
Abstract
In this study, the microalga Scenedesmus dimorphus was cultivated phototrophically using unsterilized anaerobic digestate as a nutrient medium. A bench-scale experiment was conducted by inoculating the microalga S. dimorphus with 0.05–10% dilutions of the anaerobic digestate supernatant. It was found that 1.25–2.5% dilutions, which is equivalent to 50–100 mg N/L total nitrogen concentrations and 6–12 mg P/L total phosphorus concentrations, provided sufficient nutrients to maximize the growth rate along with achieving high concentrations of algal biomass. The microalgae cultivation was scaled up to 100 L open raceway ponds, where the effect of paddlewheel mixing on the growth was investigated. It was concluded that 0.3 m/s water surface velocity yielded the highest specific growth rate and biomass concentration compared to 0.1 and 0.2 m/s. The microalga S. dimorphus was then cultivated in the raceway ponds using 2.5% diluted anaerobic digestate at 317 and 454 μmol/(m2 × s) average incident light intensities and 1.25% diluted anaerobic digestate at 234 and 384 μmol/(m2 × s) average incident light intensities. The maximum biomass concentration was 446 mg/L which was achieved in the 2.5% dilution and 454 μmol/(m2 × s) light intensity culture. Moreover, nitrogen, phosphorus, and COD removal efficiencies from the nutrient media were 65–72, 63–100, and 78–82%, respectively, whereas ammonia was completely removed from all cultures. For a successful and effective cultivation in open raceway ponds, light intensity has to be increased considerably to overcome the attenuation caused by the algal biomass as well as the suspended solids from the digestate supernatant.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
In the past, algal cultivation was limited to laboratory scale experiments which aimed at food and feed purposes. After 1948, outdoor cultivation began by utilizing flue gases from industries to provide algae with carbon dioxide and the harvested biomass was used for protein synthesis. Later, outdoor algal cultivation systems were employed for wastewater treatment through a combination of microalgae and bacteria. In 1979, the production of biofuels from aquatic plants began by the Solar Energy Research Institute (SERI) for the United States Department of Energy [1].
Large-scale cultivation of microalgae for biofuel and other purposes can be achieved in open or closed photobioreactors. While closed photobioreactors offer the advantage of easier contamination control, they are more expensive to construct and operate. On the other hand, open systems are widely used for the cultivation of microalgae due to the ease of construction and scale-up [2, 3]. Among the open cultivation systems, raceway ponds are the most attractive choice for algae cultivation. A raceway pond is a closed loop that is built with concrete, compacted earth material, or plastic. The typical operating depth is 0.1–0.3 m and the biomass and nutrients are recirculated within the pond with the aid of a paddlewheel. Nutrient medium is often introduced ahead of the paddlewheel, while harvesting takes place behind it. The introduction of carbon dioxide to the culture via bubbling is optional; as it enhances the growth of microalgae [4]. One of the biggest challenges facing the cultivation of microalgae in raceway ponds is the photosynthesis inefficiency due to light attenuation in the water column, especially in deep cultures [2]. Practices such as flashing lights and forced light/dark cycles have been suggested to improve the photosynthetic efficiency and biomass productivity [2, 5]. Nevertheless, these practices might be impractical or hard to implement for outdoor cultures.
Microalgae cultivation for wastewater treatment has been studied broadly. Species such as Chlamydomonas, Botryococcus, Chlorella, Haematococcus, Spirulina, and Scenedesmus have been utilized for wastewater treatment [6]. For instance, Tam and Wong [7] revealed that sewage samples from preliminary and primary sedimentation as well as activated sludge secondary effluent supported the growth of Chlorella pyrenoidosa. Anaerobic digestion, which is a widespread approach used to manage organic solid wastes such as animal manure, produces nutrient-rich slurry (digestate) besides biogas. This nutrient-rich liquid waste can be used to cultivate microalgae; however, there are some challenges in utilizing the anaerobic digestate (AD) to cultivate microalgae such as high nutrients concentrations and suspended solids. Furthermore, growth of microalgae can be inhibited by high concentrations of constituents such as ammonia and chemical oxygen demand (COD) [8]. Several studies have indicated the potential of cultivating microalgae using anaerobic digestion effluent or slurry. For example, the microalga N. oleoabundans has been cultivated using the digestate and the effluent from digesting animal manures as well as other organic wastes [8]. Park et al. [9] reported that the microalga Scenedesmus accuminatus was cultivated using filtered and autoclaved anaerobic digestion effluent from a local piggery farm. However, they observed inhibition to the growth at 200 mg NH4-N/L or higher concentrations. Tam and Wong [10] revealed that ammonia concentrations up to 250 mg NH4-N/L did not inhibit the growth of Chlorella vulgaris. However, they indicated that ammonia inhibition is a function of pH, as nitrogen exists in the non-toxic ammonium form at pH values less than 8. Travieso et al. [11] stated that the growth of the microalga C. vulgaris was not inhibited when cultivated using piggery wastewater at COD concentrations less than 1100 mg/L. They speculated that the inhibition may be caused by high suspended solids resulting in light attenuation in the water column. It is noteworthy to mention that many studies conducted on the cultivation of microalgae using wastewaters reported using sterilized instead of raw wastewaters to prevent interference from other microorganisms. Common sterilizing techniques are filtration and autoclaving [9].
Scenedesmus dimorphus is a freshwater microalga from the Chlorophyceae class that is known for its high growth rates and total fatty acids content [12]. This microalga can grow in four distinguishable forms depending on the growth conditions such as temperature, radiation, and nutrients availability. The typical form is a colony of four cells with crescent-shaped outer cells. Other forms include single cells, rounded four-cell colonies resembling the shape of S. obliquus, and rounded single cells [13].
Several studies have been conducted on the microalga S. dimorphus and its applicability for biofuels production. Besides biodiesel synthesis, anaerobic digestion of the S. dimorphus biomass has been suggested as an effective way to produce bio-methane due to the high methane yield of this species which can reach up to 400 mL methane/g VS [14]. Moreover, this microalga is a potential candidate for carbon dioxide sequestration produced from fossil fuels burning as well as flue gases due to its high tolerance to elevated concentrations of carbon dioxide, a range of pH values, and salinity concentrations [15]. It has been indicated that S. dimorphus can tolerate up to 20% CO2, 500 ppm NO, and up to 100 ppm SO2 concentrations when fed with flue gas. These conditions might impose toxic effects for many other algal species [16]. Giannetto et al. [17] demonstrated that the tolerance of this species to elevated carbon dioxide concentrations can work as a tool to mitigate the contamination risk by other invasive species; thus, maintaining the abundance of S. dimorphus in the culture. Additionally, S. dimorphus is known for its ability to remove ammonia and phosphorus from a variety of wastewaters [18]. In fact, Kang and Wen [19] revealed that S. dimorphus was able to remove gaseous ammonia when blended with carbon dioxide and air. High biomass concentrations in the 2.5–6.5 g/L range were reported in the literature for this microalga [16, 20, 21]. In addition to its remarkable growth rates and high biomass concentrations, this species can accumulate high lipid content which can range from 18 to 32% by weight [22, 23]. The most abundant fatty acids in this microalga are palimitic acid, alpha-linolenic acid, cis-9-oleic acid, and cis-9,12-linoleic acid [15, 20].
One of the major obstacles facing the large scale cultivation of microalgae is the expensive production cost as a result of nutrients and energy requirements [8, 24]. One way to offset the production costs of microalgae biomass is by utilizing readily available nutrient media including a variety of wastewaters [8]. Although studied widely, the large-scale industrial application of microalgae in wastewater treatment is still limited [25]. Many studies have focused on cultivating S. dimorphus for nutrients removal from wastewaters; however, very few studies focused on the controlled cultivation of this microalga in open raceway ponds using wastewater as a nutrient medium. In fact, to our best knowledge, this is the first study to discuss the cultivation of S. dimorphus in an open raceway pond using diluted AD.
In this study, the microalga S. dimorphus was cultivated in a sustainable manner using diluted AD. The nutrient medium was not sterilized or filtered in our experiment as opposed to most of the studies on this topic. A bench-scale experiment was conducted by inoculating the microalga S. dimorphus with diluted AD supernatant at a range of dilutions to determine the optimum dilution that will provide sufficient nutrients to support the growth of this microalga. The microalgae cultivation was then scaled up to 100 L raceway ponds. The microalga S. dimorphus was cultivated using two dilutions of the AD supernatant and each dilution was tested under two light intensities. The growth of the microalgae as well as the nutrients removal efficiencies was evaluated by frequent monitoring.
Materials and methods
Anaerobic digestate
AD was sampled from a commercial digester in Columbus, OH, that typically accepts food waste and animal manure for anaerobic digestion. The unsterilized digestate was stored in the laboratory at 4 °C until the time of analysis.
Analytical methods
Total nitrogen, total phosphorus, ammonia, and COD concentrations were determined according to the colorimetric methods in compliance with APHA Standard Methods for the Examination of Water and Wastewater and EPA methods (HACH methods 10072, 10127, and 10031, and 8000) using HACH DR 3900 spectrophotometer. The algal biomass concentration was quantified by measuring the optical densities at 680 and 750 nm wavelengths (OD 680 and OD 750). A correlation was established between the S. dimorphus biomass concentration as total suspended solids (TSS) and the OD 680. However, this correlation was not valid when this microalga was grown with turbid media such as the AD supernatant. It was observed during the experimental work that the ratio between OD 680 and OD 750 was almost constant for the microalga S. dimorphus, but this ratio tends to decrease as the microalgae solution was mixed with AD supernatant. Thus, a correlation between the OD 680/OD 750 ratio and the actual portion of microalgae biomass concentration in the mixture of microalgae and AD supernatant was established. It was indicated by de Winter et al. [26] that the OD 680/OD 750 ratio is a relative measure of pigment/chlorophyll content of the algal cells. For this purpose, three supernatant dilutions were prepared (TSS concentrations: 40; 80, and 142 mg/L) and mixed with S. dimorphus solution (biomass concentration: 330 mg/L) at four mixing ratios (20, 40, 60, and 80% microalgae by volume); thus, a total of 12 solutions were tested. The OD 680 and OD 750 were measured for each separate component of the mixture (microalgae or AD supernatant) as well as the 12 combinations of microalgae and AD supernatant. In addition, the actual TSS concentrations of algae and supernatant were predetermined prior to mixing. The correlation was established between the OD 680/OD 750 ratio and the fraction of the actual microalgae biomass concentration with respect to the inaccurate biomass concentration determined by the OD 680—TSS correlation. Hence, the biomass concentrations estimated using OD 680—TSS correlation were corrected to account for the actual algal portion in the mixture of microalgae and AD supernatant. Measurement of TSS was performed according to the APHA Standard Methods for the Examination of Water and Wastewater (method 2540 D) [27].
Microalgae selection
The microalga S. dimorphus, which was previously obtained from Algaeventure Systems (Marysville, OH, USA) and cultivated in our laboratory, was selected for cultivation in the bench-scale experiments as well as in the raceway ponds. The microalgae were cultivated in BG-11 medium [28] and kept in an incubator at 25 °C with 14:10 light/dark cycle using two built-in fluorescent lamps with an average light intensity of 50 μmol/(m2 × s).
Bench-scale microalgae cultivation
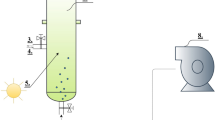
The microalga S. dimorphus was cultivated on a bench scale to determine the optimum dilution or range of dilutions which will provide sufficient nutrients to optimize the growth of this microalga on a larger scale. The nutrient medium used was the diluted and unsterilized AD supernatant which was prepared in a similar manner as explained in Abu Hajar et al. [8]. A range of 0.05–10% dilutions were tested that is equivalent to 2–400 mg N/L and 0.23–48 mg P/L. Each dilution was tested in five replicates by mixing 4 mL of nutrient medium with 2 mL microalgae (initial OD 680 = 0.125) in 16 mm round glass vials (Fig. 1a). The vials were kept inside an incubator at 25 °C with an average light intensity of 50 μmol/(m2 × s) and 14:10 light–dark cycle. The biomass concentration was measured by placing each vial inside the spectrophotometer, and OD 680 and OD 750 were recorded for each vial. Specific growth rates were calculated for each dilution using the least-squares regression method, and statistical analysis was performed using IBM SPSS software to compare the algal growth under different dilutions.

Microalgae cultivation: a bench-scale experiment, b raceway ponds
Microalgae cultivation in the raceway ponds
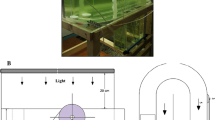
The microalga S. dimorphus was cultivated in 100 L raceway ponds which were purchased from MicroBio Engineering (http://www.microbioengineering.com). The length and width of the pond were 95 and 40 cm, respectively, and the radius of the bend was 20 cm. The water depth equivalent to 100 L was 20 cm. Temperature and pH of the microalgae cultures in each pond were monitored using Neptune APEX controller (http://www.neptunesystems.com), while pH was regulated by carbon dioxide bubbling to maintain a pH value of 7.5 ± 0.1. Light was supplied by a 760 W UFO LED Grow Light providing red, blue, orange, and white colors (Fig. 1b). Temperature was maintained at 25 ± 1 °C using Eheim Jager 200 W aquarium heater. To examine the effect of paddlewheel mixing, the microalga S. dimorphus was cultivated in the raceway ponds at three different water surface velocities: 0.1, 0.2, and 0.3 m/s using BG-11 as a nutrient medium at an average incident water surface light intensity of 454 μmol/(m2 × s) and 14:10 light–dark cycle. Then, the microalgae were cultivated using AD supernatant as nutrient media, where two dilutions were selected based on the bench-scale experiment. The inoculation ratio was 10% microalgae (by volume) which yielded initial microalgae concentrations of 40–60 mg/L. Four average incident water surface light intensities were tested at 14:10 light–dark cycle: 234 and 384 μmol/(m2 × s) for one dilution and 317 and 454 μmol/(m2 × s) for the other dilution. The difference in the light intensity between the two dilutions was to account for the difference in the initial turbidity between the two dilutions. This variation in the initial turbidity may not have a pronounced effect on the bench-scale experiment; however, in raceway ponds, light will likely be the limiting factor. Light intensity was measured using an LI-COR 190 quantum sensor connected to LI-250 A light meter (https://www.licor.com). Algal biomass concentration was quantified in triplicate by the frequent measurement of OD 680 and OD 750 as indicated earlier. The samples were then stored in the fridge at 4 °C and allowed to settle for approximately 12 h, and the nutrients’ concentrations were measured from the supernatant.
Results and discussion
Biomass and optical density calibration
The biomass concentration of the microalga S. dimorphus expressed as TSS was correlated with the OD 680 as shown in Fig. 2a. As stated earlier, the biomass concentration estimated using this approach is not accurate due to the interference by the AD supernatant. Hence, 12 solutions were prepared by mixing different concentrations of microalgae and AD supernatant. The TSS of each component (microalgae or AD supernatant) was measured separately and the OD 680 and OD 750 values were measured for the 12 solutions of combined microalgae and AD supernatant. The calibration shown in Fig. 2b was established between OD 680/OD 750 and the ratio between the accurate predetermined microalgae biomass concentration and the inaccurate concentration estimated by the equation in Fig. 2a. In other words, the biomass concentration can be given by the following equation:

Biomass concentration and optical density calibration: a OD 680 and TSS calibration for the microalga S. dimorphus, b OD 680/OD 750 vs. actual microalgae biomass fraction
Bench-scale cultivation of S. dimorphus
The growth of S. dimorphus using diluted AD supernatant on the bench scale is shown in Fig. 3a. In the beginning of the experiment, all vials were not significantly different from each other at 95% confidence level. It was clear that the initial dark turbidity expressed as OD 680 was caused by the AD supernatant, especially in the highly concentrated solutions as shown in Fig. 3b. This brown turbidity decreased in the first 4–8 days and then the color began turning green as a result of the algal growth. After 4 days of monitoring, the highest biomass concentration observed was for the 0.6% dilution which was significantly higher than all other dilutions except the 1.25% dilution. Two days later, the maximum biomass concentration was observed in the 1.25% dilution which was significantly higher than all other dilutions and this remained to be the case until 8 days. After 8 days, the growth in the less concentrated dilutions (≤1.25%) started to slow down while the more concentrated solutions continued to grow under either exponential or linear growth mode. From 10 to 13 days, the maximum biomass concentration was observed in the 2.5% dilution; however, the biomass concentration at this dilution was not significantly different from those observed in the 1.25, 3.75, and 5% dilutions. As a result, it appears that the lower diluted solutions resulted in faster growth at an earlier stage and reached the stationary phase faster than the more concentrated dilutions, which can be explained by the potential chemical inhibition in the more concentrated solutions due to elevated concentrations of ammonia and COD. Furthermore, by increasing the concentration of the supernatant solution, the turbidity increased as shown in Fig. 3b, where the initial OD 680 ranged from 0.1 for the 0.05% dilution to 0.76 for the 10% dilution. This turbidity can decrease the light penetration by more than 30% in the highly concentrated solutions (10 and 7.5% dilutions), while the light attenuation is significantly lower in the less concentrated solutions.

Microalgae growth using diluted anaerobic digestate supernatant: a biomass concentration (mg/L), b OD 680
The maximum biomass concentrations corresponding to each dilution are shown in Table 1. These biomass concentrations were achieved after 20–28 days for the 2.5–10% dilutions and after 1–11 days for the 0.05–1.25% dilutions. The highest biomass concentration of 667 mg/L which was observed in the 10% dilution was significantly higher than all other dilutions except the 5 and 7.5%. It is clear also from Table 1 that there was essentially no growth in the 0.05 and 0.1% dilutions and that nutrients may become limited below the 0.6% dilution. Even though the maximum specific growth rate of 0.432 day−1 was observed in the 0.6% dilution, the exponential growth phase only extended until 4 days, after which the growth entered the stationary phase and the biomass concentration declined thereafter. Furthermore, the low biomass concentration observed in this dilution necessitates utilizing a more concentrated dilution. For instance, the 1.25% dilution, which had the second highest specific growth rate, resulted in 80% higher biomass concentration compared to the 0.6%, and the exponential growth extended to 6 days. The 2.5% dilution resulted in further increase in the biomass concentration and longer growth. Higher biomass concentrations were achieved in the more concentrated dilutions; however, the specific growth rates were lower and the initial turbidities were higher compared to the 1.25 and 2.5% dilutions. As a result, it was hypothesized that 50–100 mg N/L range provides sufficient nutrients to maximize the specific growth rate of the microalga S. dimorphus while still producing a high concentration of biomass. This conclusion is in agreement with many studies in the literature. For example, Wang et al. [21] indicated that high nitrogen concentrations such as those witnessed in a full strength BG-11 may have inhibitory effects on the growth of S. dimorphus. They also recommended an optimum nitrogen concentration range of 64–128 mg N/L for biomass and lipid. On the other hand, Xu et al. [29] reported that the optimum nitrogen concentration for the microalga S. dimorphus was 152 mg N/L when a manure wastewater was used as a nutrient medium. Goswami and Kalita [22] cultivated the microalga S. dimorphus using BG-11 medium and by replacing nitrate with urea as a nitrogen source. They concluded that urea concentration of 0.1 g/L, that is 46.6 mg N/L, was optimum for the biomass growth with a maximum specific growth rate of 0.54 day−1.
Cultivation of S. dimorphus in raceway ponds
The effect of paddlewheel mixing was evaluated by testing the growth of S. dimorphus at different water surface velocities. Sufficient mixing is necessary to maintain the algal suspension throughout the depth of light penetration and prevent thermal stratification [30, 31]. According to Hadiyanto et al. [31], 0.1–0.3 m/s water surface velocity is sufficient to maintain good mixing in the algal culture. Lower velocities result in poor mixing which leads to stagnant zones and may eventually result in anaerobic bacteria propagation and potentially toxic compounds. On the other hand, higher velocities may harm the growth, especially for cells that are sensitive to hydrodynamic stress. Hence, S. dimorphus growth was evaluated at 0.1, 0.2, and 0.3 m/s water surface velocities. As shown in Fig. 4, there was a clear impact of the mixing velocity on the growth of S. dimorphus; as the maximum specific growth rates corresponding to the 0.3, 0.2, and 0.1 m/s velocities were 0.294, 0.272, and 0.198 day−1, respectively. Additionally, the maximum biomass concentrations achieved were 761, 669, and 450 mg/L for the 0.3, 0.2, and 0.1 m/s velocities, respectively. As a result, the ponds were operated at 0.3 m/s surface water velocity for all the following experiments.

S. dimorphus growth at different mixing velocities
Based on the results of the bench-scale experiment, the microalga S. dimorphus was cultivated in 100 L raceway ponds on four separate tests using 2.5 and 1.25% diluted AD supernatant that is equivalent to 100 and 50 mg N/L, respectively. As indicated in “Materials and methods”, two light intensities were tested for each dilution: 317 and 454 μmol/(m2 × s) for the 2.5% dilution and 234 and 384 μmol/(m2 × s) for the 1.25% dilution. The initial characterization of the nutrient media for the four cultivation conditions is presented in Table 2, and the growth of the microalga S. dimorphus under these four conditions is shown in Fig. 5a.

S. dimorphus growth in the raceway ponds under different dilutions and light intensities: a biomass concentration (mg/L), b OD 680
As expected, light intensity clearly impacted the growth of the microalga S. dimorphus, particularly in the first 4 days; as higher biomass concentrations were achieved under higher light intensity regardless of the dilution. This is in agreement with Wang et al. [21] who indicated that for the microalga S. dimorphus, the specific growth rate increased linearly with an increase in the light intensity until reaching a saturation light intensity of 510 μmol/(m2 × s). On the other hand, it was reported by Xu et al. [29] that the optimal light intensity for the growth of S. dimorphus is 238 μmol/(m2 × s); however, growth under the 1.25% dilution at 234 μmol/(m2 × s) light intensity in this experiment yielded the lowest biomass concentrations among all other conditions, which suggests that this light intensity is far from the optimum for the growth of this microalga in a raceway pond using a turbid media such as the diluted AD supernatant. It was demonstrated earlier that remarkable biomass concentrations have been reported in the literature for this microalga. However, the experimental conditions differ from those adapted in our experiments. For example, Wang et al. [21] stated that the highest S. dimorphus biomass concentration achieved was 6.5 g/L. However, their experiments were conducted outdoors in 1.9–7.6 cm deep glass panel photobioreactors with a peak light intensity of 1600 μmol/(m2 × s). Similarly, Jiang et al. [16] reported that the highest S. dimorphus biomass concentration was 5.17 g/L which was achieved in 3–5 cm diameter columns. The low biomass yield in our experiments compared to the previous studies can be explained simply by the high depth of liquid (20 cm), and the lower light intensities combined with the high turbidity caused by the suspended solids in the nutrient medium, especially when the microalgae were cultivated using the 2.5% dilution with OD 680 as high as 1.2 (Fig. 5b). For the 1.25% dilution, growth rate was likely limited by either low light penetration from increasing biomass concentration, nitrogen limitation, and/or phosphorus limitation particularly after 4 days when the nitrogen and phosphorus concentrations dropped below 30 mg N/L and 3 mg P/L, respectively (Figs. 5, 6a, b). This conclusion matches the findings in Table 1 where it appears that at 0.6% dilution or below, which is equivalent to 24 mg N/L nitrogen concentration and 2.9 mg P/L phosphorus concentration, nutrients became limiting. It is not clear, however, whether a single or multiple nutrients were limiting. Many studies have investigated the effect of limiting nutrients on the algal growth. Kunikane et al. [32] and Kunikane and Kaneko [33] reported that it is necessary to consider the multiplicative effect of nitrogen and phosphorus on the growth of the microalga S. dimorphus. On the other hand, Rhee [34] reported that the multiplicative or additive effect of nitrogen and phosphorus was not applicable to the microalga S. dimorphus.

Nutrients concentrations in the S. dimorphus cultures grown in the raceway pond: a nitrogen, b phosphorus, c ammonia, d COD
The highest biomass concentration in our experiments was 446 mg/L which was observed after approximately 10 days of cultivation using the 2.5% dilution and under a light intensity of 454 μmol/(m2 × s). The second highest biomass concentration was 376 mg/L for the same dilution but with 317 μmol/(m2 × s) which was close to the biomass concentration achieved using the 1.25% dilution at 384 μmol/(m2 × s) (349 mg/L). Finally, the highest biomass concentration achieved in the 1.25% dilution and under 234 μmol/(m2 × s) light intensity was 159 mg/L. The biomass productivities were calculated between the initial inoculation time and the time the maximum biomass concentration was achieved. These productivities were 42.4, 33.1, 28.3, and 9.2 mg/(L × day) for the 2.5% dilution—454 μmol/(m2 × s), 2.5% dilution—317 μmol/(m2 × s), 1.25% dilution—384 μmol/(m2 × s), and 1.25% dilution—234 μmol/(m2 × s) cultures, respectively. However, the corresponding productivities for the first 4 days of cultivation were 67.8, 40.3, 54.8, and 28.3 mg/(L × day), respectively, which suggests that a significant improvement to the biomass productivity can be achieved by a continuous or semi-continuous cultivation mode compared to the batch cultivation which was followed in our experiments where nutrients were added only in the beginning of cultivation.
Besides the cultivation of S. dimorphus for biofuels production, this microalga has gained considerable interest for its ability to remove nutrients from wastewater. As shown in Fig. 6a, the total nitrogen removal efficiencies were in the 65–72% range for all four conditions. Ammonia was completely removed in all four conditions. However, it appears that higher light intensity resulted in faster ammonia uptake; as there was a sharp drop in the ammonia concentration in the first four days for the 384 and 454 μmol/(m2 × s) cultures compared to the other two cultures (Fig. 6c). Ammonia can be removed from wastewater via different methods such as volatilization (stripping), nitrification and denitrification, or assimilation by microalgae. For stripping to occur, the algae culture must be alkaline and under high temperature conditions, while nitrification and denitrification are characterized by a relatively long generation time of bacteria; besides a rapid decrease in the pH when nitrification occurs [7, 10]. Throughout the cultivation in our system, the pH was maintained at 7.5 ± 0.1 and temperature was 25 °C; thus, the stripping effect can be excluded. Further, no nitrate was detected in the samples, which concludes that nitrification did not contribute to ammonia removal. Hence, it can be hypothesized that the ammonia removal was the result of microalgae uptake. This can also suggest that ammonia is a favorable form of nitrogen for this microalga compared to organic nitrogen in the nutrient medium or even residual nitrate in the microalgae inoculum. This is in agreement with Tam and Wong [7], who indicated that microalgae generally prefer ammonium over other nitrogen forms. Wang and Lan [24] revealed that ammonium consumption by the microalga N. oleoabundans was faster than nitrate. However, Vidyashankar et al. [15] reported that the microalga S. dimorphus grew better with sodium nitrate and ammonium acetate as nitrogen sources compared to ammonium carbonate and urea. Arumugam et al. [35] reported similar results when they observed that the microalga S. bijugatus grew better with potassium nitrate and sodium nitrate as nitrogen sources compared to urea and ammonium nitrate.
N/P ratio is an important aspect in the microalgae cultivation. According to Klausmeier et al. [36], the atomic N/P ratio for phytoplankton varies between 8.2 and 45, and that in general, the cellular stoichiometry is flexible based on culture conditions. Rhee [34] reported that for Scenedesmus sp., the optimal atomic N/P ratio for cell density was 30, below which nitrogen becomes limited, while above this ratio phosphorus becomes limited. Kunikane et al. [32] found that the specific growth rate of the microalga S. dimorphus was independent of phosphorous concentration at TN/TP ratios of 2–10 that is equivalent to an atomic ratio of 4.4–22.1. In our experiments, the atomic N/P ratios were in the range 14.4–28.3 except for the 1.25% dilution (384) when the phosphorus was entirely consumed towards the end of cultivation which increased the ratio to over 50. Similarly, when the phosphorus was nearly depleted in the 1.25% dilution (234), the N/P ratio increased to 40. The phosphorus removal efficiencies were 63, 63, 76, and 100% for the 2.5% dilution (317), 2.5% dilution (454), 1.25% dilution (234), and 1.25% dilution (384), respectively. This suggests that lower initial phosphorus concentrations in the diluted supernatant yielded higher removal efficiencies.
The existence of BOD or COD in a water sample implies the requirement of oxygen by microorganisms to oxidize the organic matter; thus, high BOD or COD concentrations indicate the potential of dissolved oxygen depletion in the receiving waters. The COD removal efficiencies ranged from 78 to 82% in all four cultures. Given the fact that microalgae provide oxygen, COD can be removed as a result of the mixed culture of microalgae and bacteria in an open pond using unsterilized digestate as a nutrient medium [6].
In summary, utilizing this microalga can be an effective way of removing nutrients from wastewaters as well as producing a biomass for biofuels synthesis and other applications. However, in order to increase the efficiency of the process, light should be provided at higher intensities compared to those provided in this study. It is apparent from the nutrients analysis that light is likely to be the limiting factor when the microalga S. dimorphus was cultivated in 100 mg N/L AD supernatant dilution. However, at least one nutrient was limiting after 4 days of cultivation when the 1.25% dilution was used as a nutrient medium. Thus, by increasing the light intensity while using 100 mg N/L, it is expected that nitrogen and phosphorus will be removed at higher efficiencies concurrent with higher biomass concentrations and growth rates. On the other hand, continuous cultivation as opposed to batch can be achieved effectively using the 1.25% dilution; which is expected to increase the biomass productivity.
Outdoor cultivation is an attractive option for summer cultivation in a greenhouse, where higher light intensities are available for the algal growth. Light intensities during summer days can peak at 1500 μmol/(m2 × s) or higher in Ohio. Further, the increased temperature can enhance the growth rate of this microalga. Even though many studies focused on the 20–25 °C temperature as the optimal range for biomass and lipid accumulation, Benider et al. [37] reported that the optimum growth for the microalga S. dimorphus was achieved at 35 °C. Further, they revealed that the optimal light intensity for the cultivation of this microalga increases with an increase of the temperature up to 35 °C.
Conclusions
In this study, the microalga S. dimorphus was cultivated phototrophically using diluted anaerobic digestate supernatant. A bench-scale experiment was conducted over a wide range of dilutions, and it was found that 50–100 mg N/L dilutions provided sufficient nutrients to optimize the microalgal growth on a larger scale. The cultivation was scaled up to 100 L raceway ponds. Two dilutions (100 and 50 mg N/L) were selected and each dilution was tested under two light intensities. The highest biomass concentration achieved in the bench-scale experiment was 667 mg/L, whereas in the raceway ponds the highest biomass concentration was 446 mg/L. Furthermore, it was found that the nitrogen removal efficiency was 65–72%, whereas ammonia was completely removed from the nutrient media. Phosphorus was removed at 63–100% efficiencies, while COD removal efficiencies were in the 78–82% range. It is believed that summertime outdoor cultivation will improve the biomass productivity and nutrients removal efficiencies due to the significant increase in light intensity.
References
Terry KT, Raymond LP (1985) System design for the autotrophic production of microalgae. Enzyme Microb Technol 7:474–487
Chen Y, Wang J, Zhang W, Chen L, Gao L, Liu T (2013) Forced light/dark circulation operation of open pond for microalgae cultivation. Biomass Bioenergy 56:464–470
Mata TM, Martins AA, Caetano NS (2010) Microalgae for biodiesel production and other applications: a review. Renew Sustain Energy Rev 14:217–232
James SC, Boriah V (2010) Modeling algae growth in an open-channel raceway. J Comput Biol 17:895–906
Lunka AA, Bayless DJ (2013) Effects of flashing light-emitting diodes on algal biomass productivity. J Appl Phycol 25(6):1679–1685
Choi HJ, Lee SM (2012) Effects of microalgae on the removal of nutrients from wastewater: various concentrations of Chlorella vulgaris. Environ Eng Res 17(S1):S3–S8
Tam NFY, Wong YS (1990) The comparison of growth and nutrient removal efficiency of Chlorella pyrenoidosa in settled and activated sewages. Environ Pollut 65(2):93–108
Abu Hajar HA, Riefler RG, Stuart BJ (2016) Anaerobic digestate as a nutrient medium for the growth of the green microalga Neochloris oleoabundas. Environ Eng Res 21(3):265–275
Park J, Jin HF, Lim BR, Park KY, Lee K (2010) Ammonia removal from anaerobic digestion effluent of livestock waste using green alga Scenedesmus sp. Bioresour Technol 101(22):8649–8657
Tam NFY, Wong YS (1996) Effect of ammonia concentrations on growth of Chlorella vulgaris and nitrogen removal from media. Bioresour Technol 57(1):45–50
Travieso L, Benítez F, Sánchez E, Borja R, Martín A, Colmenarejo MF (2006) Batch mixed culture of Chlorella vulgaris using settled and diluted piggery waste. Ecol Eng 28(2):158–165
Renaud SM, Parry DL, Thinh LV (1994) Microalgae for use in tropical aquaculture I: gross chemical and fatty acid composition of twelve species of microalgae from the Northern Territory, Australia. J Appl Phycol 6(3):337–345
Oron G, Shelef G, Levi A (1981) Environmental phenotypic variation of Scenedesmus dimorphus in high-rate algae ponds and its relationship to wastewater treatment and biomass production. Biotechnol Bioeng 23(10):2185–2198
Frigon JC, Matteau-Lebrun F, Abdou RH, McGinn PJ, O’Leary SJ, Guiot SR (2013) Screening microalgae strains for their productivity in methane following anaerobic digestion. Appl Energy 108:100–107
Vidyashankar S, Deviprasad K, Chauhan VS, Ravishankar GA, Sarada R (2013) Selection and evaluation of CO2 tolerant indigenous microalga Scenedesmus dimorphus for unsaturated fatty acid rich lipid production under different culture conditions. Bioresour Technol 144:28–37
Jiang Y, Zhang W, Wang J, Chen Y, Shen S, Liu T (2013) Utilization of simulated flue gas for cultivation of Scenedesmus dimorphus. Bioresour Technol 128:359–364
Giannetto MJ, Retotar A, Rismani-Yazdi H, Peccia J (2015) Using carbon dioxide to maintain an elevated oleaginous microalga concentration in mixed-culture photo-bioreactors. Bioresour Technol 185:178–184
González LE, Cañizares RO, Baena S (1997) Efficiency of ammonia and phosphorus removal from a Colombian agroindustrial wastewater by the microalgae Chlorella vulgaris and Scenedesmus dimorphus. Bioresour Technol 60(3):259–262
Kang J, Wen Z (2015) Use of microalgae for mitigating ammonia and CO2 emissions from animal production operations—evaluation of gas removal efficiency and algal biomass composition. Algal Res 11:204–210
Ruangsomboon S, Ganmanee M, Choochote S (2013) Effects of different nitrogen, phosphorus, and iron concentrations and salinity on lipid production in newly isolated strain of the tropical green microalga, Scenedesmus dimorphus KMITL. J Appl Phycol 25(3):867–874
Wang L, Li Y, Sommerfeld M, Hu Q (2013) A flexible culture process for production of the green microalga Scenedesmus dimorphus rich in protein, carbohydrate or lipid. Bioresour Technol 129:289–295
Goswami RCD, Kalita MC (2011) Scenedesmus dimorphus and Scenedesmus quadricauda: two potent indigenous microalgae strains for biomass production and CO2 mitigation—a study on their growth behavior and lipid productivity under different concentration of urea as nitrogen source. J Algal Biomass Utln 2(4):42–49
Welter C, Schwenk J, Kanani B, Van Blargan J, Belovich JM (2013) Minimal medium for optimal growth and lipid production of the microalgae Scenedesmus dimorphus. Environ Prog Sustain Energy 32(4):937–945
Wang B, Lan CQ (2011) Biomass production and nitrogen and phosphorus removal by the green alga Neochloris oleoabundans in simulated wastewater and secondary municipal wastewater effluent. Bioresour Technol 102(10):5639–5644
Pittman JK, Dean AP, Osundeko O (2011) The potential of sustainable algal biofuel production using wastewater resources. Bioresour Technol 102(1):17–25
de Winter L, Klok AJ, Franco MC, Barbosa MJ, Wijffels RH (2013) The synchronized cell cycle of Neochloris oleoabundans and its influence on biomass composition under constant light conditions. Algal Res 2(4):313–320
Clesceri L, Greenberg A, Eaton A (1998) Standard methods for the examination of water and wastewater, 20th edn. American Public Health Association, Washington DC
James DE (1978) Culturing algae. Carolina Biological Supply Company, Burlington, NC
Xu X, Shen Y, Chen J (2015) Cultivation of Scenedesmus dimorphus for C/N/P removal and lipid production. Electron J Biotechnol 18(1):46–50
Green FB, Lundquist TJ, Oswald WJ (1995) Energetics of advanced integrated wastewater pond systems. Water Sci Technol 31:9–20
Hadiyanto H, Elmore S, Van Gerven T, Stankiewicz A (2013) Hydrodynamic evaluations in high rate algae pond (HRAP) design. Chem Eng J 217:231–239
Kunikane S, Kaneko M, Maehara R (1984) Growth and nutrient uptake of green alga, Scenedesmus dimorphus, under a wide range of nitrogen/phosphorus ratio—I. Experimental study. Water Res 18(10):1299–1311
Kunikane S, Kaneko M (1984) Growth and nutrient uptake of green alga, Scenedesmus dimorphus, under a wide range of nitrogen/phosphorus ratio—II. Kinetic model. Water Res 18(10):1313–1326
Rhee GY (1978) Effects of N:P atomic ratios and nitrate limitation on algal growth, cell composition, and nitrate uptake. Limnol Oceanogr 23(1):10–25
Arumugam M, Agarwal A, Arya MC, Ahmed Z (2013) Influence of nitrogen sources on biomass productivity of microalgae Scenedesmus bijugatus. Bioresour Technol 131:246–249
Klausmeier CA, Litchman E, Daufresne T, Levin SA (2004) Optimal nitrogen-to-phosphorus stoichiometry of phytoplankton. Nature 429(6988):171–174
Benider A, Tahiri M, Belkoura M, Dauta A (2001) Interaction des facteurs héliothermiques sur la croissance de trois espèces du genre Scenedesmus. Ann Limnol 37(4):257–266
Acknowledgements
This work was funded entirely by the National Science Foundation (NSF) through the Sustainable Energy Pathways (SEP) program (Award # 1230961).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors of this manuscript declare that they have no competing interests.
Rights and permissions
About this article

Cite this article
Abu Hajar, H.A., Riefler, R.G. & Stuart, B.J. Cultivation of Scenedesmus dimorphus using anaerobic digestate as a nutrient medium. Bioprocess Biosyst Eng 40, 1197–1207 (2017). https://doi.org/10.1007/s00449-017-1780-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00449-017-1780-4