
Abstract
20% (w/w) Astragali radix was added to the rice medium to cultivate C. kyushuensis Kob. The fermentation product was collected at mycelium stage, coloring stage, stromata-forming initial stage and fruiting body stage of C. kyushuensis Kob. The dynamic content changes of cordycepin and adenosine were detected at different fermentation stages. In the rice medium with Astragalus radix, both cordycepin and adenosine reached the highest content value on the 30th day of fermentation, 17.31 mg/g and 0.94 mg/g, respectively, which were 8.6 times and 2.0 times of that in rice medium at the same stage. At the same time, transcriptomics technology was used to analyze C. kyushuensis Kob during these four periods.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Cordyceps is a type of worm-forming fungus with medicinal value, belonging to Clavicipitaceae, Ascomycotina. Cordyceps contains a variety of active substances, including cordycepin, adenosine, cordyceps polysaccharides, alkaloids, mannitol, and various amino acids. It was commonly used in China for the treatment of fatigue, night sweating, hyperglycemia, hyperlipidemia, asthenia after severe illness, respiratory disease, renal dysfunction, arrhythmias, heart and liver diseases during the past thousands of years. The pharmacological actions of Cordyceps such as antioxidant, immunosuppressive, hypoglycemic, hypolipidemic and antitumor have received significant attention from pharmacological researchers [1,2,3]. Recently, an increasing number of studies have demonstrated the potential of Cordyceps as dietary food and source of medicine in some countries [4,5,6,7].
Cordycepin (3′-deoxyadenosine) is an adenosine analogue, which was first isolated from Cordyceps militaris by Cunningham in 1950 [8]. It has pharmacological properties including anti-oxidation, antitumor, and immune regulation [9, 10]. Compared with adenosine, cordycepin removes the hydroxyl radical at the 3′-C [11]. Some studies have suggested that adenosine is a precursor for cordycepin synthesis [12, 13]. More and more researchers are focusing on the metabolism of C. kyushuensis Kob active substances, hoping to improve the content of cordycepin by improving the cultivation technology, and then enhance the medicinal value of artificial Cordyceps. Mao added millet or soybean to the medium and extended the culture time in dark condition to increase the content of cordycepin [14]. Raethong added different carbon sources to the culture medium to explore the content of cordycepin in Cordyceps militaris, and used transcriptomics to study the metabolic pathway of cordycepin synthesis under different carbon source conditions [15].
Astragalus radix is the dried root of legume Mongolian Astragalus or Astragalus membranaceus, which is an important traditional Chinese medicine [16]. Its main active substances are saponins, flavones, astragalus polysaccharides, which make Astragalus radix have anti-cancer, anti-oxidation, cardiovascular protection and anti-aging effects [17, 18]. Fermentation of traditional Chinese medicine with medicinal fungi can increase the content and efficacy of certain active substances by biotransformation. Lin et al. added Astragalus radix to liquid medium to cultivate Cordyceps militaris, found that the fermentation liquid of Astragalus medium had better antitumor activity than the product cultured in synthetic medium [19]. Bae used fungi such as Cordyceps sisnensis, Cordyceps militaris, Phelllinus linteus and other fungus to ferment Red Ginseng extracts and found that after fermentation by Cordyceps sisnensis and Cordyceps militaris, the content of ginsenosi increased [20].
Transcriptome sequencing technology can quickly and comprehensively detect almost all expressed genes in specific tissues or organs of a particular species, including protein-coding mRNAs and non-coding RNAs (tRNA, rRNA, etc.), which research on gene composition, function, and transcriptional regulation at the RNA level. At present, based on RNA-seq sequencing technology [21], the gene expression of a specific tissue or organ in a specific state can be studied, and then to explore the cell components and biological processes of the tissue or organ. So this technology was also applied to research Cordyceps, especially the exploration of cordycepin synthesis pathway. The whole genome sequencing of Cordyceps militaris, a closely related species with C. kyushuensis Kob, was completed in 2011 [22]. Liu analyzed the transcriptome of different growth stages of Cordyceps cicadae and found that cordycepin synthesis was related to purine nucleotide metabolism [23]. Zhao analyzed the transcriptome, proteome, and mitochondria of C. kyushuensis Kob and found that C. kyushuensis Kob was closely related to Cordyceps militaris, and found that there were four gene clusters related to cordycepin synthesis. They also found that the synthesis of cordycepin was accompanied by the synthesis of pentostatin [24], which is consistent with Cordyceps militaris [25].
In this study, Astragalus radix was added to the rice medium to cultivate C. kyushuensis Kob. The fermentation products were collected at the mycelium stage, coloring stage, stromata-forming initial stage and fruiting body stage of C. kyushuensis Kob, respectively. This work aims to investigate the dynamical changes of active substances during the fermentation process. To reveal the molecular basis of the dynamic changes of active substances, transcriptomics was used to analyze the four periods of C. kyushuensis Kob.
Materials and methods
Microorganism and fermentation
Cordyceps kyushuensis Kob strain was conserved in potato dextrose agar (PDA) media in our lab and C. kyushuensis Kob mycelium was inoculated in PDA liquid medium for activation. Then the active mycelium was inoculated to the medium with 16 g rice and 4 g Astragalus radix in 500 mL glass jars. At the same time, 20 g rice medium without Astragalus radix as a control, then cultured at 23 ℃ for 5–7 days in dark until white mycelium filled the surface of the medium, then at 23 ℃ during the day and 13 ℃ in darkness for 40 days until forming the mature fruiting body. During the entire culture period of C. kyushuensis Kob, four samples were collected for further research, including CKK1 (mycelium stage at the 7th day), CKK2 (coloring stage at the 17th day), CKK3 (stromata-forming initial stage at the 30th day) and CKK4 (fruiting body stage at the 50th day).
Cordycepin and adenosine were purchased from Meilun Biotechnology Co., Ltd. (Dalian, China).
Determination of cordycepin and adenosine by HPLC
Both medium and fungi from four different growth stages were collected, dried to constant weight and then powdered to sift by 60 mesh. 0.5 g powder sample of each growth stage were extracted using tenfold volume purified water to obtain cordycepin and adenosine. All solvents and samples were filtered through 0.45 μm filters before injection into HPLC. HPLC analysis was performed using Agilent 1260 Infinity II LC-system equipped with a TC-C18 column (particle size: 5 μm; length: 250 × 4.6 mm; Agilent, USA). Based on the preliminary experiments [24], the elution conditions for cordycepin and adenosine were performed with the solvents of deionized water (A) and methanol (B, 12%) at a flow rate of 0.8 mL/min for 60 min, which was monitored at a wavelength of 259 nm. The column oven was maintained at a constant temperature of 40 ℃.
RNA isolation and transcriptome sequencing
Total RNAs of C. kyushuensis Kob from different growth stages (CKK1, CKK2, CKK3 and CKK4) were extracted using TRIzol Reagent (Invitrogen)/RNeasy Mini Kit (Qiagen) according to the instructions of the manufacturer. The cDNA libraries were constructed and after quality testing by Agilent 2100 Bioanalyzer, Illumina HiseqTM2500 was used to sequence. Three technical repetitions for each sample and three biological repetitions for each stage were carried out during this experiment.
DEGs analysis
FoldChange and FDR were two criterions to analyze if the same gene has differential expression between two samples. Differential expression genes were selected by p < 0.05 and FoldChange > 2. Afterwards, GO and KEGG analyses were carried out to describe relevant functions.
Results and discussion
The content of cordycepin and adenosine
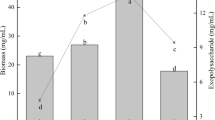
Cordycepin, a natural derivative of adenosine, has been shown to exert pharmacological properties including anti-oxidation, antitumor, and immune regulation. Cordycepin was found in C. kyushuensis Kob, and some literatures speculate that adenosine is the precursor of cordycepin, so we studied the changes of the content of these two substances at four different developmental stages of C. kyushuensis Kob by high-performance liquid chromatography. The content of adenosine and cordycepin in the rice medium increased with the fermentation time (shown in Figs. 1 and 2). On the 50th day of fermentation, the content of cordycepin was 3.32 ± 0.03 mg/g, and the adenosine content was 0.57 ± 0.002 mg/g. While, in the rice medium with Astragalus, both cordycepin and adenosine reached the highest content value on the 30th day of fermentation, 17.31 ± 0.3 mg/g and 0.939 ± 0.002 mg/g, respectively, which were 8.6 times and 2.0 times of that in rice medium at the same stage. Cordycepin and adenosine content increased rapidly from CKK2 (coloring stage at the 17th day) to CKK3 (stromata-forming initial stage at the 30th day) and CKK4 (fruiting body stage at the 45th day). After 30 days of fermentation, the content of these two substances began to decrease. Probably due to Astragalus radix changed the secondary metabolites of C. kyushuensis Kob, therefore, accelerated the degradation of cordycepin and adenosine. In general, the changes in cordycepin and adenosine during the fermentation process were consistent. The possible reason would be that adenosine was thought as the direct precursor of both cordycepin [26] and pentostatin [27]. Xia et al. proved that dual biosynthesis of cordycepin and pentostatin was initiated by a single gene cluster in the medicinal fungus Cordyceps militaris from the precursor adenosine [25].

Cordycepin content at different fermentation time. R rice medium, HQ rice medium with 20% (w/w) Astragalus radix

Adenosine content at different fermentation time. R rice medium; HQ rice medium with 20% (w/w) Astragalus radix
Summary of RNA-Seq data sets
RNA-Seq was performed for RNA samples extracted from four different developmental stages of C. kyushuensis Kob to obtain an overview for further analysis. After filtering out low-quality reads removing the contamination and linker sequences, 57,702,794, 57,412,046, 55,966,926, 56,965,700 clean reads were obtained for the samples of CKK1, CKK2, CKK3 and CKK4, respectively (Table 1). The Q20 of the four samples are more than 95%, Q30 are greater than 90%, and the GC content are about 55%, indicated that the quality of the sequencing data is reliable. The filtered sequencing clean data is compared with the reference genome Cordyceps militaris CM01. The results were shown in Table 2. The contrast ratio of the sequencing sequences that can be mapped to the genome of the four samples is about 90%.
Analysis of differentially expressed genes (DEGs)
Cuffdiff software (v2.2.1) was used to analyze the gene expression of four groups of samples. Pairwise comparison analysis of gene expression levels of four samples through the FPKM distribution map and box diagram of all genes (CKK1-VS-CKK2, CKK1-VS-CKK3, CKK1-VS-CKK4, CKK2-VS-CKK3, CKK2-VS-CKK4, CKK3-VS-CKK4). The test results were screened according to the difference significance standard (different gene expression changed more than two times and p-value ≤ 0.05), and the significant difference expression of genes was counted up and down.
The results are shown in Fig. 3. Among them, the comparison results of CKK1-VS-CKK2 showed 294 down-regulated genes and 284 up-regulated genes; the comparison results of CKK1-VS-CKK3 showed 385 down-regulated genes and 258 up-regulated genes; the comparison results of CKK1-VS-CKK4 showed 191 down-regulated genes and 331 up-regulated genes; and the comparison results of CKK2-VS-CKK3 showed 356 down-regulated genes and 238 up-regulated genes; the comparison results of CKK2-VS-CKK4 showed 214 down-regulated genes and 354 up-regulated genes; the comparison results of CKK3-VS-CKK4 showed that there are 192 down-regulated genes and 432 up-regulated genes.

Differentially expressed genes analysis. Horizontal bar chart shows the number of significant DEGs in each pairwise comparison set
In addition, the Venn diagram was used to analyze the number of common differential genes between each sample and the other three samples. The results are shown in Fig. 4. The number of common differential genes compared CCK1 with CKK2, CKK3 and CKK4 are 103; the number of common differential genes of CKK2 compared with CKK1, CKK3, and CKK4 are 38; the number of common differential genes compared CKK3 with CKK1, CKK2 and CKK4 are 60; the number of common differential genes compared CKK4 with CKK1, CKK2 and CKK3 are 146.

Venn diagram of differentially expressed genes
Functional annotation of differentially expressed genes (DEGs)
GO and KEGG enrichment analysis were performed on the selected DEGs. The final results were screened for the top 30 classification of GO enrichment analysis and the top 20 pathway of KEGG enrichment analysis.
During the growth process of C. kyushuensis Kob, it was necessary to go through the mycelial growth period (CKK1) for 7 days of dark culture. When the white mycelium filed the surface of the culture medium, it could be switched to light culture. During this period, C. kyushuensis Kob mycelium changed from white to yellow, which was the color transfer period (CKK2). GO analysis of the 38 common differential genes of CCK2 compared with CKK1, CKK3 and CKK4 (Fig. 5b) found that the GO terms of the significantly up-regulated genes in CKK2 were “structural constituent of cell wall”, “response to UV”, “oxidoreductase activity” and “nucleotide-excision repair”, etc., suggested that light enabled genes that respond to ultraviolet light expression, which might be related to color conversion. When most of mycelium of C. kyushuensis Kob turned yellow, it entered into the primordium growth stage (CKK3). GO analysis of 60 common differential genes of CCK3 compared with CKK1, CKK2, and CKK4 (Fig. 5c) found that the GO terms of the significantly up-regulated genes in CKK3 were “lipid binding”, “structural constituent of cell wall”, “Aromatic amino acid family metabolic process”, and “endoplasmic reticulum-plasma membrane tethering”, etc., which showed that in CKK3, a large number of substances such as lipids and aromatic amino acids were synthesized, and the intracellular information transmission was increased, which promotes the formation of stromata. GO analysis of 146 common differential genes of CCK4 compared with CKK1, CKK2, and CKK3 (Fig. 5d) found that the GO terms of the significantly up-regulated genes in CKK4 were “nucleic acid binding”, “oxidoreductase activity”, “small-subunit processome”, “ribosome biogenesis”, “carbohydrate metabolic process”, “RNA processing” and “transmembrane transport”. These results indicated that in CKK4, nucleic acid metabolism, ribosome formation and carbon metabolism were accelerated, and cells proliferated to form fruit bodies and spores.

GO enrichment analysis of differentially expressed genes a CKK1; b CKK2; c CKK3; d CKK4
From the GO analysis of the common differential genes in each sample, compared with the other three periods, CKK4 contained the most differential genes. Compared with CKK1, genes significantly up-regulated in CKK4 were more concentrated in “nucleic acid binding”, “carbohydrate metabolic process”, “oxidoreductase activity”, “oxidation–reduction process”, “RNA binding”, “ribosome biogenesis” and “Hydrolase activity, hydrolyzing O-glycosyl compounds” and other entries (Fig. 6a), which indicated that compared with CKK1, there were more genes involved in nucleic acid metabolism, carbon metabolism and redox reactions in CCK4 stage. Compared with CKK2, genes significantly up-regulated in CKK4 were enriched in entries such as “transmembrane transporter activity”, “carbohydrate metabolic process”, “nucleic acid binding” and “transmembrane transport” (Fig. 6b), indicating that there were more genes involved in processes such as nucleic acid metabolism, carbon metabolism and transmembrane transport of substances in CKK4. Compared with CKK3, genes significantly up-regulated in CKK4 were enriched in “transmembrane transport”, “ribosome biogenesis”, “integral component of membrane”, “nucleic acid binding”, “carbohydrate metabolic process” and “hydrolase activity, hydrolyzing O-glycosyl compounds” and other items (Fig. 6c), suggesting there were more genes in CCK4 stage involved in transmembrane transport of substances, nucleic acid metabolism, ribosome generation, carbon metabolism and hydrolysis reactions.

GO analysis of differentially expressed genes a CKK1-VS-CKK4; b CKK2-VS-CKK4; c CKK3-VS-CKK4
KEGG enrichment analysis was performed on the differential genes of CKK1-VS-CKK4, CKK2-VS-CKK4 and CKK3-VS-CKK4. Compared with CKK1, genes significantly up-regulated in CKK4 were enriched in pathways such as “Ribosome biogenesis in eukaryotes”, “Amino sugar and nucleotide sugar metabolism”, “Fatty acid metabolism” and “Sesquiterpenoid and triterpenoid biosynthesis” (Fig. 7a), these pathways are more related to cell growth and proliferation. It is worth noting that compared with CKK1, the “Sesquiterpenoid and triterpenoid biosynthesis” pathway was significantly up-regulated during CCK4, which might be related to cholesterol synthesis. Compared with CKK2, genes significantly up-regulated in CKK4 were concentrated in pathways such as “Ribosome biogenesis in eukaryotes” and “Other glycan degradation” (Fig. 7b). Compared with CKK3, genes significantly up-regulated in CKK4 were concentrated in pathways such as “Ribosome biogenesis in eukaryotes”, “Amino sugar and nucleotide sugar metabolism” and “Other glycan degradation” (Fig. 7c). KEGG enrichment analysis of differential genes specific to CKK4 found that genes significantly up-regulated in CKK4 were enriched in ribosome formation, metabolism of certain amino acids and synthesis or degradation of secondary metabolites (Fig. 7d).

KEGG analysis of differentially expressed genes and KEGG analysis of common Differential genes in CCK4 a CKK1-VS-CKK4; b CKK2-VS-CKK4; c CKK3-VS-CKK4; d CKK4
Uncovering major changes in the transcriptional regulation of metabolism mediated by Astragalus radix
To reveal how the addition of Astragalus radix affects the four growth stages of C. kyushuensis Kob, the differentially expressed genes were subjected to hierarchical clustering analysis to determine the relative expression pattern between the four growth stages of C. kyushuensis Kob grown on Astragalus radix medium (Fig. 8). According to the functional importance of the annotated DEGs, the key genes with regulatory functions were screened (Table 3), such as purine synthesis, carbon metabolism and fatty acid synthesis.

Clustering of differentially expressed genes. Clustering is based on the log10 (FPKM + 1) value. Red indicates high-expressed genes and blue indicates low-expressed genes. The color changes from blue to red, which means that the gene expression is getting higher and higher
During the mycelium growth stage of C. kyushuensis Kob (CKK1), the up-regulated gene was CCM_07964 (hydrophobin). Hydrophobin can help fungi break through the air–liquid barrier to form aerial structures, and at the same time protect the mycelium [28,29,30]. Another gene of concern was CCM_07859 (L-xylulose reductase, [EC:1.1.1.10]), whose product can participate in pentose and glucuronate interconversions, generate xylitol, and then participate in the pentose phosphate pathway to produce ribose 5-phosphate, which is a precursor substance of purines de novo synthesize [31].
Higher content of adenosine and cordycepin were measured during the color changing period of C. kyushuensis Kob, which means the synthesis of purine nucleotides was higher during this period. The genes up-regulated in CKK2 was CCM_05895 (gluconokinase, [EC:2.7.1.12]), which product, glucokinase, participated in the pentose phosphate pathway, thereby promoting the production of ribose 5-phosphate and then providing precursors for purine synthesis. Another up-regulated gene was CCM_08738 (amidophosphoribosyltransferase, [EC:2.4.2.14]). Amidophosphoribosyltransferase catalyzes the synthesis of 5-phosphoribosylamine and then participates in purine metabolism pathway. Purine nucleoside phosphorylase, a product of CCM_04505 (purine nucleoside phosphorylase, [EC:2.4.2.1]), is one of the key enzymes in the purine rescue synthesis pathway. The 5′-nucleotidase producted by CCM_00622 may be involved in the hydrolysis of AMP to generate adenosine. The above enzymes may be involved in the synthesis of cordycepin and adenosine. In addition, no changes in genes related to Astragalus radix saponin synthesis were found. Therefore, after fermentation by C. kyushuensis Kob, the saponin substances in the culture medium were transformed to produce more saponins. But what kinds of enzyme played a role in this process need to be further studied.
At C. kyushuensis Kob fruit body growth stage, cells proliferated in large quantities, and also produced a large number of secondary metabolites. During this stage, the content of adenosine and cordycepin was lower than CKK2. It is speculated that the increase in nucleic acid metabolism may promote more substrate synthesis nucleic acids, and on the other hand, adenosine and cordycepin were degraded. So there was a phenomenon that the adenosine and cordycepin content of CKK4 was lower than CKK2. Most of the genes up-regulated during the CKK4 period were also related to DNA replication, transcription, translation and secondary metabolite synthesis. Among which, CCM_08738 (amidophosphoribosyltransferase, [EC:2.4.2.14]), CCM_06768 (adenylosuccinate synthetase, [EC:6.3.4.4]), which also appeared to be up-regulated during this period, meaning that the products of purine metabolism might be more involved in nucleic acid metabolism to promote cell proliferation. In addition, a large amount of energy was generated during this period, CCM_02997 (fatty acid synthase subunit beta, fungi type, [EC: 2.3.1.86]), which regulated the synthesis of fatty acids. CCM_08316 (hexokinase, [EC: 2.7.1.1]) and CCM_06062 (pyruvate kinase, [EC: 2.7.1.40]) regulated the glycolytic pathway and provided a large amount of precursor material and energy for cell proliferation. It is worth mentioning that CCM_02867 (secreted glucosidase, [EC: 3.2.1]) appeared to be up-regulated during this period. It is speculated that the cells might secrete more glycosidases, which can promote the decomposition of nutrients in the culture medium. In addition, CCM_07184 (squalene monooxygenase, [EC:1.14.14.17]) appeared to be up-regulated in CKK4, which production squalene monooxygenase is one of the important enzymes in cholesterol biosynthesis and also participates in other saponins synthesis [32].
Conclusion
In this study, it was found that by adding 20% (w/w) Astragalus radix to the cultivate medium of C. kyushuensis Kob could improve the content of cordycepin and saponins greatly during the fermentation process. Thus, C. kyushuensis Kob enriched cordycepin and saponins could be cultivated through controlling the cultivate medium and fermentation time. At the same time, the transcriptome technology was used to analyze the gene expression in four growth periods of C. kyushuensis Kob, which to some extent explained the reason for the increase of cordycepin and saponins. In addition, the anti-tumor, anti-aging and cardioprotective effects of the bidirectional fermentation products increased significantly. The relative experiment results will be reported subsequently.
Abbreviations
- CKK:
-
C. kyushuensis Kob
- CKK1:
-
Mycelium stage at the 7th day
- CKK2:
-
Coloring stage at the 17th day
- CKK3:
-
Stromata-forming initial stage at the 30th day
- CKK4:
-
Fruiting body stage at the 50th day
- DEGs:
-
Differentially expressed genes
- HPLC:
-
High performance liquid chromatography
References
Kim HG, Shrestha B, Lim SY (2006) Cordycepin inhibits lipopolysaccharide-induced inflammation by the suppression of NF-kappaB through Akt and p38 inhibition in RAW 264.7 macrophage cells. Eur J Pharmacol 545:192–199
Nakamura K, Yoshikawa N, Yamaguchi Y, Kagota S, Shinozuka K, Kunitomo M (2006) Antitumor effect of cordycepin (3’-deoxyadenosine) on mouse melanoma and lung carcinoma cells involves adenosine A3 receptor stimulation. Anticancer Res 26:43–47
Zhang GY, Yin QS, Han T, Zhao YX, Su JJ, Li MZ, Ling JY (2015) Purification and antioxidant effect of novel fungal polysaccharides from the stroma of Cordyceps kyushuensis. Ind Crop Prod 69:485–491
Cao C, Yang S, Zhou Z (2020) The potential application of Cordyceps in metabolic-related disorders. Phytother Res 34(2):295–305
Liu X, Shang S, Chu W, Ma L, Shao B (2020) Astragaloside IV ameliorates radiation-induced senescence via antioxidative mechanism. J Pharm Pharmacol 72(8):1110–1118
Zhu YL, Yu XF, Qi G, Li J, Ouyang Z (2020) Antioxidant and anti-aging activities of polysaccharides from Cordyceps cicadae. Int J Biol Macromol 157:394–400
Li J, Cai HW, Sun HH (2020) Extracts of Cordyceps sinensis inhibit breast cancer growth through promoting M1 macrophage polarization via NF-κB pathway activation. J Ethnopharmacol 260(5):112969
Cunningham KG, Manson W, Spring FS, Hutchinson SA (1950) Cordycepin, a metabolic product isolated from cultures of Cordyceps militaris (Linn.) Link Nature 166:949–949
Tuli HS, Sharma AK, Sandhu SS, Kashyap D (2013) Cordycepin: A bioactive metabolite with therapeutic potential. Life Sci 93:863–869
Yang R, Wang XL, Xi DS, Mo J, Luo Y (2020) Cordycepin Attenuates IFN-γ-Induced Macrophage IP-10 and Mig Expressions by Inhibiting STAT1 Activity in CFA-Induced Inflammation Mice Model. Inflammation 43(2):752–764
Ling JY, Sun YJ, Zhang HY (2002) Measurement of cordycepin and adenosine in stroma of Cordyceps sp. by capillary zone electrophoresis (CZE). J Biosci Bioeng 94:371–374
Chassy BM, Suhadolnik RJ (1969) Metabolic fate of adenosine and cordycepin by Cordyceps militaris during cordycepin biosynthesis. Biochim Biophys Acta 182(2):307–315
Suparmin A, Kato T, Dohra H, Park EY (2017) Insight into cordycepin biosynthesis of Cordyceps militaris: Comparison between a liquid surface culture and a submerged culture through transcriptomic analysis. PLoS ONE 12:e0187052
Mao XB, Eksriwong T, Chauvatcharin S, Zhong JJ (2005) Optimization of carbon source/nitrogen ratio for cordycepin production by submerged cultivation of medicinal mushroom cordyceps militaris. Process Biochem 40:1667–1672
Nachon R, Kobkul L, Wanwipa V (2018) Uncovering global metabolic response to cordycepin production in Cordyceps militaris through transcriptome and genome-scale network-driven analysis. Sci Rep 8:9250
Wang WT, Zhao ZY, Han YM (2010) Effects of astragaloside IV derivatiue on heart failure in rats. Chin Herb Med 2:48–53
Verotta L, Guerrini M, El-Sebakhy NA, Assad AM, Pezzuto JM (2002) Cycloartane and oleanane saponins from Egyptian Astragalus spp. as modulators of lymphocyte proliferation. Planta Med 68:986–994
Jung Y, Jerng U, Lee S (2016) A systematic review of anticancer effects of Radix Astragali. Chin J Integra Med 22:225–236
Lin YW, Chiang BH (2008) Anti-tumor activity of the fermentation broth of Cordyceps militaris cultured in the medium of Radix astragali. Process Biochem 43:244–250
Bae SH, Lee HS, Kim MR, Kim SY, Kim JM, Suh HJ (2011) Changes of ginsenoside content by mushroom mycelial fermentation in red ginseng extract. J Ginseng Res 35(2):235–242
Wang Z, Mark G, Michael S (2009) RNA-Seq: a revolutionary tool for transcriptomics. Nat Rev Genet 10:57–63
Zheng P, Xia YL, Xiao GH, Xiong C, Hu X, Zhang S (2011) Genome sequence of the insect pathogenic fungus Cordyceps militaris, a valued traditional Chinese medicine. Genome Biol 12:R116
Liu T, Liu Z, Yao X, Huang Y, Qu Q, Shi X, Zhang H, Shi X (2018) Identification of cordycepin biosynthesis-related genes through de novo transcriptome assembly and analysis in Cordyceps cicadae. Roy Soc Open Sci 5(12):181247
Zhao X, Zhang GY, Li CY, Ling JY (2019) Cordycepin and pentostatin biosynthesis gene identified through transcriptome and proteomics analysis of Cordyceps kyushuensis Kob. Microbiol Res 218:12–21
Xia YL, Luo FF, Shang YF, Chen PL, Lu YZ, Wang CS (2017) Fungal Cordycepin biosynthesis is coupled with the production of the safeguard molecule pentostatin. Cell Chem Biol 24:1479–1489
Lennon MB, Suhadolnik RJ (1976) Biosynthesis of 3’-deoxyadeno-sine by Cordyceps militaris: mechanism of reduction. Biochim Biophys Acta 425:532–536
Hanvey JC, Hardman JK, Suhadolnik RJ, Baker DC (1984) Evidence for the conversion of adenosine to 2’-deoxycoformycin by Streptomyces antibioticus. Biochemistry 23:904–907
Asgeirsdottir SA, de Vries OMH, Wessels JGH (1998) Identification of three differentially expressed hydrophobins in Pleurotus ostreatus (oyster mushroom). Microbiology 144(11):2961–2969
Mosbach A, Leroch M, Mendgen KW, Hahn M (2011) Lack of evidence for a role of hydrophobins in conferring surface hydrophobicity to conidia and hyphae of Botrytis cinerea. BMC Microbiol 11(1):1–14
Wosten HAB (2001) Hydrophobins: Multipurpose proteins. Annu Rev Microbiol 55:625–646
Mujchariyakul W, Raethong, (2017) Genome-scale metabolic network of Cordyceps militaris useful for comparative analysis of entomopathogenic fungi. Gene 626:132–139
Chua NK, Scott NA, Brown AJ (2019) Valosin-containing protein mediates the ERAD of squalene monooxygenase and its cholesterol-responsive degron. Biochem J 476(18):2545–2560
Acknowledgements
The research work was supported by National Natural Science Foundation of China (No. 21877075, 21807066). The authors also gratefully acknowledge the financial support of the Key Research and Development Program (No.2019GSF107003) and the Science and Technology Development Program of traditional Chinese medicine (No. 2019-0026) in Shandong Province.
Author information
Authors and Affiliations
Contributions
Junyu Zhang: Fermentation and transcriptome experiment, formal analysis, data curation, methodology, writing—original draft. Tongtong Jian: HPLC experiment and data analysis. Yu Zhang: Fermentation experiment. Guoying Zhang: Writing—review & editing, funding acquisition, supervision. Jianya Ling: Review and editing, funding acquisition, supervision.
Corresponding authors
Ethics declarations
Conflict of interests
On behalf of all authors, the corresponding author states that there is no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Zhang, J., Jian, T., Zhang, Y. et al. Dynamic content changes of cordycepin and adenosine and transcriptome in Cordyceps kyushuensis Kob at different fermentation stages. Bioprocess Biosyst Eng 44, 1793–1803 (2021). https://doi.org/10.1007/s00449-021-02561-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00449-021-02561-3