fort Sherman, Panama. In: Proceedings of the Association for Tropical Biology Annual Meeting, Panama City, Republic of Panama
Santiago LS (2003) Leaf traits of canopy trees on a precipitation gradient in Panama: Integrating plant physiological ecology and ecosystem science. PhD dissertation. University of Florida, Gainesville, Fla.
Santiago LS, Kitajima K, Wright SJ, Mulkey SS (2004) Coordinated changes in photosynthesis, water relations and leaf nutritional traits of canopy trees along a precipitation gradient in lowland tropical forest. Oecologia 139:495–502
Santiago LS, Goldstein G, Meinzer FC, Fownes J, Mueller-Dombois D (2000) Transpiration and forest structure in relation to soil waterlogging in a Hawaiian montane cloud forest. Tree Physiol 20:673–681
Schlesinger WH (1991) Biogeochemistry: an analysis of global change. Academic Press, New York
Schuur EAG (2001) The effect of water on decomposition dynamics in mesic to wet Hawaiian montane forests. Ecosystems 4:259–273
Schuur EAG (2003) Productivity and global climate revisited: the sensitivity of tropical forest growth to precipitation. Ecology 84:1165–1170
Schuur EAG, Matson PA (2001) Net primary productivity and nutrient cycling across a mesic to wet precipitation gradient in Hawaiian montane forest. Oecologia 128:431–442
Sobrado MA (1991) Cost–benefit relationships in deciduous and evergreen leaves of tropical dry forest species. Funct Ecol 5:608–616
ter Steege H, Sabatier D, Van Castellanos H, Andel T, Duivenvoorden J, De Oliveira AA, Ek R, Lilwah R, Maas P, Mori S (2000) An analysis of the floristic composition and diversity of Amazonian forests including those of the Guiana Shield. J Trop Ecol 16:801–828
Vázquez JA, Givnish T (1998) Altitudinal gradients in tropical forest composition, structure, and diversity in the Sierra de Manantlán. J Ecol 86:999–1020
Vitousek PM, Sanford RL (1986) Nutrient cycling in moist tropical forest. Annu Rev Ecol Syst 17:137–167
Vitousek PM, Turner DR, Parton WJ, Sanford RL (1994) Litter decomposition on the Mauna Loa environmental matrix, Hawaii: patterns, mechanisms, and models. Ecology 75:418–429
Wardle DA, Bonner KI, Barker GM (2002) Linkages between plant litter decomposition, litter quality, and vegetation responses to herbivores. Funct Ecol 16:585–595
Wolda H (1988) Insect seasonality: why? Annu Rev Ecol Syst 19:1–18
Woodring W, Stewart R, Stewart J (1980) Geologic map of the Panama canal. U.S. Geological Survey, Reston, Va.
Wright SJ (1992) Seasonal drought, soil fertility and the species density of tropical forest plant communities. Trends Ecol Evol 7:260–263
Wright IJ, Westoby M (2002) Leaves at low versus high rainfall: coordination of structure, lifespan and physiology. New Phytol 155:403–416
Wright IJ, Westoby M (2003) Nutrient concentration, resorption and lifespan: leaf traits of Australian sclerophyll species. Funct Ecol 17:10–19
Wright IJ, Reich PB, Westoby M (2001) Strategy shifts in leaf physiology, structure and nutrient content between species of high- and low-rainfall and high- and low-nutrient habitats. Funct Ecol 15:423–434
Yavitt JB, Wieder RK, Wright SJ (1993) Soil nutrient dynamics in response to irrigation of a Panamanian tropical moist forest. Biogeochemistry 19:1–25
Acknowledgements
We would like to thank Missy Holbrook and Marco Gutiérrez for organizing the symposium and inviting this paper, Jeff Chambers, David Burslem, Saara deWalt, William Grauel, Chris Mullins, Antonio Vázquez and Amy Zanne for stimulating discussions during the conference, and Ted Schuur, Katia Silvera, and Kevin Simonin for comments on the ideas presented here. Funding was provided by an EPA STAR Graduate Fellowship to L. S. Santiago
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Santiago, L.S., Mulkey, S.S. Leaf productivity along a precipitation gradient in lowland Panama: patterns from leaf to ecosystem. Trees 19, 349–356 (2005). https://doi.org/10.1007/s00468-004-0389-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00468-004-0389-9