
Abstract
A new design of cross-linked enzyme aggregates (CLEAs) of Burkholderia cepacia lipase (BCL) based mainly on the use of lignocellulosic residue of palm fiber as an additive was proposed. Different parameters for the preparation of active CLEAs in the hydrolysis of olive oil, such as precipitation agents, crosslinking agent concentration, additives, and coating agents were investigated. The highest activity yield (121.1 ± 0.1%) and volumetric activity (1578.1 ± 2.5 U/mL) were achieved for CLEAs prepared using the combination of a coating step with Triton® X-100 and polyethyleneimine plus the use of palm fiber as an additive. The variations of the secondary structures of BCL-CLEAs were analyzed by second-derivative infrared spectra, mainly indicating a reduction of the α-helix structure, which was responsible for the lipase activation in the supramolecular structure of the CLEAs. Thus, these results provided evidence of an innovative design of BCL-CLEAs as a sustainable and biocompatible opportunity for biotechnology applications.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Enzymatic processes have become a promising alternative to chemical catalysis due to advantages such as high specificity to the substrate type and operation under mild conditions of temperature, pressure and pH with high conversion rates [1]. Nevertheless, compared to the chemical route, the use of enzymatic processes has several limitations for its application at the industrial level, such as the high cost and low stability of enzymes, and when lipases are in the reaction medium, they are usually solubilized and this makes their recovery difficult [2,3,4,5].
Thereby, cross-linked enzyme aggregates (CLEAs) as an immobilization technique have become an alternative to obtain active, insoluble, and low-cost biocatalysts [6,7,8]. This technique is based on the enzyme precipitation (usually polymers, salts or solvents), followed by a cross-linking step with a bifunctional or polyfunctional agent [9]. Normally, the most used cross-linking agent is glutaraldehyde due to its low cost and availability [2]. However, in-depth studies regarding the CLEAs formation are still required, because their use on an industrial scale is difficult due to the low mechanical resistance. On the other hand, the size and porosity of these aggregates can directly affect the catalytic activity of the biocatalyst [1, 10].
To improve the immobilization efficiency and the stabilization of the CLEAs, the use of different additives are investigated, such as amino-rich proteins, ionic liquids, natural and synthetic polymers, surfactants, and magnetic particles [11,12,13,14,15,16,17,18]. Among amino groups-rich proteins, bovine serum albumin (BSA) and soy protein stand out for acting as a protein feeder and to promote increase cross-linking efficiency [6, 12, 14,15,16,17]. In addition, BSA improves the tolerance of the enzyme to the glutaraldehyde (used as cross-linking agent), protecting it against its inactivation [11, 12, 19]. Thus, due to the complex nature involving the enzymatic stabilization, this work proposes for the first time the use of natural fibers as an additive for lipase immobilization by CLEA technique. Palm fiber (from Elaeis guineensis) may result in even a more stable biocatalyst by increasing the mechanical strength of the immobilized material due to the presence of lignin, cellulose, and hemicellulose in the polymeric structures [20]. In addition, it may establish an alternative pathway for bioenergetic sustainability as it is a lignocellulosic residue. Recently, CLEAs of lipase from Rhizopus chinensis was prepared in octyl-modified mesocellular foams using oxidized gum arabic, which exhibited excellent thermal and mechanical stability, and could still maintain 69% of its initial activity after five reaction cycles [21]. Cross-linking of enzymes with nanofibers has shown better residual activity of lysozyme-CLEA using chitosan nanofibers. The immobilized lysozyme-CLEA retained more than 75% of its initial activity [22].
Another strategy in the search for more stable CLEAs is the use of coating agents (such as Triton® X-100, polyethylene glycol, and polyethyleneimine) prior to the protein precipitation step. Coating agents are known to increase the mechanical strength by protecting the enzyme from inactivation and oxidation during storage of immobilized biocatalysts and improving enzyme–substrate interaction [7, 23, 24]. However, the concomitant use of these coating agents has not been evaluated, which could be an interesting strategy to take the better advantages of the individual agents.
Therefore, in this work, a new protocol based on the enzyme surface coating (in absence and presence of additives), precipitation and cross-linking steps for the immobilization of Burkholderia cepacia lipase by CLEA technique was proposed. The three precipitating agents (acetone, ethanol, and ammonium sulfate) and the crosslinking agent (glutaraldehyde) were evaluated considering different concentrations. The best conditions were further evaluated by two different strategies: (1) use of BSA and palm fiber (from Elaeis guineensis) as protein and non-protein feeders, respectively, and (2) application of an enzyme surface coating step with Triton® X-100 and polyethyleneimine prior to the precipitation step. The activity yields of each step were evaluated in an olive oil hydrolysis reaction and the secondary structure of the CLEAs were analyzed using second-derivative infrared spectra by FTIR.
Material and methods
Material
Lipase from Burkholderia cepacia (BCL, ≥ 2.900 U g−1, pH 7.0, 50 °C, and 38.14 mg protein/g), polyethyleneimine (PEI, 50% (w/v) in H2O), and bovine serum albumin (BSA, ≥ 96% of purity) were purchased from Sigma-Aldrich Co (St. Louis, MO, USA). The precipitation agents (acetone, ethanol, and ammonium sulfate, all > 99% of purity) and the gum arabic were purchased from Synth (São Paulo, SP, Brazil). Glutaraldehyde solution (25% in H2O) was purchased from Vetec Quimica (Duque de Caxias, RJ, Brazil), Triton® X-100 (TRX-100, 99.5% of purity) was acquired from Dinâmica (São Paulo, Brazil), the in natura palm fiber (from Elaeis guineensis) was obtained from the Costa do Dendê region (Valença, Bahia, Brazil), and olive oil (Carbonell) was purchased at a local market.
CLEAs synthesis
CLEAs of BCL were prepared following a standard protocol described by Cao et al. [9]. The lipase was first dissolved in sodium phosphate buffer (pH 7.0, 100 mM) at a concentration of 0.3 g/mL and the precipitating agent was added. The resulting mixture of enzyme solution and the precipitating agent was kept for 30 min in an ice bath. Then, an appropriate amount of glutaraldehyde (25% in H2O) was added to the solution and the crosslinking reaction proceeded for 2.5 h. The final suspension was centrifuged at 3400 rpm for 5 min. The precipitate was washed 3 times with sodium phosphate buffer (pH 7.0, 100 mM) and the resulting CLEAs were resuspended in sodium phosphate buffer (pH 7.0, 100 mM).
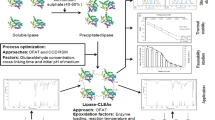
In this work, each step was evaluated separately and the optimum conditions for the precipitating agent, additives, and coating agents were determined. Thus, each step is separately detailed below. Figure 1 shows the simplified procedure for the synthesis of BCL-CLEAs.

Simplified procedure for the synthesis of CLEAS of Burkholderia cepacia lipase
Precipitating agent
Acetone, ethanol and ammonium sulfate were evaluated as precipitating agents. Each precipitant was evaluated separately at the volumetric ratios 1:1, 1:3, 1:5, 1:7, and 1:9 (enzymatic solution:precipitant). Then, the phases were separated, and the solid phase was redissolved in 100 mM sodium phosphate buffer pH 7.0. Tukey's tests were applied for statistical comparisons of the media, with a significance level of 5% (p < 0.05). The precipitation yield in terms of protein content (P) and enzyme activity (EA) was determined by Eq. (1).
where [PAG] and EAAG are the protein concentration and total enzymatic activity of the redissolved aggregates, respectively. [P] is the initial protein concentration (mg/mL), and EAi is the total enzymatic activity of the initial solution of the enzyme.
Cross-linking agent
The cross-linking agent (glutaraldehyde) concentration was evaluated between 25 and 200 mM. After the precipitation step, glutaraldehyde was added, and the crosslinking reaction proceeded for 2.5 h at 4 ± 1 °sC using a water–ice bath to control the temperature. The final suspension was centrifuged at 3400 rpm for 5 min. The solid was washed three times with sodium phosphate buffer (pH 7.0, 100 mM), and the resulting CLEAs were resuspended in sodium phosphate buffer (pH 7.0, 100 mM).
Recovered activity (in %) and total activity yield (in %) of CLEAs (here and after) were determined from Eqs. (2) and (3), respectively.
where EACLEAs is the total enzymatic activity of the CLEAs, EAi is the total activity of the initial solution of enzyme introduced to the immobilization, EAs is the total enzymatic activity in the supernatant at the end of the crosslinking step and EAB is the total enzymatic activity of the wash supernatants of the CLEAs.
Use of additives
BSA and palm fiber were evaluated as additives in the preparation of CLEAs. The additives were added to the enzyme solution (prior to the precipitation step) at the 1:1, 1:3, 3:1, 1:5 and 5:1 mass ratios (enzyme:additive). After, precipitation and crosslinking steps followed as described above.
Coating agents
TRX-100 and PEI were evaluated as coating agents in the preparation of CLEAs. In this case, the BCL solution (0.3 g/mL prepared in 100 mM sodium phosphate buffer pH 7.0, without and with additives—BSA or palm fiber) was treated for 3 h with TRX-100 (1%, w/v) and for 30 min with PEI (1:1, volumetric ratio). The precipitation and crosslinking steps followed as described above.
Protein determination
The total protein concentration was determined by the Lowry method (when organic solvents were used) [25] and direct method (when using neutral salt) [26]. The absorbance of the samples was performed in a UV/Vis spectrophotometer (Varian Cary-50) using a calibration curve constructed with BSA as standard protein. Controls for each sample (without enzyme) were prepared under the same conditions.
Enzymatic activity
The hydrolysis reaction was used to evaluate the enzymatic activity. The method was described by Soares et al. [27] using a substrate formed by (10 mL) olive oil, (10 mL) sodium phosphate buffer (at 100 mM and pH 7.0) and 7% (w/v) gum arabic. The reaction medium containing 5 mL of the substrate and 0.1 mL of enzyme solution (0.3 g/mL) was incubated at 37 °C under constant agitation (80 rpm) for 5 min in a Marconi MA-095 shaker. Then, 2 mL of an acetone/ethanol/water solution (1:1:1, v/v/v) was used to stop the hydrolysis reaction. The liberated fatty acids were titrated with potassium hydroxide solution (0.03 M) using phenolphthalein as an indicator. One unit (U) of enzyme activity was defined as the amount of enzyme that liberated 1 micromol of free fatty acid per min under the assay conditions. For all reactions, three independent experiments were performed. The enzymatic activity (EA) was determined by Eq. (4)
where VTS is the volume of titrated sample (mL), VTB is the volume of titrated blank sample (mL), M is the molarity of KOH (mol L−1) and RT is the reaction time.
FTIR analysis
The secondary structure of the free enzyme and BCL-CLEAs were analyzed by Fourier transform infrared spectroscopy (FTIR) analysis (Agilent Technologies Cary 630 FTIR instrument). The deconvolutions of the Amide I region (1600–1700 cm−1) were performed using the Origin 8.5 software.
Results and discussion
Effect of precipitating agents
The first step of the CLEAs preparation was the selection of the precipitating agent. Precipitation yields (in %) of BCL using different precipitating agents (acetone, ethanol, and ammonium sulfate) in terms of precipitated protein content and enzymatic activity are shown in Figs. 2 and 3, respectively. Figure 2 shows the precipitation yield in terms of precipitated protein content as a function of precipitating agent content used (enzyme solution: precipitating agent, volumetric ratio). In the Fig. 2 is clearly observed that from an enzyme solution:precipitating agent volume ratio of 1:3, the precipitation efficiency remains almost constant for all the evaluated precipitants, with a slightly higher capacity for acetone (88.65 ± 0.04%), followed by ethanol (85.47 ± 0.01%) and ammonium sulfate (84.14 ± 0.01%). The precipitated physical aggregates are formed by non-covalent bonds that can be blocked to the access of large substrates when the cross-linking agent is added. Therefore, it is possible to have an enzymatic conformation that is active upon redissolution with water but inefficient when the aggregate is formed [28].

Precipitation yield (in %) in terms of protein (P) using different volumetric ratios and precipitation agents (acetone, ethanol, and saturated solution of ammonium sulfate). Each point is the average of at least using different volumetric ratios and precipitation agents is and vertical bars indicate standard error of the mean. Differences between means were determined with factorial analyses of variance and Tukey's test for multiple comparisons, All significant a diferences with a significance level of 5% (p < 0.05)

Precipitation yield (in %) in terms of enzyme activity (EA) using different volumetric ratios and precipitation agents (acetone, ethanol, and saturated solution of ammonium sulfate)
Figure 3 shows the precipitation yield as a function of lipase hydrolytic activity. Ammonium sulfate was more efficient to prepare higher active CLEAs of BCL than the other precipitating agents tested (acetone and ethanol), with an activity yield of 290.3 ± 2.9% at an enzyme solution:precipitant ratio of 1:9 (v/v). These results were expected, since organic solvents can denature proteins, mainly due to the disruption of intra-noncovalent interactions [19, 29, 30]. The active enzyme conformation may be restored after redissolving the aggregates in water, but not when they are “freeze” in the CLEAs strucutre. Pan et al. [19] and Hara et al. [12] also obtained similar results regarding the use of ammonium sulfate as precipitating agent for the formation of CLEAs of Serratia marcescens and Burkholderia cepacia lipases, respectively.
Regarding the organic solvents, both showed similar behavior. The highest activity yields were obtained at 1:3 volumetric ratio (214.2 ± 1.4% for acetone and 195.5 ± 2.3% for ethanol), with a decreasing as the amount of solvent was increased, thus reinforcing the deleterious effects on lipase activity. In view of these results, ammonium sulfate at a volumetric ratio of 1:9 (enzyme solution:ammonium sulfate solution) was selected for further studies.
Effect of the cross-linker concentration
The concentration of glutaraldehyde was evaluated in the range of 25–200 mM, using the precipitant and the volumetric ratio of enzyme solution:precipitant previously stablished (Sect. 3.1). The highest total activity yield was 51.6 ± 0.35% at 100 mM glutaraldehyde and with maximal enzymatic activity of 617.5 ± 2.5 U/mL (Table 1). The reduction of the total activity yields for glutaraldehyde concentrations higher than 100 mM may be justified by excessive crosslinking, resulting in loss of enzyme flexibility or diffusion resistance. These results have also been described for CLEAs of Ictularus punctatus and Burkholderia cepacia lipases [12, 31]. Furthermore, at higher concentrations of glutaraldehyde, there is expected to form CLEAs with higher diffusional limitations due to the formation of larger particle size [28]. At low glutaraldehyde concentrations (below 100 mM) the opposite effect is also expected, i.e. insufficient cross-linking yields very flexible and unstable CLEAs allowing enzyme leaching [6, 32].
Effect of additives
The use of non-protein materials, such as chitosan nanofibers and magnetic particles has been successfully described as additives in the CLEAs preparation of different enzymes, aiming to increase the crosslinking efficiency, mechanical resistance of the CLEAs, and also to facilitate their recovery from the reaction medium [8, 13, 18, 22]. Thus, in this work, palm fiber was first evaluated as an additive in the CLEA preparation and the results in terms of total activity yield were compared to those using BSA, a protein additive conventionally used in the CLEA preparation.
Both CLEAs (with BSA and palm fiber) were more active than that prepared in the absence of additives (51.6% total activity yield), achieving total activity yields of 106.7 and 109.1% at an enzyme:additive mass ratio of 1:5, respectively (Fig. 4). Using BSA as co-feeder, the increase in the activity yield may be explained by the high density of amino groups from lysine residues present in the protein surface, which contribute to a more effective crosslinking of the supramolecular structure of the BCL-BSA aggregates [10, 15]. In addition, BSA also contributes to reduce intraparticle diffusional delay [16], mainly when large substrate are used. Thus, it can be supposed that palm fiber also contributed to reduce intraparticle diffusional delays and to increase the crosslinks with glutaraldehyde, because this reagent can react with several functional groups [33]. Thus, the use of palm fiber in the preparation of CLEAs is more attractive than the use of BSA in terms of cost, and also from an economic/environmental point of view, due to the addition of value to an agro-industrial waste.

Total activity yield of CLEAs in the presence of BSA and palm fiber additives using (NH4)2SO4 as precipitation agent (1:9, volumetric ratio) and glutaraldehyde at 100 mM as cross-linking agent
Effect of coating agent
TRX-100 and PEI were used as coating agents prior to the lipase precipitation and crosslinking steps aiming to prepare more active CLEAs. In the case of TRX-100, it has been reported that this non-ionic surfactant changes the lipase conformation in solution to the open form improving its interaction with its substrate [24]. In the case of using PEI to co-aggregate with the enzyme, it can improve the crosslinking efficiency, preventing enzyme leakage and promoting the generation of a hydrophilic microenvironment that protects the enzyme from organic solvents [34,35,36], but it may also contribute to enlarge the pore sizes [18].
CLEAs of BCL prepared in the presence of TRX-100 slightly increased the recovered activity and the total activity yield from 51.6 ± 0.3% (control without coating agent) to 57.4 ± 0.3% (Fig. 5), but almost doubled the volumetric activity of the CLEA (Table 2). This is because the total activity yield was calculated based on the total activity of the initial solution of the enzyme (already hyperactivated with TRX-100) introduced to the CLEA preparation. López-serrano et al. [29] evaluated the preparation of CLEAs of lipases from Candida antarctica (lipase B), Thermomyces lanuginosus, Rhizomucor miehei, Pseudomonas alcaligenes and Candida rugosa using TRX-100. In all cases, the activity yield of the CLEAs based on the hydrolysis of p-nitrophenyl was increased compared to those prepared in the absence of TRX-100.

Recovered activity and total activity yields of the different BCL-CLEAs studied in this work using (NH4)2SO4 as precipitation agent (1:9, mass ratio) and glutaraldehyde at 100 mM as cross-linking agent
Using both TRX-100 and PEI as coating agents, the volumetric activity of the CLEA (1259.9 ± 6.7 U/mL) was 2.0-fold higher than that prepared without any coating agent (BCL control, 617.5 ± 2.5 U/mL), but in this case, the total activity yield was 97.1 ± 0.5% (Table 2). The combined use of TRX-100 and PEI probably could activate the lipase (lid opening due to the surfactant) and form a supramolecular structure more porous and better crosslinked due to the polymer containing a high density of ionized amino groups [37], which is capable to strongly interact with negatively charged functional groups on the protein surface [38], and in certain instances stabilizing the protein [13]. In fact, Yan et al. [39] also prepared firmly cross-linked enzyme aggregates with increased hydrolysis yields from 12 to 42% compared to free lipase by enzyme precipitation with acetone and crosslinking with glutaraldehyde in the presence of PEI.
The combination of coating agents (TRX-100 + PEI) and additives (BSA or palm fiber) was also evaluated in the preparation of BCL CLEAs (Table 2 and Fig. 5). This combination did not greatly increase the total activity yield: from 97.1% (for TRX-100 + PEI) to 113.6 and 121.1% to TRX-100 + PEI + BSA and TRX-100 + PEI + Palm fiber, respectively. But, the presence of additives, particularly palm fiber, can improve other properties of the CLEAs, such as the mechanical strength in stirred tanks and also their recovery from the reaction medium.
Analysis of the BCL secondary structure
Since the formation of CLEAs alters the three-dimensional structure of the protein, the second-derivative infrared spectra (FTIR) in the region between 1700 and 1600 cm−1 was evaluated (Fig. 6). Through FTIR spectra, the secondary structures of the enzyme were observed by amide bands I (1700–1600 cm−1), which present higher sensitivity [40,41,42].

Secondary-derivative spectra of Burkholderia cepacia lipase (BCL) in the amide I region (1700–1600 cm−1) of the: a free lipase; b control CLEAs; c CLEAs with BSA additive; d CLEAs with palm fiber additive; and e CLEAs with palm fiber additive and TRX-100 + PEI. β-sheet is shown in green, random coil in blue, α-helix in purple, and β-turn in orange colours
According to the spectra shown in Fig. 6a, the free enzyme exhibits β-sheet (1638 cm−1), α-helix (1660 cm−1), random coil (1645 cm−1) and β-turn (1670 cm−1). Control CLEAs showed an increase in the α-helix, and changes in the β-sheet, random coil, and β-turn contents (Fig. 6b). These helical structures, responsible for enzymatic activation, were affected by the precipitation and cross-linking steps without the presence of additives or coating agents, leading to the experimental decrease observed in the activity recovery for the control BCL-CLEA. For the CLEAs formed with the use of BSA additive (Fig. 6c), there was no significant reduction of α-helix. However, the β-sheet, random coil, and side chain are less evident compared to the free enzyme, concluding that such structures are bonded together, and thus reducing the absorbance and exposure of the structure. The CLEAs prepared with palm fiber (Fig. 6d) showed a reduction in the β-sheet, side chain and α-helix contents compared to the free enzyme, but the random coil remained more intense. Finally, CLEAs formed from TRX-100, PEI and palm fiber (Fig. 6e) showed the lowest intensity of the α-helix content.
Decreased α-helix content in the formation of the various CLEAs prepared in this work indicates greater lipase activation since the active site of BCL is formed by α-helix. Thus, as the α-helix content is reduced, there is greater exposure of the active site, making the substrate access easier and favouring the increase of enzymatic activity. These results are in agreement with that described by Pan et al. [43] and Barbosa et al. [40] for the increase of BCL enzymatic activity in the presence of ionic liquids. Therefore, these data reinforce the better results for CLEAs prepared with Triton® X-100, PEI and palm fiber, which achieved a total activity yield of 121.1 ± 0.1% and EA of 1578.1 ± 2.5 U/mL.
Conclusion
This work showed the importance of systemically study the preparation of CLEAs, taking in account the influence of the precipitating agent, as well as cross-linking agent concentration, use of additives and coating agents in the preparation of cross-linked aggregates of BCL with high activity. The immobilization of BCL by CLEA technique was more efficient using ammonium sulfate (1:9, volumetric ratio) as precipitation agent, glutaraldehyde as cross-linker at 100 mM concentration, the presence of TRX-100 and PEI as coating agents, and the use of palm fiber as an additive. Under these conditions, CLEAs of BCL were prepared with a total activity yield of approximately 120% (EA of 1578.1 ± 2.5 U/mL). The higher activity of this CLEA was explained by analysis of the lipase secondary structure, showing that the new protocol using palm fiber as additive and TRX-100 and PEI as coating agents may be a promising and more sustainable alternative to the formation of CLEAs.
References
Filho DG, Silva AG, Guidini CZ (2019) Lipases: sources, immobilization methods, and industrial applications. Appl Microbiol Biotechnol 103:7399–7423
Mohamad NR, Marzuki NHC, Buang NA, Huyop F, Wahab RA (2015) An overview of technologies for immobilization of enzymes and surface analysis techniques for immobilized enzymes. Biotechnol Biotechnol Equip 29:205–220
Souza RL, Faria EP, Figueiredo R, Fricks A, Zanin G, Santos OA, Lima A, Soares CF (2014) Use of polyethylene glycol in the process of sol–gel encapsulation of Burkholderia cepacia lipase. J Therm Anal Calorim 117:301–306
Adrio LJ, Demain LA (2014) Microbial enzymes: tools for biotechnological processes. Biomolecules. https://doi.org/10.3390/biom4010117
Souza RL, Faria ELP, Figueiredo RT, Mettedi S, Santos OAA, Lima AS, Soares CMF (2017) Protic ionic liquid applied to enhance the immobilization of lipase in sol–gel matrices. J Therm Anal Calorim 128:833–840
Sheldon AR (2019) CLEAs, Combi-CLEAs and ‘Smart’ magnetic CLEAs: biocatalysis in a bio-based economy. Catalysts. https://doi.org/10.3390/catal9030261
Sheldon RA (2007) Cross-linked enzyme aggregates (CLEA®s): stable and recyclable biocatalysts. Biochem Soc Trans 35:1583–1587
Mehde AA, Mehdi WA, Özacar M, Özacar ZZ (2018) Evaluation of different saccharides and chitin as eco-friendly additive to improve the magnetic cross-linked enzyme aggregates (CLEAs) activities. Int J Biol Macromol 118:2040–2050
Cao L, van Rantwijk F, Sheldon RA (2000) Cross-linked enzyme aggregates: a simple and effective method for the immobilization of penicillin acylase. Org Lett 2:1361–1364
Illanes A, Cauerhff A, Wilson L, Castro GR (2012) Recent trends in biocatalysis engineering. Bioresour Technol 115:48–57
Mahmod SS, Yusof F, Jami MS, Khanahmadi S (2016) Optimizing the preparation conditions and characterization of a stable and recyclable cross-linked enzyme aggregate (CLEA)-protease. Bioresour Bioprocess 3:3
Hara P, Hanefeld U, Kanerva LT (2008) Sol–gels and cross-linked aggregates of lipase PS from Burkholderia cepacia and their application in dry organic solvents. J Mol Catal B Enzym 50:80–86
Guimarães JR, Giordano RdLC, Fernandez-Lafuente R, Tardioli PW (2018) Evaluation of strategies to produce highly porous cross-linked aggregates of porcine pancreas lipase with magnetic properties. Molecules. https://doi.org/10.3390/molecules23112993
Mafra CA, Ulrich GL, Kornecki FJ, Fernandez-Lafuente R, Tardioli WP, Ribeiro PM (2019) Combi-CLEAs of glucose oxidase and catalase for conversion of glucose to gluconic acid eliminating the hydrogen peroxide to maintain enzyme activity in a bubble column reactor. Catalysts. https://doi.org/10.3390/catal9080657
Ramos MD, Miranda LP, Giordano RLC, Fernandez-Lafuente R, Kopp W, Tardioli PW (2018) 1,3-Regiospecific ethanolysis of soybean oil catalyzed by crosslinked porcine pancreas lipase aggregates. Biotechnol Prog 34:910–920
Mafra ACO, Kopp W, Beltrame MB, de Giordano RLC, de Ribeiro MPA, Tardioli PW (2016) Diffusion effects of bovine serum albumin on cross-linked aggregates of catalase. J Mol Catal B Enzym 133:107–116
Rojas JM, Amaral-Fonseca M, Zanin MG, Fernandez-Lafuente R, Giordano DR, Tardioli WP (2019) Preparation of crosslinked enzyme aggregates of a thermostable cyclodextrin glucosyltransferase from Thermoanaerobacter sp. critical effect of the crosslinking agent. Catalysts. https://doi.org/10.3390/catal9020120
Amaral-Fonseca M, Kopp W, Giordano DR, Fernández-Lafuente R, Tardioli WP (2018) Preparation of magnetic cross-linked amyloglucosidase aggregates: solving some activity problems. Catalysts. https://doi.org/10.3390/catal8110496
Pan J, Kong X-D, Li C-X, Ye Q, Xu J-H, Imanaka T (2011) Crosslinking of enzyme coaggregate with polyethyleneimine: a simple and promising method for preparing stable biocatalyst of Serratia marcescens lipase. J Mol Catal B Enzym 68:256–261
Datta S, Christena LR, Rajaram YRS (2013) Enzyme immobilization: an overview on techniques and support materials. 3Biotech 3:1–9
Jin W, Xu Y, Yu X-W (2019) Preparation of lipase cross-linked enzyme aggregates in octyl-modified mesocellular foams. Int J Biol Macromol 130:342–347
Park J-M, Kim M, Park H-S, Jang A, Min J, Kim Y-H (2013) Immobilization of lysozyme-CLEA onto electrospun chitosan nanofiber for effective antibacterial applications. Int J Biol Macromol 54:37–43
López-Gallego F, Betancor L, Hidalgo A, Alonso N, Fernández-Lafuente R, Guisán JM (2005) Co-aggregation of enzymes and polyethyleneimine: a simple method to prepare stable and immobilized derivatives of glutaryl acylase. Biomacromology 6:1839–1842
Perna RF, Tiosso PC, Sgobi LM, Vieira AMS, Vieira MF, Tardioli PW, Soares CMF, Zanin GM (2017) Effects of Triton X-100 and PEG on the catalytic properties and thermal stability of lipase from Candida rugosa free and immobilized on glyoxyl-agarose. Open Biochem J 11:66–76
Lowry OH, Rosebrough NJ, Farr AL, Randall RJ (1951) protein measurement with the folin phenol reagent. J Biol Chem 193:265–275
Stoscheck CM (1990) Quantitation of protein. In: Deutscher MP (ed) Methods in enzymology. Academic Press, New York
Soares CMF, De Castro HF, De Moraes FF, Zanin GM (1999) Characterization and utilization of Candida rugosa lipase immobilized on controlled pore silica. Appl Biochem Biotechnol 77–9:745–757
Diaz-Vidal T, Armenta-Perez VP, Rosales-Rivera LC, Mateos-Díaz JC, Rodríguez JA (2019) Cross-linked enzyme aggregates of recombinant Candida antarctica lipase B for the efficient synthesis of olvanil, a nonpungent capsaicin analogue. Biotechnol Prog 35:e2807
López-Serrano P, Cao L, van Rantwijk F, Sheldon RA (2002) Cross-linked enzyme aggregates with enhanced activity: application to lipases. Biotechnol Lett 24:1379–1383
Asakura T, Adachi K, Schwartz E (1978) Stabilizing effect of various organic solvents on protein. J Biol 253:6423–6425
Senan Mahmod S, Yusof F, Jami M (2014) Production of cross-linked enzyme aggregate-lipase from channel catfish (Ictularus punctatus) Viscera. In: International conference on biological, chemical and environmental sciences. https://doi.org/10.15242/IICBE.C614017
Sheldon RA (2011) Characteristic features and biotechnological applications of cross-linked enzyme aggregates (CLEAs). Appl Microbiol Biotechnol 92:467–477
Migneault I, Dartiguenave C, Bertrand MJ, Waldron KC (2004) Glutaraldehyde: behavior in aqueous solution, reaction with proteins, and application to enzyme crosslinking. Biotechniques 37:790–802
Zheng J, Chen Y, Yang L, Li M, Zhang J (2014) Preparation of cross-linked enzyme aggregates of trehalose synthase via co-aggregation with polyethyleneimine. Appl Biochem Biotechnol 174:2067–2078
Virgen-Ortíz JJ, dos Santos JCS, Berenguer-Murcia Á, Barbosa O, Rodrigues RC, Fernandez-Lafuente R (2017) Polyethylenimine: a very useful ionic polymer in the design of immobilized enzyme biocatalysts. J Mater Chem B 5:7461–7490
Mateo C, Abian O, Fernandez-Lafuente R, Guisan JM (2000) Reversible enzyme immobilization via a very strong and nondistorting ionic adsorption on support–polyethylenimine composites. Biotechnol Bioeng 68:98–105
Bolivar JM, Rocha-Martin J, Mateo C, Cava F, Berenguer J, Fernandez-Lafuente R, Guisan JM (2009) Coating of soluble and immobilized enzymes with ionic polymers: full stabilization of the quaternary structure of multimeric enzymes. Biomacromology 10:742–747
Wilson L, Betancor L, Fernández-Lorente G, Fuentes M, Hidalgo A, Guisán JM, Pessela BCC, Fernández-Lafuente R (2004) Cross-linked aggregates of multimeric enzymes: a simple and efficient methodology to stabilize their quaternary structure. Biomacromology 5:814–817
Yan J, Gui X, Wang G, Yan Y (2012) Improving stability and activity of cross-linked enzyme aggregates based on polyethylenimine in hydrolysis of fish oil for enrichment of polyunsaturated fatty acids. Appl Biochem Biotechnol 166:925–932
Barbosa MS, Freire CCC, Souza RL, Cabrera-Padilla RY, Pereira MM, Freire MG, Lima ÁS, Soares CMF (2019) Effects of phosphonium-based ionic liquids on the lipase activity evaluated by experimental results and molecular docking. Biotechnol Prog 35:e2816
Madeira Lau R, Sorgedrager MJ, Carrea G, van Rantwijk F, Secundo F, Sheldon RA (2004) Dissolution of Candida antarctica lipase B in ionic liquids: effects on structure and activity. Green Chem 6:483–487
Wi S, Pancoska P, Keiderling TA (1998) Predictions of protein secondary structures using factor analysis on Fourier transform infrared spectra: Effect of Fourier self-deconvolution of the amide I and amide II bands. Biospectroscopy 4:93–106
Pan S, Liu X, Xie Y, Yi Y, Li C, Yan Y, Liu Y (2010) Esterification activity and conformation studies of Burkholderia cepacia lipase in conventional organic solvents, ionic liquids and their co-solvent mixture media. Biores Technol 101:9822–9824
Acknowledgements
The authors thank the financial support of FAPITEC/SE, Coordenação de Aperfeiçoamento de Pessoal de Nível Superior—Brasil (CAPES)—Finance Code 001; Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq); and Universidade Tiradentes (PROSUP II—UNIT) for the scholarship to Alves, N.R. The authors also thank São Paulo Research Foundation (grant 2016/10636-8) for some kind of financial help.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Alves, N.R., Pereira, M.M., Giordano, R.L.C. et al. Design for preparation of more active cross-linked enzyme aggregates of Burkholderia cepacia lipase using palm fiber residue. Bioprocess Biosyst Eng 44, 57–66 (2021). https://doi.org/10.1007/s00449-020-02419-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00449-020-02419-0