
Abstract
Background
Chronic kidney disease (CKD) in children is characterized by severe growth failure. The growth hormone/insulin-like growth factor-1 (GH/IGF-1) axis in uremic animals shows a post-receptor impaired phosphorylation of Janus kinase 2/signal transducer and activator of transcription (JAK-STAT) proteins. The objective of our study was to characterize the intracellular phosphorylation of JAK-STAT signaling in fibroblasts from children with CKD on chronic peritoneal dialysis (PD).
Methods
Serum GH-binding protein (GHBP), IGF-1 and IGFBP3 were measured in 15 prepubertal CKD stage-5 children on PD. Cytoplasmic JAK2, cytoplasmic/nuclear STAT5b and nuclear IGFBP3, acid-labile subunit (ALS) and IGF-1 mRNA expression were quantified in fibroblasts obtained from skin biopsies before and after stimulation with 200 ng/ml recombinant human growth hormone (rhGH). Phosphorylation activity at both the cytoplasmic and nuclear level was expressed as the ratio phosphorylated (p)/total (t) abundance of the product (p/t) at 30 and 60 min. Fifteen healthy children were recruited as the control group. Values were expressed in arbitrary units (AU) and normalized for comparison. Significance was defined as p < 0.05.
Results
Thirty minutes after rhGH stimulus, the cytoplasmic (p/t)JAK2 ratio was significantly lower in patients than in controls [median and interquartile range (IQR): 7.4 (4.56) vs. 20.5 (50.06) AU]. At 60 min after rhGH stimulation, median JAK2 phosphorylation activity was still significantly lower in the patients [7.14 (IQR 3.8) vs. 10.2 (IQR 29.8) AU; p < 0.05]. The increase in the cytoplasmic (p/t)STAT5b/β-actin ratio was lower at both measurement points in the patients compared to the controls, without reaching statistical significance between groups. Median IGFBP3 mRNA abundance was significantly decreased in fibroblasts from uremic patients 24 h after rhGH stimulation compared to the healthy controls [1.27 (IQR 0.83) vs. 2.37 (IQR 0.80) AU]. Median ALS and IGF-1 mRNA expression changed in response to rhGH stimuli at 24 and 48 h.
Conclusion
In this study, children with CKD undergoing PD therapy showed an impaired phosphorylation of JAK2/STAT5b signaling in fibroblasts after GH stimulation, as well as impaired IGFBP3 mRNA abundance. Both impairments may be partially responsible for the observed resistance to the growth-promoting actions of GH in chronic kidney failure.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Growth failure is a major problem in uremic children. According to the 2008 North American Pediatric Renal Trials and Collaborative Studies (NAPRTCS) data [1], almost 40 % of children with chronic kidney disease (CKD) stages II–IV and almost 100 % of CKD stage-V pediatric patients experience growth failure. The 2011 Annual Report of NAPRTCS communicated that, based on the data from more than 2500 patients on peritoneal dialysis (PD), the basal height/age standard deviation score (SDS) of −1.71 drops to −1.76 as early as within the first 12 months of dialysis [2].
In children older than 2 years, growing is mostly dependent on the growth hormone/insulin-like growth factor-1 (GH/IGF-1) axis. Liver-derived IGF-1 is the primary factor that mediates GH’s major effects on postnatal growth [3–8]. In CKD children, multiple abnormalities have been described in this axis, most of which are characterized by a normal or high GH fasting level, prolonged GH half-life due to decreased renal GH clearance and normal or low IGF-1 levels [9, 10], suggesting a GH resistance state in these patients. Various mechanisms have been suggested as possibly contributing to this resistance, including a reduced GH receptor (GHR) density in target organs, an increased production or reduced clearance of IGF-binding proteins (IGFBPs), a defect in the post-receptor GH-activated Janus kinase 2 (JAK2) and signal transducer and activator of transcription (STAT) protein pathways and an oversuppression of GH signaling secondary to the upregulation of the suppressor of cytokine signaling (SOCS) transcription factors SOCS2 and SOCS3 [11–16].
The JAK-STAT intracellular signaling pathway is involved in the binding of GH to its membrane receptor, resulting in the dimerization of GHR, which induces the tyrosine phosphorylation of JAK2, a tyrosine kinase associated with the intracellular domain of the receptor. These tyrosines form binding sites for a number of signaling proteins, including members of the STAT family of signal transducers [17, 18]. To date, seven different STAT isoforms have been identified in humans (STAT1, -2, -3, -4, -5a, -5b, -6), which are expressed in multiple cell types. They respond to several growth factors, including GH, prolactin, interleukins and cytokines, and participate in diverse biological activities [19–23]. STAT proteins, originally identified in interferon signaling pathways, are latent, SH2-domain-containing cytoplasmic factors. Following tyrosyl-phosphorylation, often via a JAK kinase-initiated cascade, cytoplasmic STAT proteins complex with other STAT proteins via an SH2 domain-phosphorylated tyrosine interaction and translocate to the nucleus where they bind to DNA and activate transcription of the target genes.
Downstream GH signals stimulate IGFBP3 nuclear transcription, thereby increasing its serum levels. IGF-1 and IGFBP3 form a ternary complex in the circulation with another protein called acid-labile subunit (ALS), whose gene expression is also regulated by GH [24, 25]. ALS is crucial for maintaining the integrity of the circulating IGF/IGFBP system. It is secreted under the influence of GH, prolongs the half-life of the IGF/IGFBP3 complexes and plays a role in the modulation of IGF-1 availability. In the absence of ALS, serum IGF-1 and in particular serum IGFBP3 activity are markedly reduced [24, 25]. More recently, a novel family of proteins that binds to the cytokine receptor–JAK2 complexes and inhibits JAK2 kinase activity has been identified. These proteins are suppressors of cytokine signaling [SOCS1, -2, -3; cytokine-inducible SH2-containing] and inhibit the GH-induced activation of the GHR-JAK2 complex, thereby acting as an intracellular feedback loop regulating GH sensitivity; they are also induced by GH via the JAK-STAT signaling pathway [14, 26–29].
The current body of knowledge on the JAK-STAT axis in a uremic setting is scarce and only experimental. Schaefer et al. [30] reported that the GH-induced tyrosine phosphorylation of JAK2, STAT5 and STAT3 was 75 % lower in hepatic cells of CKD rats than in those of control animals. These authors also found lower nuclear levels of phosphorylated STAT3 and STAT5 proteins and increased mRNA expression of the suppressor cytokines SOCS2 and SOCS3. These results led the authors to conclude that in this experimental rat model, uremia causes a post-receptor defect in GH signal transduction which is associated to an impaired phosphorylation and nuclear translocation of GH-activated STAT proteins. Even though several research groups have already described an impaired activation of the JAK-STAT axis in various uremic animal models, the characterization of the JAK-STAT axis has not yet been evaluated in uremic humans. The aim of this study was to characterize JAK2/STAT5b pathway in response to GH stimulus in fibroblast cultures from children with CKD receiving chronic PD therapy.
Patients and methods
Patients
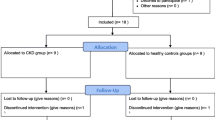
Fifteen prepubertal CKD stage-5 children on PD dialysis were enrolled in the study. All of these children were followed at the Pediatric Nephrology Division, Luis Calvo Mackenna Children’s Hospital, University of Chile. Inclusion criteria were age older than 2 years and Tanner stage 1. Patients on stable PD therapy for at least ≥2 months were selected for entry. Stable PD was defined based on the biochemical parameters of the last 3 months according to the Kidney Disease Outcomes Quality Initiative (K-DOQI) guidelines [31]. Exclusion criteria were Tanner stage 2 or higher, steroid therapy, endocrine diseases, genetic syndromes, gastrointestinal diseases and active nephrotic syndrome.
GH signaling pathway methodology
Cytoplasmic JAK2, cytoplasmic and nuclear STAT5b, nuclear IGFBP3, ALS and IGF-1 mRNA expression were quantified in fibroblasts from skin biopsies before and after stimulation with 200 ng/ml recombinant human GH (rhGH). Phosphorylation activity was expressed as the ratio phosphorylated (p)/total (t) abundance of the product (p/t) at cytoplasmic (normalized by β-actin) and nuclear [normalized by transcription factor IIB (TFIIB)] levels at 30 and 60 min after stimulation. All of these molecules were evaluated in fibroblast cultures obtained from skin biopsy, namely, from a 1-mm2 sample from the non-dominant forearm at the second month of follow-up in patients or from skin of a surgical area in healthy controls. The control group consisted of 15 prepubertal children undergoing elective surgery for phimosis or hernia. Biochemical and histological variables were evaluated at the second month of the protocol to characterize the serum GH/IGF-1 axis.
Clinical evaluation and standard laboratory tests
Anthropometric and nutritional parameters were obtained at the time of enrolment and monthly over a 12-month follow-up period. Height, weight and body mass index (BMI) measurements were expressed as Z-score values. Dialysis adequacy was adjusted according to standard guidelines [31]. The minimal target total dialysis dose (Kt/V) was 2.1. The protein equivalent of urea nitrogen appearance was calculated as previously published [32]. Routine laboratory chemistry values were obtained for all patients using the Vitros® 4600 Chemistry System (Ortho Clinical Diagnostics, Rochester, NY) and included measurements of creatinine, blood urea nitrogen (BUN), total protein, albumin (gradient), hemoglobin, hematocrit, serum ferritin, blood gases, plasma electrolytes, biochemical profile and parathormone levels.
Evaluation of serum GH/IGF-1 axis
Serum GH binding protein (GHBP) was measured at follow-up month 2 using a sandwich enzyme immunoassay technique with detection range of 10–200 ng/ml (Cusabio® ELISA Kit; Cusabio, Wuhan, P.R. China). Serum IGF-1 and IGFBP3 levels were measured concomitantly by immunoradiometric assay (IRMA, IMMULITE 1000) at the central laboratory of the hospital.
Evaluation of intracellular GH/IGF-1 axis
Fibroblast cultures
Fibroblasts were cultured in Dulbecco’s modified Eagle’s medium (DMEM)-red phenol medium containing 10 % simulated body fluid (SBF), 0.1 mg/ml penicillin/streptomycin, 0.23 UI/ml fungisone and 2 mM glutamax. When confluence reached 80 %, the cells were transferred into a trypsin vehicle. When a sufficient number of cells were available, they were frozen in SFB/DMSO at −80 °C for 2 days and then stored in liquid nitrogen until needed for the experiments. In the third cell passage and after 48 h of serum deprivation, cells in DMEM-red phenol were stimulated with 200 ng/ml rhGH (Lot no. AFP8990A; National Institute of Diabetes and Digestive and Kidney Disease, Bethesda, MD). For the protein studies, cytosol and nuclear lysates were obtained at 30 and 60 min after rhGH stimulation to measure cellular response. For the mRNA studies, mRNA of each target protein was isolated at 24 and 48 h after rhGH stimulation for measurement.
Western blot analysis
The nuclear and cytoplasmic fractions were first separated using the NE-PER Nuclear and Cytoplasmic Extraction kit (Thermo Fisher Scientific Inc., Waltham, MA), followed by addition of the protease inhibitor (Complete Mini; Roche Applied Science, Indianapolis, IN) and phosphatase inhibitor cocktails (Phosphatase Inhibitor Cocktails 1 & 2; Sigma, St. Louis, MO). The purity of the nuclear and cytoplasmic fractions was tested using anti-p38α/β (Santa Cruz Biotechnology, Inc., Dallas, TX) and anti-TIFIIB (BD Biosciences, San Jose, CA), respectively. The supernatant was stored (−70 °C) until needed for sodium dodecyl sulfate-polyacrylamide gel electrophoresis and Western blot analysis. Total protein content was measured using the BCA Protein Assay kit (Pierce, Rockford, IL). Proteins were separated on 7.5 or 10 % tris-glycine gels and transferred to nitrocellulose membranes (Invitrogen, Carlsbad, CA). Western blot analysis was performed according to standard conditions. In brief, after blocking non-specific binding, membranes were incubated overnight at 4 °C with rabbit anti-STAT5b (BD Biosciences; the anti-STAT5b antibody does not cross-react with STAT5A), rabbit anti-phospho SAT5b (pY694; BD Biosciences), rabbit anti-JAK2 (Cell Signaling Technology, Danvers, MA), rabbit anti-Phospho-Jak2 (Tyr1007/1008; C80C3; Cell Signaling Technology), rabbit anti-tubulin (Cell Signaling Technology) and anti-TFIIB (BD Biosciences) antibody. After washing with 0.1 % Tween-20 in phosphate buffered saline, blots were incubated with the appropriate horseradish peroxidase-conjugated secondary antibody for 1 h at room temperature. Proteins were detected using an enhanced chemi-luminescence technique (PerkinElmer Life Sciences, Boston, MA). The blots were scanned and densitometry analysis was performed using the public domain NIH Image program v1.61 (US National Institutes of Health: http://rsb.info.nih.gov/nih-image). The results of the measurements of the JAK-STAT molecules were expressed in relative arbitrary units (AU) in order to normalize the basal measure to a value of 1.
Real-time PCR
Total RNA was isolated using TRIzol (GIBCO, Life Sciences Technologies, Thermo Fisher Scientific Inc.) as per the manufacturer’s instructions. RNA concentration was determined by spectrophotometry, and the integrity of the RNA was assessed by agarose gel electrophoresis. cDNA was prepared from total RNA (0.5 μg) using a reverse transcription system (random hexamers; Improm II Reverse Transcriptase System; Promega, Madison, WI). PCR was performed on 8- and 80-ng cDNA samples per 20:1 reaction in triplicate for each experiment (GoTaq Flexi DNA polymerase; Promega). Amplicons were detected real-time fluorescence using the Rotor-Gene Q real-time PCR cycler of Qiagen (Venlo, The Netherlands). This detection system records the number of PCR cycles (Ct) required to produce an amount of product equal to a threshold value, which is a constant. From the Ct values, we calculated the relative mRNA abundance in each experimental condition.
Statistical analysis
Continuous variables were described using the mean and median with the associated standard deviation and interquartile range (IQR), respectively, as variability measures. Basal measurements between patients and controls were compared by the Mann–Whitney test. Responses of the studied variables to rhGH stimulation were compared over time by a two-way analysis of variance for repeated measures using a mixed model. Results were expressed in arbitrary units (AU) normalized by β-actin for cytoplasic measurements and TFIIB for nuclear measurements. Data were processed in statistical package STATA release 12.0 (StataCorp LP, College Station, TX), and significance was defined as p < 0.05.
Results
Clinical and general data
Clinical and anthropometric characteristics of the 15 prepubertal patients (8 boys) and 15 prepubertal controls (14 boys) are shown in Table 1. Mean age of the patients and controls was 5.6 ± 3.02 and 5 ± 1.1 years, respectively. Mean time that patients had been on PD therapy was 10.8 ± 7.8 months. Etiology of CKD was renal dysplasia (n = 8), CAKUT (n = 4), hemolytic uremic syndrome (n = 2) and unknown (n = 1). Height/age Z-scores (SDS) were significantly lower in the patients than in the healthy controls (−2.1 ± 1.7 vs. 0.05 ± 0.06; p < 0.05). The BMI SDS was −0.19 ± 0.83 versus −0.89 ± 2.08 (p = not significant). Bone age in CKD patients was 1.1 ± 1.1 years lower than the chronological age, and growth velocity adjusted for bone age corresponded to the 37 ± 30 percentile (range 1–83 percentile). Biochemical evaluation, creatinine, BUN, total protein, albumin (gradient), hemoglobin, hematocrit, serum ferritin, blood gases, plasma electrolytes, biochemical profile and parathormone values were found to fall within the K-DOQI recommended range (data not shown).
Measurements of serum GH axis
Among the patients, serum IGF-1 and IGFBP3 levels expressed as SDS for chronological age were 1.48 ± 3.16 and 1.34 ± 1.58 SDS, respectively. Following adjustment for bone age, the IGF-1 and IGFBP-3 SDS was 2.7 ± 4.95 and −3.2 ± 2.18, respectively. Median serum GHBP was 21.6 (range 8.7–231) ng/ml and showed a negative correlation to chronological age, bone age, weight, height and BMI and a positive correlation to height velocity (Spearman’s Rho = 0.78, p = 0.002). The relationship between GHBP and residual renal function was found to be non-significant.
Cytoplasmic JAK2 and STAT5b/β-actin phosphorylation activity
Basal cytoplasmic total (t) and phosphorylated (p) JAK2 and STAT5b protein levels [expressed as median (IQR)] in the CKD patients and controls are shown in Table 2. There was a significant difference in the basal cytoplasmic levels of total (t)JAK2 and (t)STAT5b between the patients and controls, being lower in the patients, with (t)JAK2 levels of 0.005 (0.006) versus 0.011 (0.0138) (p = 0.02), respectively, and (t)STAT5b levels of 0.003 (0.002) versus 0.009 (0.012) (p = 0.0002), respectively. Basal levels of (p)JAK2 and (p))STAT5b were also statistically different between the patients and controls, being higher in the patients, with (p)JAK2 levels of 0.09 (0.02) versus 0.004 (0.003) (p = 0.007), respectively, and (p)STAT5b levels of 0.025 (0.005) versus 0.003 (0.005) (p = 0.005), respectively.
Thirty minutes after rhGH stimulus, the (p/t)JAK2/β-actin ratio was significantly lower in patients than in the controls [median (IQR) 7.4 (4.56) vs. 20.5 (50.06) AU, respectively]. At 60 min post-rhGH stimulus, phosphorylation activity was still significantly lower in the patients, with values of 7.14 (3.8) AU in patients and 10.2 (29.8) AU in the controls (p < 0.05) (Fig 1; Table 3). The increase in the cytoplasmic (p/t)STAT5b/β-actin ratio was lower at both time-points in patients [30 min: 6.3 (12.8); 60 min: 5.9 (12.6)] than in controls [30 min:16.6 (27); 60 min: 12.8 (17.9) AU], but the differences between groups were not statistically significant (Fig 2; Table 3). When the values of two outliers from each group were excluded from the analysis, the differences became statistically significant (data not shown).

Cytoplasmic (cyt) phosphorylated/total (p/t) Janus Kinase 2 (JAK2)/β-actin ratios at 0, 30 and 60 min after stimulation with 200 ng/ml recombinant human growth hormone (rhGH) in fibroblast cultures from controls and patients with chronic kidney disease (CKD). The (p/t)JAK2 ratio in both groups was significantly higher than the basal level at 30 and 60 min after stimulation with rhGH (p < 0.05). Patients showed a significantly lower increase in the (p/t)JAK2 ratio than controls at 30 and 60 min (p < 0.05). Box plot Minimum and values, respectively, quantiles 25 %, 50 %, 75 % and maximum value, dots outliers

Cytoplasmic phosphorylated/total (p/t) signal transducer and activator of transcription 5b (STAT5b)/β-actin ratio at 0, 30 and 60 min after stimulation with 200 ng/ml recombinant human growth hormone (rhGH) in fibroblast cultures of controls and patients with chronic kidney disease (CKD). The (p/t)STAT5b ratio was significantly higher in the controls and CKD patients at 30 and 60 min versus the basal levels (p < 0.05). At both 30 and 60 min post-stimulation, the difference in (p/t)JAK2 ratios between the patients and controls was non-significant
Nuclear STAT5b/TFIIB levels
Basal protein abundance of total and phosphorylated STAT5b in the nuclear fraction is shown in Table 2. There were no differences between patients and controls in basal total or phosphorylated STAT5b. After rhGH stimulus, the level of (p/t)STAT5b/TFIIB in the nuclear fraction increased at both time-points in both patients and controls, but the increase was not statistically significant [patients (30 and 60 min): 23.91 (53.7) and 19.5 (39.9) AU; controls: 19.1 (16.7) and 10.1 (24.8), respectively; Fig 3; Table 3].

Nuclear (nuc) phosphorylated/total (p/t) STAT5b/transcription factor IIB (TFIIB) abundance at 0, 30 and 60 min after stimulation with 200 ng/ml recombinant human growth hormone (rhGH) in fibroblast cultures of controls and patients with chronic kidney disease (CKD). The (p/t)STAT5b ratio was significantly higher in the controls and CKD patients at 30 and 60 min versus the basal levels (p < 0.05). At both 30 and 60 min post-stimulation, the difference in (p/t)JAK2 ratios between the patients and controls was non-significant
Figure 4 shows a comparison between cytoplasmic and nuclear (p/t) STAT5b abundance after stimulation with rhGH in both groups. In patients, (p/t) STAT5b abundance was significantly higher in the nuclear fraction than in the cytoplasmic fraction at both 30 and 60 min (p < 0.05) In controls, (p/t) STAT5b abundance was similar at both time-points.

Cytoplasmic versus nuclear (p/t)STAT5b abundance at 30 and 60 min after stimulation with 200 ng/ml recombinant human growth hormone (rhGH) in fibroblast cultures of controls and patients with chronic kidney disease (CKD). In patients, (p/t) STAT5b abundance was significantly higher in the nuclear fraction than in the cytoplasmic fraction at both time-points (p < 0.05) In controls, (p/t) STAT5b abundance was similar at both time-points
Ternary complex mRNA expression of IGFBP3, ALS and IGF-1
Basal IGFBP3, ALS and IGF-1 mRNA expression levels are shown in Table 2. There were no statistical differences between patients and controls. At 24 h following stimulation with rhGH, IGFBP3 mRNA levels had decreased significantly in fibroblasts from uremic patients compared to those of the healthy controls [1.27 (0.83) vs. 2.37 (0.80) AU, respectively; p < 0.05]. At 48 h post-stimulation, the difference between groups was non-significant. There was no significant difference in the expression of ALS mRNA and IGF-1 mRNA in response to rhGH stimuli at 24 and 48 h in the groups (Table 3).
Discussion
The results of our human ex-vivo study confirm that the phosphorylation of the JAK2/STAT5b signaling pathway after rhGH stimulation is impaired in uremic children. The signal transduction cascade induced by GH, acting through its receptor (GHR), triggers an array of multiple biochemical events which involve a large number of distinct molecules. These tyrosines induce phosphorylation of a number of intracellular signaling proteins, including some members of the STAT family, mainly STAT5a and STAT5b. STAT dimers translocate into the nucleus where they bind to specific DNA elements and initiate gene transcription [3, 10, 13–15]. As GHBP is produced by a proteolytic cleavage of the GHR and the extracellular domain is released into the circulation, raised GHBP levels in patients with CKD have been proposed by some authors to represent GH receptor density in target organs [33–35]. In our study, serum GHBP was measured at the time of skin biopsies, when it showed a significant positive correlation to height velocity. The patients in our study showed increased serum GHBP values in comparison to those reported in other studies [36, 37]. As in the case of other binding proteins, this could reflect the accumulation of uremic waste products, but more studies are needed to clarify these observations.
Animal models have been widely used to study GH resistance/insensitivity in CKD. Schaefer et al [30] studied GH signal transduction in hepatic cells of a rat model of chronic renal failure and found a significant decrease of GHR mRNA levels, without any difference in the basal abundance of GHR, JAK2 and STAT1, -3 and -5 protein. These authors reported that cytoplasmic phosphorylation of JAK2 and STAT3 and -5 was significantly impaired following GH stimulus and that the abundance of nuclear (p)STAT5 and -3 was reduced. Also in a rat model, Troib et al [11] described normal GHR and (t)STAT5 basal levels in the serum and growth plate, with decreased levels of (p/t)JAK2, STAT5 and SOCS2. Following the administration of GH, these authors observed a decrease in the phosphorylation of JAK-2 and STAT5b in uremic animals, as well as decreased levels of (p/t)STAT5, normal levels of IGF-1 mRNA and low levels of IGF-1 receptor and serum IGF-1 in CKD rats. Basal levels of cytoplasmic proteins measured in fibroblasts from our patients showed a significant decrease in (t)JAK2 and (t)STAT5b proteins and a significant increase of the phosphorylated form of both proteins. These findings may reflect a compensatory attempt to increase the GH cascade pathway in the uremic state. At the nuclear level we did not observe significant differences between the patient and control groups in terms of basal STAT5b, IGFBP-3, ALS and IGF-1 mRNA expression, results similar to those of Troib [11] and Rabkin [5]. Following the GH stimulus, the phosphorylation activity of JAK2 and STAT5b at the cytoplasmic level was lower in patients than in healthy controls, suggesting that the GH pathway activity in CKD patients is not well preserved. Regarding STAT5b phosphorylation, the cytoplasmic (p/t)STAT5b ratio was lower at both 30 and 60 min following stimulation by GH, but it did not reach any statistical difference between the groups. Of note, when the values of two outliers from each group were excluded from the analysis, the decrease at these two time-points became statistically significant. Although not statistically different, the cytoplasmic (p/t)STAT5b ratio at 30 and 60 min after rhGH injection was three- and twofold lower in uremic fibroblasts than in those from the control biopsy, similar to the results of Schaefer et al. in rats [30]. When analyzing STAT5 activity, isoforms “a” and “b” must be clearly differentiated; however, Schaefer et al. [30] used a STATb antibody which was unable to discriminate between both molecules. All members of the STAT family share a similar structure, with STAT5a and STAT5b being by far the most similar, sharing a 92 % of identity at the amino acid residue level. This similar structure was the reason why STAT5a and STAT5b were considered for many years to be a single entity, with interchangeable biological actions. The identification of different biological patterns associated to mutations in each of these proteins allowed researchers to identify distinct and non-interchangeable functions for each protein [38–40]. In particular, mutations of the STAT5a gene in humans has not yet been associated to any growth retardation. In the present study, we studied only STAT5b and not STAT5a because previous experimental and clinical research indicates that this component of the intracellular GH activation pathway seems to be critical for the expression of both IGF-1 mRNA and IGF-1. The nuclear (p/t)STAT5b ratio in fibroblasts from uremic children was different from that in fibroblasts from the controls, but the difference was statistically non-significant. However, our comparison of cytoplasmic and nuclear (p/t)STAT5b abundance, which reflects translocation activity, revealed a significantly higher nuclear (p/t)STAT5b abundance at the 30 and 60 min post-stimulation time-points compared to that of cytoplasmic (p/t) STAT5b. This observation may reflect the possibility that a higher amount of (p/t) STAT5b was transferred from the cytoplasm to the nucleus in our CKD patients, suggesting some kind of compensation to the low phosphorylation activity observed at the cytoplasmic level. Our results confirm those of previously published studies [15, 20, 22, 23]. Wiezel et al. [15] recently showed in subtotal nephrectomized and pair-fed sham-operated control rats that GHR mRNA and protein levels in the remnant kidney cells, as well as in plasma were reduced, and that basal phosphorylation of JAK2 and STAT5 was significantly impaired. However, in contrast to our observations in our ex-vivo study, these authors reported that GH administration normalized cytoplasmic STAT5 phosphorylated levels, while JAK2 phosphorylation remained impaired.
IGF-1 circulates in the serum as a ternary complex with IGFBP3 and ALS [10, 13, 21, 24, 25]. The major function of ALS is to prolong the half-life of the IGF-1–IGFBP3 binary complex. We measured the expression of ternary complex mRNA after GH stimulation in a subgroup of patients. After 24 h of observation, IGFBP3 mRNA abundance was significantly decreased in fibroblasts from uremic patients compared to those from the healthy controls. This difference was not observed after 48 h of observation. In contrast, ALS mRNA abundance and IGF-1 mRNA expression did not show any differences in response to rhGH stimuli at 24 and 48 h. Schaefer et al. [30] evaluated the effect of rhGH stimuli on IGF-1 and IGFBP1 gene transcription in uremic rats and showed that basal levels of IGF-1 mRNA in the control group did not differ from those of uremic animals. Following GH stimulus, however, IGF-1 mRNA expression in the control rats increased by 79 ± 19 %, while no effect was observed in CKD animals [30].
In this study, we observed that fibroblasts of uremic patients maintained their characteristic uremic activity even after three cell passages, but we believe that it is highly unlikely that human primary cell cultures will fully maintain their uremic character after multiple passages. Every cell in the human body contains the same genetic information, with few exceptions, yet each cell type enacts a distinct gene expression program to allow for highly specialized functions. These tightly controlled programs are the results of transcriptional regulation, by transcription factors and chromatin regulators, as well as by post-transcriptional regulation, mediated in part by microRNAs. Additionally, cells must respond to external cues, and signal transduction pathways converge on the gene regulatory machinery to shape cellular identity and function. Therefore, it is quite possible that uremic cells may acquire some of the features as a result of previous, long-standing uremic conditions. It is also possible that the recovery of the primary human cell cultures have influenced our results, although the differences we documented suggest that the uremic state has—at least in part—an impact on the activation/phosphorylation of the JAK/STAT5 pathways. To clarify these findings, we plan to study the influence of experimental uremic media on human fibroblasts in the near future. We also hope that similar vitro studies will help to clarify the differences in JAK/STAT5 signaling in human, mice and rat fibroblasts under the same conditions as we believe that different species have specific individual responses to GH and uremia.
Conclusion
In summary, in fibroblasts of uremic children on PD, intracellular GH signaling following GH stimuli was associated with a statistically significant depressed response in JAK2 phosphorylation, a 50 % reduction in STAT5b phosphorylation that did not reach statistical significance and decreased IGFBP3 mRNA transcription. Our results suggest that mRNA IGF-1 abundance is well preserved but that its biological activity could be impaired by a decreased IGFBP3 transcription that affects the ternary complex. These results confirm an impaired intracellular GH signaling in CKD children on PD may contribute to growth failure in uremic children. Additional ex-vivo studies are needed to obtain a more detailed insight into the mechanism causing this disturbance to the GH/IGF-1 axis.
References
North American Pediatric Renal Trials and Collaborative Studies (NAPRTCS) (2008) NAPRTCS 2008 Annual Report. Available at: https://web.emmes.com/study/ped/annlrept/annualreport2008.pdf.
North American Pediatric Renal Trials and. Collaborative Studies (NAPRTCS) (2011) NAPRTCS 2011 Annual Report. Available at: https://web.emmes.com/study/ped/annlrept/annualreport2011.pdf
Mahesh S, Kaskel F (2008) Growth hormone axis in chronic kidney disease. Pediatr Nephrol 23:41–48
Wong C, Gipson D, Gillen D, Emerson S, Koepsell T, Sherrard DJ, Watkins S, Stehman-Breen C (2000) Anthropometric measures and risk of death in children with end-stage renal disease. Am J Kidney Dis 36(4):811–819
Rabkin R, Sun D, Chen Y, Tan J, Schaefer F (2005) Growth hormone resistance in uremia, a role for impaired JAK/STAT signaling. Pediatr Nephrol 20(3):313–318
Mahan JD, Warady BS, the Consensus Committee (2006) Assessment and treatment of short stature in pediatric patients with chronic kidney disease: a consensus statement. Pediatr Nephrol 21:917–930
Salevic P, Radovic P, Milic N, Bogdanovic N, Papirovic D, Papirovic A, Golubovic E, Milosevic B, Mulic B, Peco-Antic A (2014) Growth in children with chronic kidney disease: 13 years follow up study. J Nephrol 27(5):537–544
Liu JL, Le Roith D (1999) Insulin-like growth factor I is essential for postnatal growth in response to growth hormone. Endocrinology 140:5178–5184
Roelfsema V, Clark R (2001) The growth hormone and insulin-like growth factor axis: its manipulation for the benefit of growth disorders in renal failure. J Am Soc Nephrol 2001(12):1297–1306
Tonshoff B, Blum WF, Wingen AM, Mehls O (1995) Serum insulin-like growth factors (IGFs) and IGF binding proteins 1, 2, and 3 in children with chronic renal failure: relationship to height and glomerular filtration rate. The european study group for nutritional treatment of chronic renal failure in childhood. J Clin Endocrinol Metab 80:2684–2691
Troib A, Landau D, Kachko L, Rabkin R, Segey Y (2013) Epiphyseal growth plate growth hormone receptor signaling is decreased in chronic kidney disease-related growth retardation. Kidney Int 84(5):940–949
Postel-Vinay C, Tar A, Crosnier H, Broyer M, Rappaport R, Tonshoff B, Mehls O (1991) Plasma growth hormone-binding activity is low in uraemic children. Pediatr Nephrol 5(4):545–547
Houang M, Cabrol S, Perin L, Ducos B, Bensman A, Le Bouc Y (2000) Insulin-like growth factor-I (IGF-I), insulin-like growth factor binding proteins (IGFBP) and insulin-like growth factor type I receptor in children with various status of chronic renal failure. Growth Hormon IGF Res 10(6):332–341
Ocaranza P, Morales F, Román R, Iñiguez G, Cassorla F (2012) Expression of SOCS1, SOCS2, and SOCS3 in growth hormone-stimulated skin fibroblasts from children with idiopathic short stature. J Pediatr Endocrinol Metab 25(3-4):273–278
Wiezel D, Hani Assadi M, Landau D, Troib A, Kachko L, Rabkin R, Segey Y (2014) Impaired renal growth hormone JAK/STAT5 signaling in chronic kidney disease. Nephrol Dial Transplant 29:799–808
Tönshoff B, Edén S, Weiser E, Carlsson B, Robinson IC, Blum WF, Mehls O (1994) Reduced hepatic growth hormone (GH) receptor gene expression and increase in plasma GH binding protein in experimental uremia. Kidney Int 45:1085–1092
Herrington J, Carter-Su C (2001) Signaling pathways activated by the growth hormone receptor. Trends Endocrinol Metab 12(6):252–257
Han Y, Leaman D, Watling D, Rogers N, Groner B, Kerr I, Wood W, Stark G (1996) Participation of JAK and STAT proteins in growth hormone-induced signaling. J Biol Chem 1996(271):5947–5952
Herrington J, Smit LS, Schwartz J, Carter-Su C (2000) The role of STAT proteins in growth hormone signaling. Oncogene 19(21):2585–2597
Davey H, Xie T, Mc Lachlan M, Wilkins R, Waxman D, Grattan D (2001) STAT5b is required for GH-induced liver IGF-I gene expression. Endocrinology 142(9):3836–3841
Denley A, Cosgrove L, Booker G, Wallace J, Forbes B (2005) Molecular interactions of the IGF system. Cytokine Growth Factor Rev 16:421–439
Ram P, Park S, Choi H, Waxman D (1996) Growth hormone activation of Stat 1, Stat 3, and Stat 5 in rat liver. J Biol Chem 271:5929–5940
Davey H, Wilkins R, Waxman D (1999) STAT5 signaling in sexually dimorphic gene expression and growth patterns. Am J Hum Genet 65:959–965
Domené H, Hwa V, Argente J, Wit JM, Camacho-Hubner C, Jasper HG, Pozo J, Van Duyvenvoorde H, Yakar S, Fofanova-Gambetti O, Rosenfeld R, on behalf of the International ALS Collaborative Group (2009) Human acid-labile subunit deficiency: clinical, endocrine and metabolic consequences. Horm Res 72:129–141
Liao J, Hodge C, Meyer D, Sun Ho P, Rosenspire K, Schwaartz J (1997) Growth hormone regulates ternary complex factors and serum response factor associated with the c-fos serum response element. J Biol Chem 272:25951–25958
Starr R, Willson T, Viney E, Murray L, Rayner J, Jenkins B, Gonda T, Alexander W, Metcalf D, Nicola N, Hilton D (1997) A family of cytokine-inducible inhibitors of signalling. Nature 387:917–921
Greenhalgh C, Rico-Bautista E, Lorentzon M, Thaus A, Morgan P, Wilson T, Zervoudakis P, Metcalf P, Street I, Nicola N, Nash A, Fabri L, Norstedt G, Ohlsson C, Flores-Morales A, Alexander W, Hilton D (2005) SOCS2 negatively regulates growth hormone action in vitro and in vivo. J Clin Invest 115:397–406
Flores-Morales A, Greenhalgh C, Norstedt G, Rico-Bautista E (2006) Negative regulation of growth hormone receptor signalling. Mol Endocrinol 20:241–253
Yoshimura A, Naka T, Kubo M (2007) SOCS proteins, cytokine signalling and immune regulation. Nat Rev Immunol 7:454–465
Schaefer F, Chen Y, Tsao T, Nouri P, Rabkin R (2001) Impaired JAK-STAT signal transduction contributes to growth hormone resistance in chronic uremia. J Clin Invest 108:467–475
The National Kidney Foundation Kidney Disease Outcome Quality Initiative (NKF/KDOQI). Available at: https://www.kidney.org/professionals/kdoqi/. Accessed 20 Dec 2014.
KDOQI Workgroup (2009) KDOQI clinical practice guideline for nutrition in children with CKD, 2008 update. Am J Kidney Dis 53[3 Suppl 2]:16-26
Ross RJ, Esposito XY, Shen SA (1997) Short isoform of the human growth hormone receptor functions as a dominant negative inhibitor of the full length receptor and generates large amounts of binding protein. Mol Endocrinol 11:265–273
Tonshoff B, Kiepe D, Ciarmatori S (2005) Growth hormone/insulin-like growth factor system in children with chronic renal failure. Pediatr Nephrol 20:279–289
Leung D, Spencer S, Cachianes G, Hammnonds R, Collins C, Hentzel W (1987) Growth hormone receptor and serum binding protein: Purification, cloning and expression. Nature 330:537–543
Tonshoff B, Cronin M, Reichert M, Haffner D, Wingen A, Blum W, Mehls O (1997) The European study group for nutritional treatment of chronic renal failure in childhood, and members of the German study group for growth hormone treatment in chronic renal failure. Reduced concentration of serum growth hormone (GH)-binding protein in children with chronic renal failure: correlation with GH insensitivity. J Clin Endocrinol Metab 82:1007–1013
Ong K, Elmlinger M, Jones R, Emmett P, Holly J, Ranke B, Dunger B, the ALSPAC Study Team (2007) Growth hormone binding protein levels in children are associated with birth weight, postnatal weight gain, and insulin secretion. Metabolism 56:1412–1417
Rosenfeld R, Belgorosky A, Camacho-Hubner C, Savage M, Wit J, Hwa V (2007) Defects in growth hormone receptor signaling. Trends Endocrinol Metab 18:134–141
Udy GB, Towers RP, Snell RG, Wilkins RJ, Park SH, Ram PA, Waxman DJ, Davey HW (1997) Requirement of STAT5b for sexual dimorphism of body growth rates and liver gene expression. Proc Natl Acad Sci USA 94(14):7239–7244
Hwa V, Nadeau K, Wit J, Rosenfeld R (2011) STAT5b deficiency: lessons from STAT5b gene mutations. Best Pract Res Clin Endocrinol Metab 25:61–75
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
The Ethical Committee, Faculty of Medicine, University of Chile, and the Ethical Committee of the National Research Institution FONDECYT approved this study, and informed consent and assent were obtained from all parents, patients and controls.
Conflict of interest
The authors declare no conflict of interest.
Rights and permissions
About this article

Cite this article
Ugarte, F., Irarrazabal, C., Oh, J. et al. Impaired phosphorylation of JAK2-STAT5b signaling in fibroblasts from uremic children. Pediatr Nephrol 31, 965–974 (2016). https://doi.org/10.1007/s00467-015-3289-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00467-015-3289-x