
Abstract
Improvement of light penetration within tree canopies has been a constant objective of fruit tree architecture manipulation through the setting up of training systems. Recently, centrifugal training, i.e. the removal of fruiting shoots in the tree centre and on the underside of branches, has been proposed to improve fruit size and colour as well as return-bloom as compared to conventional solaxe-trained trees with equivalent crop loads. The present study was conducted to quantify the benefits of centrifugal training on light interception by the fruiting shoots via computer-assisted three-dimensional representations of foliage geometry. Data were collected on six 5-year-old apple trees cv.Galaxy, trained either with solaxe or centrifugal training systems, using an electromagnetic 3D digitiser. The 3D distribution of the foliage in the tree canopy was recreated by combining both the spatial locations of shoots (as measured from 3D digitising) and foliage reconstruction. Light interception efficiency properties of the trees were characterised by silhouette to total area ratio (STAR) values computed from images of the 3D mock-ups. Compared to the solaxe system, centrifugal training significantly improved the STAR of the whole tree by 20%. It also increased both leaf area and STAR of the fruiting shoots by approximately 15%, regardless of their position in the canopy. In this paper, we discuss the role of this enhanced light interception by the canopy in increasing the autonomy of the fruiting shoot, i.e. improved fruit size and colour, and return-bloom.
Similar content being viewed by others



Avoid common mistakes on your manuscript.
Introduction
Light distribution within tree canopies depends on various inter-related factors. One factor is the intrinsic architectural pattern of the cultivar itself (Sansavini and Corelli-Grappadelli 1992), from upright highly branched scaffolds to low-branched tip-bearing cultivars (Lespinasse and Delort 1986). For a given cultivar, the rootstock also plays a major role via the canopy volume that confers on the cultivar (Mierowska et al. 2002; Palmer 1980). A second factor is the planting system (Jackson 1980) which includes both tree arrangement in the orchard (planting distances, row orientation) and training of the tree canopy by pruning and bending procedures (tree shape and height). Training partly defines the proportion and distribution in space of the various shoot categories, long versus short shoots and vegetative versus fruiting shoots. Since whole-tree photosynthesis is primarily light limited (Lakso 1980), manipulation of canopy architecture to enhance global light interception is a major objective of all planting systems (Lakso and Corelli-Grappadelli 1992; Palmer 1980; Sansavini and Corelli-Grappadelli 1992; Tustin et al. 1988, 1998; Wagenmakers et al. 1991).
The contribution of the various shoot categories to tree productivity is now well documented and, as a general trend, whole-tree yield has been shown to strongly depend on light interception by the spur canopy (Corelli-Grappadelli et al. 1994; Lakso et al. 1999; Wünsche et al. 1996; Wünsche and Lakso 2000). This probably arises from both morphological features of short as compared to long shoots, the former having higher leaf to shoot ratios (Lauri and Kelner 2001), and faster carbon exportation capacity early in the season (Sansavini and Corelli-Grappadelli 1992). These results clearly illustrate the advantages of managing well-illuminated spur-rich canopies with high porosity to light (Lakso 1994; Sansavini and Corelli-Grappadelli 1992).
Improvement of light distribution and interception in the orchard can take place at two levels: management of discontinuities between tree crowns which depends on tree shape and planting distances (Tustin et al. 1998), and development of training procedures which decrease within-tree shading. The latter factor has not been adequately studied and is more difficult to manage, and training systems which typically aim at increasing light interception by the canopy—e.g. Lincoln Canopy and, to a lesser extent, V-trellis and Y-trellis—often result in excessive annual, vertical growth on the exposed sides of scaffolds (Palmer and Warrington 2000). Unless these shoots are removed during the growing season, they increase shading on fruiting spurs located lower down. An improvement of training systems should therefore integrate not only the management of tree height and shape and discontinuities between trees, but also a better knowledge of the tree reactions to training methods, specifically shoot orientation and pruning.
For several years now in France, the thinning of flowering spurs—referred to as artificial extinction or just extinction—has been proposed as a complement and possibly an alternative to both conventional pruning and flower and fruitlet-thinning procedures to better control the balance between growth and fruiting and to enhance return-bloom on individual spurs (Lauri et al. 1997, 2004; Lauri and Lespinasse 1999). When carried out more specifically along and around the trunk and on the underside of fruiting branches of solaxe-trained trees (Lespinasse 1996), artificial extinction favours the peripheral layer, removes potentially shaded fruiting laterals and, thus, is aimed at improving light penetration within the tree (Larrive et al. 2000). This procedure (Fig. 1), referred to as centrifugal training, increases the proportion of large-sized well-coloured fruit (Crete et al. 2002; Ferre et al. 2002; Lauri et al. 2004). The research presented here examines and quantifies the influence of training procedures, i.e. original solaxe-trained trees without artificial extinction versus centrifugal trained-trees, on canopy morphology, light interception and light distribution in apple trees. For this purpose, three-dimensional (3D) representations of foliage geometry (Sinoquet et al. 1998) were created to assess canopy structure and light interception properties.

Top and lateral views of centrifugal training concept and light distribution (arrows) in the tree
Materials and methods
Plant material
GalaxyC.O.V. scions grafted onto Pajam 2 were planted in 1998 at the CEHM Experimental Station—Marsillargues, south eastern France (43°67′N; 4°18′E). They were spaced 4 m ×1.25 m (2,000 trees/ha), bent at 2.5 m and initially trained using the solaxe system. In spring 2001, the year of the first profuse flowering (more than 90% of laterals were flowering), a trial was initiated in a split-plot design (Tarisse 2001) to analyse the effects of different types of fruiting branch management on fruit quality and return-bloom over several years. The same fruit-load, i.e. four fruits/cm2 of fruiting branch cross-sectional area (FBCSA), considered as optimal for this orchard (MAFCOT 1999), was applied in three different types of training management defined as follows:
-
Crop load adjusted only by conventional chemical-thinning and hand-thinning of fruitlets on solaxe-trained trees, hereafter referred to as original solaxe (OS).
-
Crop load adjusted by artificial extinction to four fruiting laterals/cm2 FBCSA, with supplemental hand-thinning to leave one fruit per flower cluster, hereafter referred to as centrifugal-trained trees with one fruit per flower cluster (CT1).
-
Crop load adjusted by artificial extinction to two fruiting laterals/cm2 FBCSA, with two fruits per flower cluster, hereafter referred to as centrifugal-trained trees with two fruits per flower cluster (CT2).
In spring 2002, crop load was again adjusted according to each specific type of branch management, either by chemical-thinning and, if necessary, hand-thinning. On centrifugal-trained trees, only a little additional extinction was necessary to maintain the desired crop load.
Typology of annual shoots
Growing 2002 shoots were distinguished by both their type, i.e. vegetative versus bourse and the associated bourse-shoot(s), and their length, i.e. short versus long shoots. The threshold value of 5 cm, distinguishing annual shoots with very short internodes from those with longer ones, commonly found in literature (Lauri and Kelner 2001; Pratt 1990; Sansavini and Corelli-Grappadelli 1992), has been determined as an easy-to-use field discrimination method. The bourse itself always belongs to the short class. Five type-length combinations were then determined: vegetative short shoot (VS), vegetative long shoot (VL), long bourse-shoot, short bourse-shoot and bourse. In the latter case, the bourse and associated bourse-shoot(s) were separately measured and geometrically reconstructed, but they were gathered in the analyses of leaf area and light interception properties, since they both contribute significantly to the carbon budget of the growing fruit (Wünsche and Lakso 2000). Together, they were referred to as fruiting shoots (Fr).
Measurement of canopy structure
In order to have comparable data on trees, field work was carried out during a short phenological window, i.e. after first growth arrest, in southeast France usually at the end of May, and before fruit weight significantly changes shoot orientation. Due to digitising time duration, only two healthy trees per treatment were chosen for their homogeneity of height, volume and branch distribution along the trunk. For all current-year shoots in each tree, the above-defined shoot category and the spatial co-ordinates of the proximal and distal tips were recorded at the end of May 2002. Spatial co-ordinates were measured with an electromagnetic 3D digitiser (Fastrak, Polhemus, Cochester, Vt., USA) and POL95 software (Adam 1999), according to Sinoquet and Rivet (1997).
On a sample of 20–30 shoots per category, the number of leaves (Nl), the length of individual shoots (Ls), the length of petioles (Lp) and the length (Ll) and width (Wl) of each lamina on a shoot were measured with a ruler. In addition, each leaf was 3D-digitised in order to record location (i.e. spatial co-ordinates of the petiole–midrib junction point) and orientation at Euler angles (azimuth, elevation and roll), according to Sinoquet et al. (1998). Individual leaf area (Al) was measured with a LICOR 3100 area-meter. Allometric relationships were then determined at two levels from the data set of sampled shoots:
-
Individual leaf level: Lp as a function of Ll (Fig. 2a), Wl as a function of Ll (Fig. 2b) and Al as a function of Ll (Fig. 2c).
Fig. 2 
Allometric relationships between a petiole length, Lp, and lamina length, Ll, b lamina width, Wl, and lamina length, Ll, and c leaf area, Al, and the square of lamina length, Ll2. Data collected on 230 leaves of 5-year-old “Galaxy” trees. All correlations were best adjusted by linear regressions and were significant at P<0.05
-
Shoot level for each shoot type: Nl (Fig. 3a) and total shoot leaf area as a function of Ls (Fig. 3b).
Fig. 3 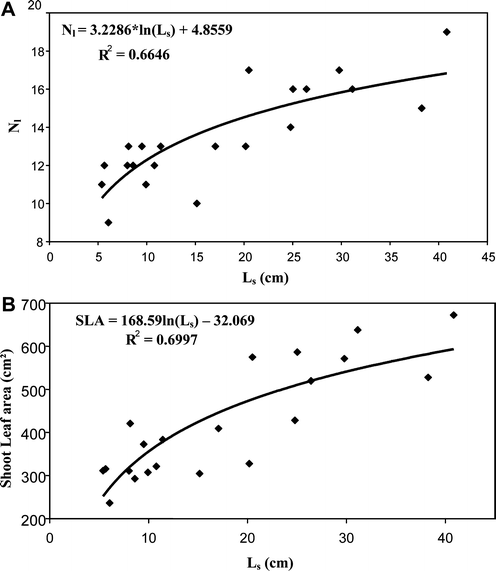
Examples of vegetative long (VL) shoots of allometric relationships between a leaf number, Nl, and length of individual shoot, Ls, b shoot leaf area and length of individual shoots, Ls. Data collected on 21 vegetative long shoots of 5-year-old “Galaxy” trees


All relationships further used for the foliage reconstruction were significant (P<0.05).
Foliage reconstruction and light interception computations
The 3D distribution of the foliage in the tree canopy was recreated by combining both the spatial locations of shoots (as measured from 3D digitising) and foliage reconstruction rules derived from the 3D-digitised shoots at leaf scale. For each shoot, the number of leaves and leaf area attached to the shoot were derived from the above-mentioned allometric relationships. All leaves in a shoot were assumed to have the same leaf area, equivalent to the ratio of shoot leaf area to number of leaves. Leaf length and width were derived from leaf area using allometric relationships at leaf scale. Spatial co-ordinates of the leaves were computed with the assumption of constant internode length. Midrib inclination and lamina rolling angle around the midrib were randomly sampled in the leaf angle distributions derived from the leaf-scale digitising data, while midrib azimuth was assumed to be uniformly distributed. The 3D reconstruction of the tree foliage was implemented in a computer program called MakeGalaxy where the input data are the spatial co-ordinates of the leafy shoots and the reconstruction rules, while the output file is the list of the tree leaves with their geometrical attributes, namely leaf dimensions, angles and spatial co-ordinates.
Light interception by the 3D-reconstructed trees was computed by using VegeSTAR 2.0 software (Adam et al. 2001). VegeSTAR uses the geometrical information inferred by the MakeGalaxy programme to synthesize virtual images of the trees. It then computes light interception by processing the virtual images of the 3D plant mock-ups. Indeed, leaf area viewed from any direction is the sunlit leaf area in this direction. The software computes the silhouette to total area ratio (STAR, Stenberg 1996), where the silhouette area is the projected leaf area in the incident light direction. The STAR value thus describes directional light interception by the canopy. In order to get sky-integrated indexes of light interception properties, the sky vault was divided into 46 solid angles according to the TURTLE sky division (Den Dulk 1989) and directional STAR values for the 46 directions were summed up after weighting according to the SOC (Standard OverCast) sky radiance distribution (Moon and Spencer 1942). The resulting STARSOC value characterises light interception for an overcast sky, which is assumed to be a correct approximation of light interception for any daily integration of light interception, i.e. with a clear or overcast sky (Sinoquet and Bonhomme 1992). Therefore, only STARSOC values were used in this article as integrated indexes of light interception efficiency of the trees.
The STAR value was calculated for each individual shoot in the tree, taking the presence of the other shoots into account. For this purpose, a different false colour was attributed to each shoot and plant images were processed by distinguishing each colour. STAR values at the scale of the shoot category (i.e. the set of shoots of the same category) and at whole-tree scale were inferred as an average STAR value, from values computed at shoot level. Finally, vertical profiles of STAR at shoot scale were computed by affecting fruiting shoots within 20-cm thick layers.
Data analysis
The data were analysed using Statistica software [StatSoft France (2003). STATISTICA, version 6. http://www.statsoft.com]. ANOVA followed by Newman–Keuls test was used to compare leaf area at shoot scale, and a Kolmogorov–Smirnov test adapted to non-normally distributed data when comparing STAR values and frequencies of shoots. No statistical tests were done on variables at tree scale (e.g. total number of leaves and shoots) because only two trees per treatment were measured.
Results
Canopy structure and demography
Tree dimensions, calculated from projections on horizontal and vertical plans of the farthest annual shoots (Table 1), confirmed that the selected trees had homogeneous crown extension.
Virtual pictures show the morphological characteristics of the trees trained with either the centrifugal or the solaxe system (Fig. 4). For instance, the presence of a central chimney, brought about by the extinction in the centre of the tree (Fig. 1), is noticeable in centrifugal-trained trees (Fig. 4a) but, as expected, not in OS trees (Fig. 4b).

Examples of top-sight reconstructed tree pictures drawn by VegeSTAR 2.0. a Centrifugal training with one fruit per flower cluster, tree #B (CT1-B), and b original solaxe, tree #A (OS-A)
However, shoot and leaf demography values greatly varied between trees, including within a given treatment. For example, the total number of leaves per tree varied globally from 4,447 to 6,541, and the two CT2 trees had 4,925 and 6,541 leaves (Table 2). The number of leaves per shoot category and number of shoots per tree could vary by twice as much: 654 leaves on 105 fruiting shoots on CT2-B, and 1,263 leaves on 218 fruiting points on OS-A. Due to artificial extinction, CT trees had less fruiting shoots and less leaves in this category than OS trees. The number of leaves in a category varied from 15% to 50% (Table 2), even between trees from a given treatment.
The total leaf area was less variable between the two trees in the OS treatment than in the CT treatments (Table 3) but, once again, there was no ranking between the different types of fruiting branch management. However, the tree CT1-A had fewer leaves, a higher proportion of fruiting shoots leaves (Table 2) and a smaller total leaf area (1,013.01 dm2) compared to the other trees (Table 3).
Individual shoot leaf area for VS and VL were significantly different from one tree to another but there was no consistent trend between treatments. On the other hand, individual leaf area for the fruiting shoots was more homogeneous within a treatment, and there was a significantly higher leaf area (approximately +15%) for the fruiting shoots on CT as opposed to OS trees (Table 3).
Light interception properties
CT trees had approximately 20% higher whole-tree mean STAR values than OS trees (Table 4). VL had higher STAR values, regardless of the treatment and the tree, than other shoot categories (Table 4). As a general trend, VS had lower STAR values than the corresponding tree STAR values (Table 4). In CT trees, Fr had approximately the same or a slightly higher STAR value than the whole tree it belonged to. Conversely, the STAR values of Fr in OS trees were more than 10% lower than the whole-tree mean STAR (Table 4).
OS trees had 15% lower STAR values on the fruiting shoots than CT trees (Table 4). Moreover, the distributions of STAR values of all shoot categories were more homogeneous in OS than in CT trees (Fig. 5), regardless of the height within the tree (Fig. 6).

Distribution of silhouette to total leaf area ratio (STAR) values on solaxe-trained and centrifugal-trained trees. Data merged for the two trees of each treatment. *Crossed non-parametric Kolmogorov–Smirnov tests. Different letters mean significant different values (P<0.05). OS original solaxe, CT1 centrifugal training with one fruit per flower cluster, CT2 centrifugal training with two fruit per flower cluster

Vertical profiles of the silhouette to total leaf area ratio (STAR) mean values (± SE) of fruiting shoots according to the height within the canopy (Z co-ordinates). OS original solaxe, CT1 centrifugal training with one fruit per flower cluster, CT2 centrifugal training with two fruit per flower cluster
Centrifugal training significantly increased STAR values of fruiting shoots, regardless of the height within the tree as compared to solaxe-trained trees (Fig. 6). However, there was no consistent difference in STAR mean values of the fruiting shoots between the top and bottom of trees for either training system (Fig. 6).
Discussion
This study provides further insight into the effects of centrifugal training on both leaf area development and light interception by the various shoot categories. The extinction procedure used in centrifugal training removes a certain proportion of either poorly developed or competitive fruiting shoots, with the aim of improving vegetative and fruit growth in the remaining ones. This study analysed the effects of such manipulations at two complementary levels.
The first level was the effect on leaf area of the various shoot categories. Centrifugal training significantly enhances leaf area of fruiting shoots, i.e. fruiting bourse and associated bourse-shoot(s). In centrifugal-trained trees, fruiting shoots may have benefitted from the removal of several competitive structures. However, there was no significant trend of the training effects on the leaf area of vegetative short and long shoots. This may be due to the greater sink effect of the fruiting shoot as a whole, compared to vegetative ones (Hansen 1977). Competition between bourse-shoot and fruit growth usually occurs early in the season and is detrimental to fruit-set but both phenomena are positively related later in the season (Abbott 1960; Ferree and Palmer 1982; Quinlan and Preston 1971). Indeed, an increased fruit sink demand stimulates photosynthesis of adjacent bourse and bourse-shoot leaves (Hansen 1977) and, vice versa, an increased leaf area stimulates fruit growth and calcium content (Volz et al. 1994). These positive relationships between fruit and leaf development are well supported by the present study which indicates that leaf area development was similar between fruiting shoots with either one or two fruits—i.e. the two centrifugal training treatments—although individual fruit size was similar in both cases (Lauri et al. 2004).
Fruiting shoot size, length and leaf area are important variables for explaining the return-bloom ability of the laterals (Lespinasse and Delort 1993). Along with other criteria such as bourse volume, it defines the concept of the autonomy of the fruiting shoot, i.e. the ability to develop both a fruit and a bourse-shoot ending in a fruit bud in the same year (Lauri and Lespinasse 1993, 1999). The present study, in agreement with Lauri et al. (2004) results concerning return-bloom, showed that the autonomy of the fruiting shoot was enhanced by training methods, specifically the extinction procedure, for a given cultivar.
The second level of investigation was light interception efficiency, quantified by the STAR. Centrifugal training significantly enhances light interception by the whole-tree canopy as compared to the original solaxe system. However, the effects varied depending on the category the shoots belong to. Vegetative long shoots intercepted more light than other categories, regardless of the treatment, probably because they have larger internode lengths (Takenaka 1994). This should also be related to the spatial distribution of the shoot categories in the canopy. Repeated observations on apple trees suggest that vegetative long shoots were most often in the outer zone of the canopy. This is in accordance with results on walnut trees (Sinoquet et al. 1997). On the other hand, vegetative short shoots and fruiting shoots were more generally found in the inner zone of the canopy. For the latter two shoot categories, the ranking of treatments according to their mean STAR values paralleled the one of mean STAR for the whole tree, with a significant 20% increase of light interception on centrifugal-trained trees as opposed to original solaxe-trained trees. Whereas the STAR of the fruiting shoots on the original solaxe was lower than the STAR of the whole tree, i.e. indicating that fruiting shoots were in shaded areas, centrifugal training maintained a similar STAR, i.e. equivalent light interception capacities, for fruiting shoots as compared to the whole tree. Centrifugal training had a cumulative effect: it not only improved light climate in the whole tree but it also favoured fruiting shoots in comparison with other shoot categories within the trees. This result was due to the lower fruiting shoot density and the central “chimney” in the canopy (Figs. 1, 4a) brought about by extinction in the centre of the tree and on the underside of branches.
High quality fruiting spurs, i.e. with large leaf area and large-sized fruits, are located in zones of the canopy which receive high light levels (Barritt et al. 1991). The objectives of an efficient training system is therefore to maintain a good balance between vegetative and fruit growth by proper tree management, and to improve light interception by the fruiting spurs (Jackson 1980). The positive effects of centrifugal training on fruit size and colour (Ferre et al. 2002; Lauri et al. 2004), return-bloom (Lauri et al. 2004) and fruiting shoot leaf area development (present study) should therefore be interpreted as a direct consequence of the significant enhancement of light interception by the whole tree and more especially by the fruiting shoots.
Attenuation of light intensity, whether direct or diffuse, from the outer zone—at the top and the periphery of the tree crown—to the inner zones of the canopy, is well documented (Génard and Baret 1994; Sinoquet et al. 2001; Tustin et al. 1988). The vertical distribution of STAR values in both original solaxe-trained and centrifugal-trained trees did not follow this scheme. This result therefore supports the idea that the training system, with tree height less than 3 m and with the appropriate low-vigorous rootstock (M9), successfully minimises within-tree shading, decreasing the vertical heterogeneity in light interception by fruiting shoots. The same phenomenon was observed for both the original solaxe-trained and centrifugal-trained trees, but with a tendency towards a more efficient and homogeneous light interception from top to bottom for trees in the latter category.
The study included only two trees per treatment because of (1) the need to digitise trees during a specific phenological window, and (2) time consumption due to digitising, although we used partial digitising (at shoot level) combined with foliage reconstruction. However such a small number of individuals made it possible to identify significant differences between original solaxe and centrifugal training treatments with regard to fruiting shoot leaf area (Table 3) and STAR values at both tree and fruiting shoot scales (Table 4). Ongoing studies aim at refining these results by simulating orchard populations instead of single trees (Génard et al. 2000), and also by comparing various genotypes over several years. This also needs further methodological research to reduce time cost of 3D digitising in the field.
References
Abbott DL (1960) The bourse shoot as a factor in the growth of apple fruits. Ann Appl Biol 48:434–438
Adam B (1999) POL95—software to drive a Polhemus Fastrak 3 SPACE 3D digitiser. Version 1.0. UMR PIAF INRA-UBP, Clermont-Ferrand
Adam B, Sinoquet H, Dones N (2001) VegeSTAR—software to compute light interception and canopy photosynthesis from images of 3D digitised plants. Version 2.0. UMR PIAF INRA-UBP, Clermont-Ferrand
Barritt BH, Rom CR, Konishi BJ, Dilley MA (1991) Light level influences spur quality and canopy development and light interception influence fruit production in apple. HortScience 26:993–999
Corelli-Grappadelli L, Lakso AN, Flore JA (1994) Early season patterns of carbohydrate partitioning in exposed and shaded apple branches. J Am Soc Hortic Sci 119:596–603
Crete X, Lauri PÉ, Ferré G (2002) Pommier—influence de la date d’extinction sur les résultats agronomiques. Réussir Fruits Légumes 206:53–54
Den Dulk JA (1989) The interpretation of remote sensing, a feasability study. PhD thesis. Chapter 3—Turtle, Hare and soil model
Ferre G, Lauri PÉ, Crete X, Tarisse S (2002) Pommier—extinction, branche fruitière et maîtrise de la charge. Réussir Fruits Légumes 206:54–55
Ferree DC, Palmer JW (1982) Effect of spur defoliation and ringing during bloom on fruiting, fruit mineral level, and net photosynthesis of ‘Golden Delicious’ apple. J Am Soc Hortic Sci 107:1182–1186
Génard M, Baret F (1994) Spatial and temporal variation of light inside peach trees. J Am Soc Hortic Sci 119:669–677
Génard M, Baret F, Simon DA (2000) 3D peach canopy model used to evaluate the effect of tree architecture and density on photosynthesis at a range of scales. Ecol Model 128:197–209
Hansen P (1977) Carbohydrate allocation. In: Landsberg JJ, Cutting CV (eds) Environmental effects on crop physiology. Academic, London, pp 247–258
Jackson JE (1980) Light interception and utilization by orchard systems. Hortic Rev 2:208–267
Lakso AN (1980) Aspects of canopy photosynthesis and productivity in the apple tree. Acta Hortic 114:100–109
Lakso AN (1994) Apple. In: Schaffer B, Andersen PC (eds) Handbook of environmental physiology of fruit crops. Temperate crops, vol I. CRC, Boca Raton, pp 3–42
Lakso AN, Corelli-Grappadelli L (1992) Implications of pruning and training practices to carbon partitioning and fruit development in apple. Acta Hortic 322:231–239
Lakso AN, Wünsche JN, Palmer JW, Corelli-Grappadelli L (1999) Measurement and modelling of carbon balance of the apple tree. HortScience 34:1040–1047
Larrive G, Lauri PÉ, Lespinasse JM, Ramonguilhem M (2000) Conduite “centrifuge” avec extinctions. Réussir Fruits Légumes 182:46–48
Lauri PÉ, Kelner JJ (2001) Shoot type demography and dry matter partitioning. A morphometric approach in apple (Malus × domestica Borkh.). Can J Bot 79:1270–1273
Lauri PÉ, Lespinasse JM (1993) The relationship between cultivar fruiting-type and fruiting branch characteristics in Apple trees. Acta Hortic 349:259–263
Lauri PÉ, Lespinasse JM (1999) Apple tree training in France: current concepts and practical implications. Fruits 54:441–449
Lauri PÉ, Terouanne E, Lespinasse JM (1997) Relationship between the early development of apple fruiting branches and the regularity of bearing—an approach to the strategies of various cultivars. J Hortic Sci 72:519–530
Lauri PÉ, Willaume M, Larrive G, Lespinasse JM (2004) The concept of centrifugal training in apple aimed at optimizing the relationship between growth and fruiting. XXVI International Horticultural Congress: Key Processes in the Growth and Cropping of Deciduous Fruit and Nut Trees, Acta Hortic 636
Lespinasse JM (1996) Apple orchard management practices in France. From the vertical axis to the solaxe. Compact Fruit Tree 29:83–88
Lespinasse JM, Delort F (1986) Apple tree management in vertical axis: appraisal after ten years of experiments. Acta Hortic 160:120–155
Lespinasse JM, Delort JF (1993) Regulation of fruiting in apple. Role of the bourse and crowned brindles. Acta Hortic 349:229–246
MAFCOT (1999) Dossier conduite du pommier—Branche fruitière et extinction. Réussir Fruits Légumes 173:27–34
Mierowska A, Keutgen N, Huysamer M, Smith V (2002) Photosynthetic acclimation of apple spur leaves to summer pruning. Sci Hortic 92:9–27
Moon P, Spencer DE (1942) Illumination from a non-uniform sky. Trans Illumin Eng Soc 37:707–712
Palmer JW (1980) Computed effects of spacing on light interception and distribution within hedgerow trees in relation to productivity. Acta Hortic 114:80–88
Palmer JW, Warrington IJ (2000) Underlying principles of successful apple planting systems. Acta Hortic 513:357–363
Pratt C (1990) Apple trees: morphology and anatomy. Hortic Rev 12:265–305
Quinlan JD, Preston AP (1971) The influence of shoot competition on fruit retention and cropping of apple trees. J Hortic Sci 46:525–534
Sansavini S, Corelli-Grappadelli L (1992) Canopy efficiency of apple as affected by microclimatic factors and tree structure. Acta Hortic 322:69–77
Sinoquet H, Bonhomme R (1992) Modeling radiative transfer in mixed and row intercropping systems. Agric For Meteorol 62:219–240
Sinoquet H, Rivet P (1997) Measurement and visualization of the architecture of an adult tree based on a three-dimensional digitising device. Trees 11:265–270
Sinoquet H, Rivet P, Godin C (1997) Assessment of the three-dimensional architecture of walnut trees using digitising. Silva Fenn 31:265–273
Sinoquet H, Thanisawanyangkura S, Mabrouk H, Kasemsap P (1998) Characterization of the light environment in canopies using 3D digitising and image processing. Ann Bot 82:203–212
Sinoquet H, Le Roux X, Adam B, Ameglio T, Daudet FA (2001) RATP, a model for simulating the spatial distribution of radiation absorption, transpiration and photosynthesis within canopies: application to an isolated tree crown. Plant Cell Environ 24:395–406
Stenberg P (1996) Simulations of the effects of shoot structure and orientation on vertical gradients in intercepted light by conifer canopies. Tree Physiol 16:99–108
Takenaka A (1994) Effects of leaf blade narrowness and petiole length on the light capture efficiency of a shoot. Ecol Res 9:109–114
Tarisse S (2001) Modifications expérimentales de la croissance et de la fructification du pommier (Malus × domestica Borkh. cv Galaxy) par la suppression sélective des organes fructifères (extinction artificielle). Diplôme d’ingénieur. Faculté universitaire des sciences agronomiques de Gembloux
Tustin DS, Hirst PM, Warrington IJ (1988) Influence of orientation and position of fruiting laterals on canopy light penetration, yield, and fruit quality of ‘Granny Smith’ apple. J Am Soc Hortic Sci 113:693–699
Tustin DS, Cashmore WM, Bensley RB (1998) The influence of orchard row canopy discontinuity on irradiance and leaf area distribution in apple trees. J Hortic Sci Biotechnol 73:289–297
Volz RK, Ferguson IB, Hewett EB, Wooley EW (1994) Wood age and leaf area influence fruit size and mineral composition of apple fruit. J Hortic Sci 69:385–395
Wagenmakers PS, Nijsse F, De Gendt CME (1991) Planting systems and light climate. Research Station for Fruit Growing, Wilhelminadorp, The Netherlands. Ann Rep 39–40
Wünsche JN, Lakso AN (2000) The relationship between leaf area and light interception by spur and extension shoot leaves and apple orchard productivity. HortScience 35:1202–1206
Wünsche JN, Lakso AN, Robinson TL, Lenz F, Denning SS (1996) The bases of productivity in apple production systems: the role of light interception by different shoot types. J Am Soc Hortic Sci 12:886–893
Acknowledgements
We would like to thank G. Ferré (CEHM), S. Tarisse (student) and J.-C. Salles (INRA) for setting up the experiment and technical support in the orchard, and B. Adam, N. Donès (UMR PIAF, INRA-Université Blaise-Pascal) and J.-J. Kelner for computer and material assistance.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Willaume, M., Lauri, PÉ. & Sinoquet, H. Light interception in apple trees influenced by canopy architecture manipulation. Trees 18, 705–713 (2004). https://doi.org/10.1007/s00468-004-0357-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00468-004-0357-4