
Abstract
This study investigated the effect of enzymatic and combined alkaline-enzymatic pretreatments on chemical composition and methane production from ensiled sorghum forage. Four commercial enzymatic preparations were tested and the two yielding the highest sugars release were added to evaluate any hydrolytic effect on both untreated and alkaline pretreated samples. In the combined alkaline-enzymatic pretreatment trials, the highest sugar release was found with Primafast and BGL preparations (added at a final concentration 0.12 and 0.20 mL/g TS, respectively), with a total monomeric content of 12 and 6.5 g/L. Fibre composition analysis confirmed that the combined alkaline-enzymatic pretreatment led to cellulose (up to 32 %) and hemicelluloses (up to 56 %) solubilisation, compared to the enzymatic pretreatment alone. BMP tests were performed on both untreated and pretreated samples, and time courses of methane production were fitted. Both enzymatic and combined alkaline-enzymatic pretreatment led to a methane production increase (304 and 362 mL CH4/g VS), compared to that of untreated sorghum (265 mL CH4/g VS), as +15 and +37 %, respectively. Moreover, higher specific methane production rates, compared to that of untreated sorghum (20.31 mL CH4/g VS/d), were obtained by applying the enzymatic and combined alkaline-enzymatic pretreatment (33.94 and 31.65 mL CH4/g VS/d), respectively.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The production of methane from lignocellulosic materials (i.e. agricultural residues and energy crops) appears to be one of the most promising alternatives to fossil fuels with no “food versus fuel” dilemma. Sorghum, a multi-purpose crop that can be cultivated under a wide range of environmental conditions, with a world cultivated land of 40 million ha in 2009 and with a hectare yield as high as 25 t (dry weight) per year, represents an interesting substrate for methane production [1]. In particular, ensiled sorghum forage (sorghum sudanensehybrid) is one of the most commonly used biomass in Italian agricultural biogas plants [2].
As every lignocellulosic substrate, methane production from sorghum depends mainly on its complex structure that limits its biodegradability. Indeed, sorghum is mainly composed of cellulose, hemicelluloses and lignin, strongly linked to each other. Cellulose and hemicelluloses (holocelluloses), which are the major components of most lignocellulosic materials, are quite easily degraded by anaerobic microorganisms and can be converted into methane. Biological degradation of holocelluloses within an anaerobic digester is normally facilitated by enzymes, such as cellulases and hemicellulases, secreted by microorganisms [3]. Nevertheless, the crystalline part of cellulose, as well as the presence of lignin, limits holocelluloses accessibility to hydrolytic enzymes [4, 5]. Therefore, several pretreatment methods have been studied to alter the complex structure of lignocellulosic materials, thus increasing the hydrolysis rate and thereafter the anaerobic digestion efficiency.
Pretreatments include mechanical, chemical, thermal, biological processes or a combination of them [6]. Biological pretreatment processes have received considerable attention because they can be considered energy-saving and environmentally friendly methods. Moreover, enzymatic hydrolysis can present several advantages over chemical hydrolysis, such as the high substrate and reaction specificity and the possibility to operate under mild conditions without generating by-products, mainly furfural and 5-HMF. Nevertheless, potential loss of carbohydrates during the process and the high cost of industrial enzymes are actually the major disadvantages for enzymatic pretreatment [7, 8].
Biological pretreatments can be performed by applying either commercial enzymes or fungi to lignocellulosic materials. Commercial enzymes, such as cellulase (endoglucanase, exoglucanase and β-glucosidase), xylanase, pectinases (poly-galcaturonase and pectate-lyase) or lignolytic enzymes (laccase, lignin and manganese peroxidase), are industrially synthesised by a variety of microorganisms (i.e. fungi and bacteria) grown on specific organic substrates. They are generally characterised by high enzymatic activities and they can be used to breakdown all components of lignocelluloses, including lignin, the polymer most refractory to microbial attack [9, 10].
Up to now, some studies have investigated the effects of enzymatic pretreatments on anaerobic digestion of lignocellulosic substrates. Wheat grains pretreated with Trizyme (cellulase, alpha-amylase and protease mix) at 37°C for 24 h before anaerobic digestion increased methane production by 14 % [11]. An enzymatic pretreatment applied to grass at 35°C for 24 h using two xylanases (GC 320 and Multifect) and two cellulases (IndiAge MAX L and Primafast 200), led to a 22 % increase in methane yield [12]. Other authors noticed that lignolytic enzymes can increase methane production potentials, and an energy gain of 28 and 42 % was reached using lignin peroxidase and manganese peroxidase, respectively [13].
However, as stated before, the enzymatic accessibility of holocelluloses for their further conversion into methane is mainly limited by lignin content and cellulose crystallinity. Consequently, additional pretreatments need to be applied prior to the enzymatic attack to reduce this strong physical barrier and enhance enzymatic hydrolysis of holocelluloses. Alkaline pretreatments are known to remove efficiently lignin and the lignin − hemicellulose complexes and, thus, can increase enzymatic hydrolysis of holocelluloses [14]. Combining alkaline pretreatment with enzymatic hydrolysis appears to be a promising approach to enhance methane production from lignocellulosic substrate for both environmental and energy reasons. Indeed, alkaline pretreatment was shown to be effective at low temperature, similar to that of enzymatic pretreatment, thus permitting low energy consumption. Moreover, working at high pH levels the generation of toxic effluents (i.e. HMF/furfurals) is avoided [14, 15].
Thus far, alkaline pretreatment followed by enzymatic hydrolysis has been widely investigated for bioethanol and biohydrogen production [16–20]. According to authors’ knowledge, up to date no studies were found on the application of a combined alkaline-enzymatic pretreatment to enhance methane production from sorghum.
Thus, the objective of this study was to evaluate the performance of an enzymatic or a combined alkaline-enzymatic pretreatment on methane production from ensiled sorghum forage, with the aim of identifying any possible synergic effect of the two pretreatments on methane production yield. Four commercial enzymatic preparations were initially tested; those showing the highest sugars releases were employed together on untreated and alkaline pretreated sorghum. Biochemical Methane Potential (BMP) tests were also performed to evaluate the effect of the enzymatic and combined alkaline-enzymatic hydrolysis on specific methane production rates.
Materials and methods
Substrate
Ensiled sorghum forage (Trudan 8) was collected from a farm near Cremona (Lombardy Region, Italy). It was air dried and ground into 1 mm particle size by a kitchen blender and finally stored at 20 °C in air-tight containers prior to use.
Alkaline pretreatment
Alkaline pretreatment conditions were chosen according to the best pretreatment result of our previous study performed on ensiled sorghum forage [14]. Trials were carried out in a 500 mL digestion flask, closed with a rubber septum, in which sorghum sample was soaked in a sodium hydroxide (NaOH) solution (10 g NaOH/100 g TS). The initial Total Solids (TS) concentration was 160 g TS/L. To maintain pretreatment at the desired temperature (40°C for 24 h), the flask was put in a thermostatic incubator without stirring.
Enzymatic pretreatment
First, to define the optimal enzymatic mixture to perform pretreatment tests, four commercial enzymatic preparations were characterised in terms of enzymatic activities: Agazym BGL and Ultra L (Garzanti Specialties), Pulpzyme HC (Novo Nordisk) and Primafast 200 (Genencor Inc.).
Agazym BGL is especially formulated to favour the breakdown of plant cell walls to extract tissue components during industrial processing of cereals. It is an enzymatic mix characterised by cellulose, β-glucanase, Hemicellulase and Xylanase from Aspergillus aculeatus.
Ultra L is recommended to perform alcoholic fermentation of red wines, when must is fermented in contact with grape husk, to facilitate the extraction of pigments and flavours. It is mainly characterised by polygalacturonase and pectinase from Aspergillus strains.
Pulpzyme HC is used during the process of bleaching and deinking for the production of recycled paper. It is characterised by endo-xylanases from Bacillus strains.
Primafast 200 is recommended for clothes processing such as depilling and softening and to obtain the so-called “stone-washed look”. It is characterised by endo-1-4-β-glucanases.
Then, each enzymatic preparation was put in contact with alkaline pretreated sorghum at a final concentration of 0.12 mL/g TS for Primafast, 0.20 mL/g TS for BGL and at 0.04 mg/g TS for Ultra and Pulpzyme, following the information present in each technical datasheet provided. Then, H2O was added to reach a total solids (TS) concentration of 70 g TS/L. The enzymatic activities (IU/g VS) used in these trials are reported in Table 2. Samples were incubated at each appropriate pH (7.0 for Pulpzyme HC and Agazym Ultra L, 5.0 for Agazym BGL and Primafast) and temperature (50°C for Pulpzyme, Primafast and BGL, 20°C for Ultra L) under stationary conditions. After 24 h incubation, samples were collected, centrifuged and the supernatant stored at −18 °C until analysis.
According to results obtained, Agazym BGL and Primafast 200 were chosen to perform pretreatments.
Enzymatic pretreatments were performed on untreated and alkaline pretreated ensiled sorghum forage. Trials were performed in 500 mL digestion flasks, closed with a rubber septa. In each flask, the enzymatic preparations were added to each substrate at a final concentration of 0.20 and 0.12 mL/g TS for BGL and Primafast, respectively. Then, H2O was added to reach a total solids (TS) concentration of 70 g TS/L and pH was corrected at 5.0 with HCl. Samples were then incubated at 50°C for 72 h in a thermostatic incubator under stationary conditions. Samples were then filtered through a sieve of 0.20 mm of pore size and the separated solid fraction was taken for compositional analyses.
Analytical determinations
Total solids (TS), volatile solids (VS) and chemical oxygen demand
Total Solids (TS), Volatile Solids (VS) and Chemical Oxygen Demand (COD) were determined according to the APHA Standard Methods [21].
Fats and proteins determination
Fats and proteins of untreated sorghum were determined with a NIR System (5000 Monochromator, Foss).
Determination of cellulose (CEL), hemicelluloses (H-CEL) and klason lignin (K-LIG) content
Cellulose (CEL), hemicelluloses (H-CEL) and klason lignin (K-LIG) were measured using a strong acid hydrolysis method adapted from [22]. Samples (200 mg) were first hydrolyzed with 12 M H2SO4 acid for 2 h at room temperature, then diluted to reach a final acid concentration of 1.5 M and kept at 100°C for 3 h. The insoluble residue was separated from the supernatant by filtration on fibreglas filter (GF/F, WHATMAN), washed with 50 mL of deionized water and then placed in a crucible. The crucible and the paper fibreglass were dried at 100°C during 24 h to determine by weighting the amount of klason lignin. The supernatant, after centrifugation of the sample in 2 mL Eppendorf® tubes, was filtered at 0.2 μm (Nylon membrane, Acrodlsc®). An aliquot (800 μL) was then transferred in a vial prior to the analysis by high-pressure liquid chromatography (HPLC). Structural carbohydrates (i.e. glucose, xylose, arabinose, glucuronic and galacturonic acids) were measured through an HPLC system coupled to refractometric detection (Waters R410). The components were separated by an Aminex column HPX-87H column (300 × 7.8 mm, Bio-Rad) equipped with a protective precolumn (Microguard cation H refill cartbridges, Bio-Rad). The eluting solution corresponded to 0.005 M H2SO4, and the flow rate was 0.3 mL/min. The column temperature was maintained at 50°C and the refractometric temperature was fixed at 45°C. A refractive index detector (Waters 2414) was used to quantify the carbohydrates. The system was calibrated with glucose (0–6 g/L), xylose (0–6 g/L), arabinose (0–2 g/L) and uronic acid (0–2 g/L) (galacturonic and glucuronic) standards (Sigma–Aldrich®). Thereafter, cellulose and hemicelluloses contents were estimated as follows (Eqs. 1, 2):
where 1.11 is the ratio of the molecular weights of glucose to glucan (180/162) and 1.13 is the ratio of the molecular weights of xylose and arabinose to xylan (150/132).
Determination of enzymatic activities
Endoglucanase (CMCase) enzymatic activity was determined by measuring the amount of glucose released from carboxymethylcellulose (CMC) using the Somogyi-Nelson method with glucose as standard [23]. For this purpose an aliquot of diluted sample (0.5 mL) was mixed with 0.5 mL of a CMC suspension (1 % w/v) in citrate buffer (0.05 mol/L, pH 5). Reaction mixtures were left at 55°C for 30 min and then boiled to stop the enzymatic activity. Sugar release was then determined with glucose as standard. One unit of enzyme (IU) was defined as the amount of enzyme which hydrolyzes 1 μmol of reducing sugars, expressed as glucose, in 1 min.
Exoglucanase (Avicelase) enzymatic activity was determined according to [24]. An aliquot of diluted sample (1 mL) was mixed with 1 mL of a suspension containing microcrystalline cellulose Avicel® (2 % w/v) in acetate buffer (0.1 M, pH 5). Samples were then incubated at 30°C for 24 h and then boiled to stop the enzymatic activity. The amount of glucose released from cellulose Avicel® was measured according to Somogyi-Nelson method with glucose as standard. One unit of enzyme (IU) was defined as the amount (μmol) of glucose released from 1 mL of sample, in 1 min.
Xylanase enzymatic activity was determined employing a procedure adapted from Shewale and Sadana [25] by mixing 0.5 mL of sample with the same volume of a xylan solution (1 % w/v) in citrate buffer (0.025 M, pH 5). A blank sample with 0.5 mL of deionized water was also prepared. Samples were incubated at 50°C for 30 min and then boiled to stop the enzymatic activity.
Reducing sugars were determined again through the Somogyi procedure, employing xylose as standard. One unit of enzyme (IU) was defined as the amount which releases 1 μmol of reducing sugar (either glucose or xylose) equivalent per min under the conditions specified above.
Biochemical methane potential (BMP) assay
Biochemical Methane Potential (BMP) tests were performed in duplicate, using a commercial laboratory instrument (AMTPS, Bioprocess control, Sweden). This volumetric device consists of 15 gas-tight glass bottles (500 mL working volume) placed in a water bath at 35 ± 0.5 °C. Each bottle is continuously mixed with a rotary stirrer; the BMP test duration was 30 days.
The produced biogas passes through a 3 M NaOH solution for CO2 absorption. Methane flows through an automated liquid-displacement measuring unit with a resolution of 11–13 mL. A data acquisition system allows flow-rate data to be recorded continuously.
The inoculum (“MIX”) used for BMP tests was obtained by mixing two digested sludge samples: (1) collected from a digester fed on waste activated sludge, with a total and volatile solid content of 20 ± 4 g TS/L and 12 ± 2 g VS/L, respectively; (2) collected from a digester fed on agro-wastes (cattle manure and corn silage), with a total and volatile solid content of 55 ± 3 g TS/L and 37 ± 2 g VS/L, respectively. The mixture was made of 50 % each on a VS basis, with a final content of 15 g VS/L.
Before BMP tests, the inoculum was kept under endogenous anaerobic conditions at 35 °C for about 7 days to reduce non-specific biogas generation. A sample amount of 0.75 gVS (untreated or enzymatic pretreated sorghum) was then mixed with 2.5 g VS of inoculum, obtaining a substrate/inoculum ratio around 0.5 gVS/g VS. Finally, to reach 500 mL of working volume, 50 mL of mineral macronutrients medium and deionised water were also added to each bottle, as suggested by the OECD Guidelines for the Testing of Chemicals [26]. A blank sample was performed by mixing inoculum, mineral medium and deionised water without substrate addition. As mentioned earlier, enzymatic pretreatment was performed at pH 5.5; after inoculum, mineral medium and water addition, no pH adjustment was performed as sample pH was around 7.
The Biochemical Methane Potential (BMP) production was calculated as follows (Eq. 3):
where (VCH4,s−VCH4,blank) (NmLCH4) is the net volume (at 0°C and 1 atm) of methane production measured at the end of the test, and VSs (g VS) is the mass of volatile solids of sorghum in each BMP bottle.
Time course of methane production in each trial was fitted by employing the Excel add-in DMFit software, to estimate methane production rate (mLCH4/(g VS/d), according to Baranyi and Roberts [27] model. This equation (Eq. 4), first set up to model bacterial growth, in recent years has been adopted over the modified Gompertz model [28]:
In the present paper the equation was applied to model methane production, where y(t) = mL CH4/g VS; y 0 = mL CH4/g VS present at t 0; y max = (mL CH4/gVS) max ; µ max is the maximum methane specific production rate (mL CH4/g VS/d); m is a curvature parameter to characterise the transition from the exponential phase; v is a curvature parameter to characterise the transition to exponential phase; and h 0 = v, as suggested by the authors when fitting situations other than bacterial viable counts.
Results and discussion
Chemical composition of untreated sorghum
Chemical composition of untreated ensiled sorghum forage is summarised in Table 1. Despite the high variability of substrates composition, varying according to plant type and variety, results can be considered in accordance with literature values. Forage sorghum shows cellulose content between 32–26 % TS, and hemicelluloses and lignin values variable between 20–23 % TS and 18–26 % TS, respectively [ [29], [30] ].
Characterisation and selection of commercial enzymatic preparations
Enzymatic preparations were first characterised in terms of endoglucanase, exoglucanase (avicelase) and xylanase activities. BGL was found to contain 235.7 ± 24.3 IU/mL endoglucanase activity and 126.5 ± 10.6 IU/mL xylanase. Agazym Ultra L was composed mainly by endoglucanase, 613.2 ± 42.9 IU/mL, while Pulpzyme by xylanase, 106.8 ± 1.9 IU/mL. Primafast is a highly concentrated preparation containing 2063.4 ± 0.8 IU/mL endoglucanase and 282.8 ± 5.7 IU/mL xylanase. Exoglucanase activity in all samples was found in traces (max 3.5 IU/mL in Primafast).
In a previous paper, the effects of several chemical and physical pretreatments of sorghum forage to increase biomethane production were investigated: results highlighted that alkaline pretreatment, with 10 g NaOH/100g TS, at 40°C for 24 h, produced the highest increase in methane yield (up to 32 %), compared to the untreated substrate [14].
With the aim of further improving biogas production, alkaline pretreated sorghum was thus selected as substrate to perform the enzymatic pretreatment. Alkaline sorghum samples were singularly added with the four commercial enzymatic preparations. Trials were performed with an initial total solid substrate concentration of 70 g TS/L, corresponding to an initial volatile solids concentration of 61 g VS/L. In Table 2 the resulting xylanase, endoglucanase and exoglucanase activities concentrations, expressed in terms of IU/g VS of the substrate, are reported. Comparing these data with the enzymatic activities physiologically present inside the MIX inoculum and reported elsewhere [3], it was possible to highlight that these reached enzymatic concentrations resulted hundred or thousand times higher than those naturally found in the MIX inoculum during the course of BMP test. Indeed, the maximum values of xylanase activity, 5 and 1.5 IU/g VS, were found on day 9 of BMP tests in the presence and absence of sorghum, respectively. Maximum value of exoglucanase activity was found on day 9 of BMP tests in absence (0.04 IU/g VS) and on day 35 in the presence of sorghum (0.08 IU/g VS) as substrate. Endoglucanase was always found in trace during all the time of BMP tests [3].
After 24 h incubation at each appropriate pH and temperature, liquid fractions were analysed for their monomeric sugar content. Results are reported in Table 3. The highest sugar release was found with Primafast and BGL with a total monomeric content of 12 and 6.5 g/L, respectively, while Pulpzyme and Ultra L preparations were not active on the studied substrate.
Because of the higher degree of hydrolysis observed, Primafast and BGL were chosen for the prosecution of the research. The two preparations were added together not only to alkaline pretreated sorghum, but also to untreated sorghum comparatively; samples were incubated for up to 72 h.
Effect of enzymatic pretreatment on the fibrous composition
Fibre composition changes induced by enzymatic and combined alkaline-enzymatic pretreatments on ensiled sorghum forage were investigated, by analysing the separated solid fraction (Fig. 1). Results about sole alkaline pretreatment were also reported for comparative purposes.

Fibrous composition of untreated and pretreated ensiled sorghum forage (results are expressed in terms of % initial VS). Values correspond to mean ± standard deviation of measurement performed in duplicate
Cellulose solubilisation (as 20 %), was observed after enzymatic pretreatment, due to the action of endoglucanase (CMCase) and exoglucanase (Avicelase) activities. On the contrary, the sole enzymatic pretreatment led to neither hemicelluloses nor lignin solubilisation. Indeed, the absence of lignin degrading enzymes into commercial preparations did not allow lignin solubilisation, thus probably avoiding the subsequent solubilisation of hemicelluloses not promptly available for the enzymatic attack.
As observed, alkaline pretreatment led to solubilisation of lignin (29 %), thus permitting a subsequent hydrolysis of both cellulose and hemicelluloses fractions. As expected, for both substrates, no further solubilisation of lignin, compared to that obtained after alkaline pretreatment alone, was observed after the combined alkaline-enzymatic pretreatment, due to the absence of lignin degrading enzymes into commercial preparations.
A further solubilisation of cellulose (32 %) and hemicellulose (56 %) fractions, compared to enzymatic pretreatment alone, was observed after the combination of pretreatments. However, by comparing these results with those of alkaline pretreatment alone, it is possible to observe that a further solubilisation of both cellulose (39 %) and hemicelluloses (44 %) was favoured by the combination with enzymatic pretreatment.
As observed by Barakat et al. [31], both physical distribution and composition of lignin can play an important role for enzyme accessibility and substrate digestibility. During alkaline pretreatment, a physical redistribution of lignin could occur and the composition of lignin could change, but this is strictly related to the type of substrate.
Other factors can also explain the increase of enzymatic hydrolysis of cellulose and hemicelluloses after alkaline pretreatment: first of all, the increase of accessible surface area and pore volume observed after alkaline pretreatment. Gharpuray et al. [32] observed an increase of the accessible surface area from 0.64 to 1.7 m2/g TS by pretreating wheat straw at 100°C with 10 % NaOH (w/w) for 30 min. The same authors showed that specific surface area can affect the digestibility of biomass: an increase in accessible surface area resulted in higher hydrolysis yield. However, the small increases of both accessible surface area (2.5 %) and pore volume (27 %), observed by Monlau et al. [33] between untreated and alkaline pretreated (4 g NaOH/100g TS, 55°C for 24 h) sunflower stalks suggest that other factors may affect the enhancement of hydrolysis yield. Among them, removal of uronic acids observed after alkaline pretreatment could improve the enzymatic hydrolysis. Indeed, Pakarinen et al. [34] showed that the removal of pectins (polymer of galacturonic acids) present in hemp can increase the enzymatic hydrolysis by 26 %.
Biochemical methane production tests
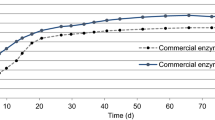
BMP tests were performed on untreated, enzymatic and combined alkaline-enzymatic pretreated samples (Fig. 2). Results of the BMP tests performed after the sole alkaline pretreatment were also reported for comparative purposes.

Methane yield (BMP, NmLCH4/gVS) trends, at normal temperature and pressure conditions, of untreated, enzymatic, alkaline and combined alkaline-enzymatic pretreated ensiled sorghum forage. Values correspond to mean ± standard deviation of measurement performed in duplicate
Methane yield of untreated sorghum was 265 ± 4 NmL CH4/g VS. By knowing the COD/VS ratio for the substrate and considering that, at normal condition, 350 mL CH4 can be obtained from 1 gCOD removed, the corresponding anaerobic biodegradability of 63 % has been obtained [35].
By performing the enzymatic pretreatment, an increase in methane yield of 15 % was observed (304 ± 11 mL CH4/g VS). According to fibrous composition analysis, this result is probably due to the effect of the added endoglucanase and xylanase, which were able to attack and solubilize the cellulosic and hemicellulosic fractions during the pretreatment. The sole alkaline pretreatment increased methane production of sorghum by 29 % (343 ± 5 mL CH4/g VS). The highest increase in methane production (37 %), compared to untreated sorghum, was observed by combining NaOH and enzymatic pretreatment (362 ± 3 mL CH4/g VS). This can be mainly due to the effect of alkaline pretreatment which led to a solubilisation of lignin, thus permitting a further hydrolysis of cellulose and hemicelluloses fractions.
Time courses of methane production were fitted employing the DMFit Excel add-in shareware package to obtain the methane specific production rates, expressed in terms of mL CH4/g VS/d of the four curves (Table 4). As regards untreated sorghum, this value was found 20.31 mL CH4/g VS/d. After the enzymatic pretreatment, specific methane production rate increased up to 33.94 mL CH4/g VS/d; a similar value, 31.65 mL CH4/g VS/d was obtained employing the combined alkaline-enzymatic pretreatment. The highest methane production rate was observed applying the alkaline pretreatment alone (47.74 mL CH4/g VS/d).
It is noteworthy that in the first 12 days the combined alkaline-enzymatic pretreated sample showed the lowest specific methane production rate. This is probably due to the presence of by-products released during the pretreatment phase that led to the necessity of adapting the microbial inoculum during BMP tests. These molecules cannot be related to phenols that are usually formed during alkaline treatments, or to furfurals, formed when high temperatures and low pH are reached [15]. Further researches are needed to identify the nature and concentration of these compounds.
Considering the final methane production achieved, 362 versus 343 NmLCH4/g VS, the performance of a combined alkaline-enzymatic pretreatment, compared with the sole alkaline, needs to be further investigated above all taking into account the cost of commercial enzymes.
In summary, the applied pretreatments led to an increase of either methane production and specific production rate compared to untreated sorghum, suggesting that enzymatic pretreatment of a lignocellulosic substrate may facilitate the hydrolytic activity of the microbial population present in sludge samples.
Conclusions
Four commercial enzymatic preparations were tested on alkaline-pretreated ensiled sorghum forage, to increase its methane production potential. The highest sugar release was found with Primafast and BGL preparations, with a total monomeric content of 12 and 6.5 g/L, respectively. Fibre composition analysis confirmed that combined alkaline-enzymatic pretreatment led to cellulose (up to 32 %) and hemicellulose (up to 56 %) solubilisation compared to enzymatic pretreatment alone. BMP tests were performed on both untreated and pretreated samples, and time courses of methane production were fitted. Both enzymatic and combined alkaline-enzymatic pretreatment led to a methane production increase, compared to that of untreated sorghum (+15 and +37 %, respectively). Higher specific methane production rates, compared to that of untreated sorghum (20.31 mL CH4/g VS/d), were obtained by applying the enzymatic and combined alkaline-enzymatic pretreatment (33.94 and 31.65 mL CH4/g VS/d), respectively.
The reported research will pave the way to the application of combined chemo-biological pretreatments to increase the methane production potential of sorghum; nevertheless, the influence of the molecules released during the hydrolytic step on the metabolism of the microbial population present in sludge samples needs to be further investigated.
References
FAO: food and agriculture organization of United States (2012). http://www.fao.org/
Appels L, Lauwers J, Degrève J, Helsen L, Lievens B, Willems K, Impe JV, Dewil R (2011) Anaerobic digestion in global bio-energy production: potential and research challenges. Renew Sustain Energy Rev 15:4295–4301
Sambusiti C, Rollini M, Ficara E, Musatti A, Manzoni M, Malpei F (2014) Enzymatic and metabolic activities of four anaerobic sludges and their impact on methane production from ensiled sorghum forage. Bioresour Technol 155:122–128
Malherbe S, Cloete TE (2002) Lignocellulose biodegradation: fundamentals and applications. Rev Environ Sci Biotechnol 1:105–114
Barakat A, de Vries H, Rouau X (2013) Dry fractionation process as an important step in current and future lignocellulose biorefineries: a review. Bioresour Technol 130:362–373
Mosier N, Wyman C, Dale B, Elander R, Lee YY, Holtzapple M, Ladisch M (2005) Features of promising technologies for pretreatment of lignocellulosic biomass. Bioresour Technol 96:673–686
Banerjee G, Scott-Craig JS, Walton JD (2010) Improving enzymes for biomass conversion: a basic research perspective. Bioen Res 3:82–92
Zhong W, Zhang Z, Luo Y, Sun S, Qiao W, Xiao M (2011) Effect of biological pretreatments in enhancing corn straw biogas production. Bioresour Technol 102:11177–11182
Lopez MJ, Vargas-Garcia MD, Suarez-Estrella F, Nichols NN, Dien BS, Moreno J (2007) Lignocellulose-degrading enzymes produced by the ascomycete Coniochaeta ligniaria and related species: application for a lignocellulosic substrate treatment. Enzyme Microb Technol 40:794–800
Howard RLE, Abotsi EL, van-Rensburg J, Howard S (2003) Lignocellulose biotechnology: issues of bioconversion and enzyme production. Afr J Biotechnol 2:602–619
Sonakya V, Raizada N, Kalia V (2001) Microbial and enzymatic improvement of anaerobic digestion of waste biomass. Biotechnol Lett 23:1463–1466
Lehtomaki A, Viinikainen TA, Ronkainen OM, Alen R, Rintala JA (2004) Effects of pre-treatments on methane production potential of energy crops and crop residues. In: Proceedings of 10th IWA World Congress on Anaerobic Digestion, 29 August–2 September, Montreal, Canada
Frigon JC, Mehta P, Guiot SR (2012) Impact of mechanical, chemical and enzymatic pre-treatments on the methane yield from the anaerobic digestion of switchgrass. Biomass Bioen 36:1–11
Sambusiti C, Monlau F, Ficara E, Carrere H, Malpei F (2013) A comparison of different pre-treatments to increase methane production from two agricultural substrates. Appl Energy 104:62–70
Monlau F, Sambusiti C, Barakat A, Quemeneur M, Trably E, Steyer J-P, Carrere H (2014) Do furanic and phenolic compounds of lignocellulosic and algae biomass hydrolyzate inhibit anaerobic mixed cultures?. Biotechnol Adv, A comprehensive review. doi:10.1016/j.biotechadv.2014.04.007
Barakat A, Chuetor S, Monlau F, Solhy A, Rouau X (2013) Ecofriendly dry chemo-mechanical pretreatments of lignocellulosic biomass: impact on energy and yield of the enzymatic hydrolysis. Appl Energy 113:97–105
Sharma SK, Kalra KL, Grewal HS (2002) Enzymatic saccharification of pretreated sunflower stalks. Biomass Bioen 23:237–243
Rabelo SC, Carrere H, Maciel Filho R, Costa AC (2011) Production of bioethanol, methane and heat from sugarcane bagasse in a biorefinery concept. Bioresour Technol 102:7887–7895
Chairattanamanokorn P, Penthamkeerati P, Reungsang A, Lo YC, Lu WB, Chang JS (2009) Production of biohydrogen from hydrolyzed bagasse with thermally preheated sludge. Int J Hydrogen En 34:7612–7617
Monlau F, Trably E, Barakat A, Hamelin J, Steyer J-P, Carrere H (2013) Two-stage alkaline-enzymatic pretreatments to enhance biohydrogen production from sunflower stalks. Environ Sci Technol 47:12591–12599
APHA (2005) Standard Methods for the Examination of Water and Wastewater, 21st edn. American Public Health Association, Washington DC
Effland M (1977) Modified procedure to determine acid-insoluble lignin in wood and pulp. Tappi 60:143–144
Somogyi M (1952) Notes on sugar determination. J Biol Chem 195:19–23
Desrochers M, Jurasek L, Paice MG (1981) High production of & #x03B2;-glucosidase in Schizophyllum commune: isolation of the enzyme and effect of the culture filtrate on cellulose hydrolysis. Appl Environ Microbiol 41:222–228
Shewale JG, Sadana JC (1978) Cellulase and & #x03B2;–glucosidase production by a basidiomycete species. Can J Microbiol 24:1204–1216
OECD (2006) Anaerobic biodegradability of organic compounds in digested sludge by measurement of gas production. Test 311:1–20
Baranyi J, Roberts TA (1994) A dynamic approach to predicting bacterial growth in food. Int J Food Microbiol 23:277–294
Quemeneur M, Bittel M, Trably E, Dumas C, Fourage L, Ravot G, Steyer J-P, Carrere H (2012) Effect of enzyme addition on fermentative hydrogen production from wheat straw. Int J Hydrogen En 37:10639–10647
Li BZ, Balan V, Yuan YJ, Dale BE (2010) Process optimization to convert forage and sweet sorghum bagasse to ethanol based on ammonia fiber expansion (AFEX) pretreatment. Bioresour Technol 101:1285–1292
Manzanares P, Ballesteros I, Negro MJ, Oliva JM, Gonzalez A, Ballesteros M (2012) Biological conversion of forage sorghum biomass to ethanol by steam explosion pretreatment and simultaneous hydrolysis and fermentation at high solid content. Biomass Conv Bioref 2:123–132
Barakat A, Chabbert B, Cathala B (2007) Effect of reaction media concentration on the solubility and the chemical structure of lignin model compounds. Phytochem 68:2118–2125
Gharpuray MM, Lee YH, Fan LT (1983) Structural modification of lignocellulosics by pretreatments to enhance enzymatic hydrolysis. Biotechnol Bioeng 25:157–172
Monlau F, Barakat A, Trably E, Dumas C, Steyer J-P, Carrère H (2012) Lignocellulosic materials into biohydrogen and biomethane: impact of structural features and pretreatment. Crit Rev Environ Sci Technol 43:260–322
Pakarinen OM, Tahti HP, Rintala JA (2009) One-stage H2 and CH4 and two-stage H2 + CH4 production from grass silage and from solid and liquid fractions of NaOH pre-treated grass silage. Biomass Bioen 33:1419–1427
Mc Carty PL (1964) Anaerobic waste treatment fundamentals. Public Works 95:91–99
Acknowledgments
This research study has been conceived and supported by Fabbrica della Bioenergia, which is gratefully acknowledged.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Rollini, M., Sambusiti, C., Musatti, A. et al. Comparative performance of enzymatic and combined alkaline-enzymatic pretreatments on methane production from ensiled sorghum forage. Bioprocess Biosyst Eng 37, 2587–2595 (2014). https://doi.org/10.1007/s00449-014-1235-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00449-014-1235-0