
Abstract
The phylogenetic community assembly approach has been used to elucidate the role of ecological and historical processes in shaping tropical tree communities. Recent studies have shown that stressful environments, such as seasonally dry, white-sand and flooded forests tend to be phylogenetically clustered, arguing for niche conservatism as the main driver for this pattern. Very few studies have attempted to identify the lineages that contribute to such assembly patterns. We aimed to improve our understanding of the assembly of flooded forest tree communities in Northern South America by asking the following questions: are seasonally flooded forests phylogenetically clustered? If so, which angiosperm lineages are over-represented in seasonally flooded forests? To assess our hypotheses, we investigated seasonally flooded and terra firme forests from the Magdalena, Orinoco and Amazon Basins, in Colombia. Our results show that, regardless of the river basin in which they are located, seasonally flooded forests of Northern South America tend to be phylogenetically clustered, which means that the more abundant taxa in these forests are more closely related to each other than expected by chance. Based on our alpha and beta phylodiversity analyses we interpret that eudicots are more likely to adapt to extreme environments such as seasonally flooded forests, which indicates the importance of environmental filtering in the assembly of the Neotropical flora.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
In tropical forests, the phylogenetic community assembly approach has been used to explain the role of ecological and historical processes in shaping these highly diverse tree communities (Eiserhardt et al. 2013; Gerhold et al. 2015; Carlucci et al. 2016). Different historical processes such as time-integrated species-area effect (Fine and Ree 2006; Fine 2015) and extinctions due to major climatic changes affect diversification rates within regional pools and leave imprints in current ecological communities (Parmentier et al. 2007; Kissling et al. 2012). It is possible to hypothesize which historical processes have influenced current community assembly through their effect on the formation of regional pools of species by evaluating whether community phylogenetic structure deviates from random patterns (Webb et al. 2002; Emerson and Gillespie 2008; Cavender-Bares et al. 2009). For instance, a community may be phylogenetically clustered when it displays lower phylogenetic diversity than expected by chance from the species pool, or over-dispersed when it presents higher phylogenetic diversity than expected by chance from the species pool. If adaptations to severe environments involve complex traits, closely related species would be likely to be more similar than expected under neutral evolution, leading to a phylogenetic signal in functional traits linked to such adaptations (Crisp and Cook 2012). In contrast, if adaptation to severe environments involves simple changes that are possible in many unrelated plant lineages, this convergent evolution of traits would result in phylogenetically over-dispersed communities (Emerson and Gillespie 2008).
A number of recent studies have argued that stressful environments, such as seasonally dry, white-sand and seasonally flooded forests tend to be phylogenetically clustered (low phylogenetic diversity) with respect to terra firme forests (Pennington 2009; Gonzalez-Caro et al. 2014; Honorio Coronado et al. 2015; Guevara et al. 2016; but see Fine and Baraloto 2016). Dissimilarities in species composition and richness among South American terra firme and seasonally flooded forests have been attributed to the differences in environmental conditions, mainly soil texture, nutrient content and hydrological stress (Haugaasen and Peres 2006). Recent studies have argued that, in Amazonian rainforests, some closely related tree species have been able to adapt to the relatively stable conditions of floodplains that can be traced as far back to as the Paleocene, and propose phylogenetic niche conservatism (PNC) as a possible explanation for the distribution patterns of the most important species in Amazonian várzea forests (Wittmann et al. 2011, 2013). Responses of tree species to flooding in the Amazon basin include a range of anatomical, morphological and physiological strategies, which can involve xeromorphic leaves, reduction of CO2 uptake, adventitious roots and development of aerenchyma, among other adaptations (Parolin 2008; Parolin and Wittmann 2010).
In the Neotropics, there have been very few studies that have investigated the phylogenetic patterns of seasonally flooded forests. Umaña et al. (2012) reported phylogenetic clustering in Colombian seasonally flooded forests and argued that the differences in species composition and community phylogenetic patterns between terra firme and igapó forests in the Colombian Amazon region are due to habitat specialization to edaphic conditions. Nevertheless, the spatial limitation of the study to a single locality did not allow the authors to make further conclusions about the possible biogeographic causes of the patterns they described. Previous studies have investigated the phylogenetic community structure of Neotropical forests, but only very few have attempted to identify the lineages that are involved in particular assembly patterns (Fine and Kembel 2011; Carlucci et al. 2016). The method used here, principal coordinates of phylogenetic structure (PCPS; Duarte 2011; Duarte et al. 2012) is a phylobetadiversity approach based on ordination that aids in the identification of the major lineages that are better represented in different communities. While PCPS evaluates shifts in phylogenetic composition across communities, the net relatedness index (NRI; Webb et al. 2002), measures local phylogenetic structure of communities (clustering or overdispersion) relative to species pools using null models. Using such an approach, it is possible to relate PCPS scores with NRI values, thereby identifying lineages that are phylogenetically clustered or over-dispersed in different communities.
We aimed to improve knowledge on the assembly of flooded forest communities in Northern South America, by asking the following questions: Are seasonally flooded forests phylogenetically clustered relative to a species pool encompassing both flooded and terra firme forests? If so, which angiosperm lineages are better represented in seasonally flooded forests? To answer these questions, we proposed three hypotheses. First, we tested the null hypothesis of a sampling artifact; seasonally flooded forests may simply have fewer individuals per sampled area, and consequently contain fewer evolutionary lineages (Fine and Kembel 2011). Second, due to environmental filtering, particular lineages may have a higher potential to evolve the traits necessary to survive and reproduce in seasonally flooded forests; thus these forests would have lower phylogenetic diversity than terra firme forests and a clustering of closely related lineages, thereby resembling what has been observed for white-sand and dry forests (Pennington 2009; Fine and Kembel 2011; Guevara et al. 2016). The third hypothesis posits that flood tolerance has independently evolved in many lineages, consistent with convergent evolution, so that seasonally flooded forests have similar phylogenetic structure to terra firme forests and plant lineages in both environments show overdispersion, similar to what was observed when comparing phylogenetic community structure and habitat heterogeneity in the Korup forest in Cameroon (Baldeck et al. 2016).
To test these hypotheses, we studied seasonally flooded (várzea and igapó sensu Prance 1989) and terra firme forests from the Magdalena, Orinoco and Amazon Basins, in Colombia. In this research, we present the most comprehensive attempt to study the phylogenetic community structure of seasonally flooded forests in Northern South America, investigating the potential general importance of environmental filtering as a major factor shaping the evolution of Neotropical floras.
Materials and methods
Study system
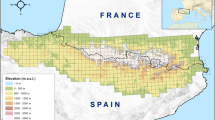
We used information on the abundance of woody species (DBH >10 cm) recorded in 32 1-ha vegetation plots (Stevenson et al. 2004; Aldana et al. 2008; Stevenson and Aldana 2008; Cano and Stevenson 2008; Correa-Gómez and Stevenson 2010; Umaña et al. 2012) from seven lowland forest sites in the Amazon, Magdalena and Orinoco Basins (Fig. 1). These three river basins have undergone radically different geologic processes (Antonelli et al. 2009; Hoorn et al. 2010; Montes et al. 2015). In each site, the plots were classified as terra firme (22), or seasonally flooded (10) if these were submerged for at least 1 month during the rainy season (ESM 1). Vegetation plots in lowland forests sometimes include tree ferns and gymnosperms; in our sites these species represent only an extremely low percentage of the total stems (0.06 %). Taking into account the great effect these basal clades can have in the calculations of phylogenetic metrics, we excluded them from the analyses (Honorio-Coronado et al. 2015).

Approximate location of the 1-ha vegetation plots in various localities of Northern South America used in this study. All sites are located in Colombia. Gray lines represent the boundaries of the three major river basins in the region. Squares represent seasonally flooded forests (2 plots per square) and triangles represent terra firme forests (3–5 plots per triangle). Detailed plot information can be found in Electronic Supplementary Material Table S1. Color version is available online
Phylogenetic tree
To build a phylogenetic tree containing the 1432 species recorded in the study sites, we used the megatree R20120829 (available at https://github.com/camwebb/tree-of-trees/blob/master/megatrees/R20120829.new), which is based on the phylogenetic backbone proposed by APG III (APG 2009) and on relationships among families according to Stevens (2013). The APG III is a consensus tree built from phylogenetic relationships based on several molecular studies that accumulated since the late 1980s, mostly based on analysis of sequences of chloroplast markers and is well resolved with respect to deep phylogenetic relationships (Stevens 2013). The branch lengths of the tree were adjusted through the BLADJ algorithm in Phylocom 4.2 software (Webb et al. 2008) following clade age estimates by (Bell et al. 2010). We used the module Phylomatic in the software Phylocom 4.2 (Webb et al. 2008) to prune the megatree for the species present in our total species pool. The resulting phylogenetic tree (ESM 2) was well resolved for deep nodes and mostly contained polytomies linking species within genera and genera within families. Since we were interested in evaluating major differences in phylogenetic composition related to deep relationships in the phylogeny, the resulting phylogenetic tree was appropriate to test our hypotheses.
Phylogenetic community structure and composition
To test our first hypothesis, a possible sampling artifact due to a lower number of individuals in seasonally flooded forests, we first evaluated the difference in the number of stems between the two categories with a t test. We then applied a rarefaction method (Gotelli and Colwell 2001) to all plots considering 344 individuals, the lowest number of stems registered in a plot, using the function rrarify in the package vegan (Oksanen et al. 2015) for R (R Core Team 2015). Finally, we calculated different phylogenetic metrics for the rarefied communities to compare them to the values of the original plots.
To assess the phylogenetic structure of communities in terms of phylogenetic clustering or overdispersion, we used NRI (Webb et al. 2002). NRI is the standardized effect size of mean phylogenetic distances (MPD) between pairs of co-occurring species (multiplied by −1 to be interpreted in terms of degrees of phylogenetic relatedness). NRI was computed using species abundances and a regional species pool defined as the total species list of all the 32 plots, thereby including both terra firme and seasonally flooded forests. The null model used was phylogeny.pool, which draws species at random with equal probability from the species pool while maintaining plot species richness (Kembel 2009; Kembel et al. 2010). NRI for each community (plot) was also calculated using the basin level species pools, to evaluate the effect of these regional pools in the resulting NRI values.
To assess the phylogenetic composition of lineages across plots, we computed principal coordinates of phylogenetic structure (PCPS, Duarte 2011), which are phylogenetic eigenvectors of an ordination (PCoA) applied to a phylogenetic fuzzy-weighted matrix of community composition (Pillar and Duarte 2010). PCPS vectors describe changes in the phylogenetic composition across plots, and have been used to interpret the dominance of different lineages in different communities (Duarte et al. 2014). NRI and PCPS can be used in tandem to interpret which lineages are related to different degrees of phylogenetic clustering or overdispersion (Pérez-Valera et al. 2015; Carlucci et al. 2016). Studies have shown that NRI and the main vectors of PCPS (first and second) are not strongly affected by phylogenies with low terminal resolution, especially those with a great number of species (Swenson 2009; Maestri et al. 2016), such as ours.
Data analyses
To assess the differences in NRI values between forest types and between river basins we used analysis of variance (ANOVA). We also assessed the difference in the values of the first PCPS axes between forest types using ANOVA. We used a correlation to evaluate the relationship between NRI values of rarefied and sampled communities, and to evaluate the relationship between NRI values and PCPS axis I and II. All analysis were performed in the R environment (R Core Team 2015).
Results
Seasonally flooded forests tend to be less diverse than terra firme forests (Table 1). As expected, seasonally flooded forests in the Amazon basin are richer in species than seasonally flooded forests in the other basins studied; although, comparatively they are less diverse than terra firme forests of the same basin. Even though floristic composition of the várzea and igapó communities included in this study are very different at the family, genus and species levels, the index of importance shows that the most important species of seasonally flooded forests mostly belong to the eudicot clade. The most important species of the terra firme forests belong to the eudicot, as well as the monocot and magnoliid clades (ESM 3).
To test our first hypothesis, the sampling artifact, under which we expected seasonally flooded forests to be phylogenetically clustered due to a lower number of individuals; we compared the number of stems between forests. We found no differences (t = −0.91, p = 0.38) in the number of trees surveyed in flooded and terra firme 1-ha vegetation plots (fl = 504.1, tf = 545.1; Table 1). Additionally, NRI values of rarified communities and NRI values calculated for the original communities were highly correlated (Spearman r = 0.986, p < 0.001), indicating that phylogenetic metrics of seasonally flooded forests are not affected by sample size in this study system.
The calculated NRI values for the original communities of seasonally flooded forests using the total species are significantly higher than terra firme communities and positive, which means that seasonally flooded forests are more phylogenetically clustered and less diverse than terra firme forests in Northern South America (Fig. 2; ESM 4). The majority of terra firme forests had negative NRI values, which means that these communities are more phylogenetically diverse and species tend to be over-dispersed in the phylogeny. When comparing NRI values among river basins we found no significant differences between them (ANOVA, df = 2, F = 0.03, p = 0.971).

Distribution of net relatedness index (NRI) values for flooded and terra firme forests in Northern South America. NRI values of seasonally flooded forests are significantly higher (ANOVA, df = 1, F = 33.95, p < 0.001)
We also examined the effect of the differences in species pools among basins and found that the calculated NRI values for each community are very similar when they are calculated using a phylogenetic tree that only contains the regional species pool from each basin (ESM 1). There was a very strong correlation between NRI calculated with a regional species pool (all 32 plots) and the NRI calculated with basin species pools (0.99, Pearson t = 56.3, p < 0.001).
The PCPS analysis showed that seasonally flooded forests are mostly represented by eudicots, whereas terra firme forests are well represented by magnoliids, monocots and eudicots (Fig. 3; ESM 3). We evaluated the relationship between the first and second axes of the PCPS and the NRI values of the plots and we found a high correlation between PCPS I and NRI (Spearman r = 0.765, p < 0.001) and a low correlation between PCPS II and NRI (Spearman r = −0.321, p > 0.05).

Scatter plot showing the variation in phylogenetic composition across flooded (squares) and terra firme (triangles) rainforest tree communities in Northern South America. Net relatedness index (NRI) values representing clustering or overdispersion are shown in different colors: communities in reddish colors tend to be phylogenetically clustered, while communities in blueish colors tend to be phylogenetically over-dispersed. Phylogenetic composition was assessed using principal coordinates of phylogenetic structure, while phylogenetic structure values (clustering and overdispersion) were computed using the net relatedness index (Webb et al. 2002). Species correlations with PCoA axes located species along PCPS I and II. The spider-like diagrams show the position of species relative to the centroid of their clades in the multivariate space. Note that a PCPS axis shows where clades have more relative importance across the phylogenetic gradient in terms of their relative contribution in abundance and pairwise phylogenetic distances between co-occurring species to communities. Color version is available online
Integrating the PCPS and NRI results, we observed that the tendency toward phylogenetic clustering (higher NRI values) in lowland seasonally flooded forests is explained by the high abundance of the eudicots clade, while the tendency towards phylogenetic overdispersion (lower NRI values) in terra firme forests was associated with a higher relative abundance of magnoliids. This result supports our second hypothesis under which we expected more closely related species to be over-represented due to the stressful environment of floodplains. When comparing PCPS I scores among forest types we found that the phylogenetic composition between flooded and terra firme forests was significantly different (Fig. 4).

Distribution of scores of PCPS I for flooded and terra firme forests in Northern South America. The differences between PCPS scores can be interpreted in terms of phylobetadiversity. Values of seasonally flooded forests are significantly higher (ANOVA, df = 1, F = 23.98, p < 0.001)
Discussion
Our results showed widespread phylogenetic clustering in seasonally flooded forests in Northern South America in relation to a species pool that also contained species from terra firme forests of the region. This pattern was consistent when phylogenetic metrics are calculated for each of the three basins separately. Seasonally flooded forests were phylogenetically clustered regardless of the type of river flooding them (várzea or igapó) or the river basin in which are located (Amazon, Magdalena or Orinoco). Therefore, the most abundant taxa in these seasonally flooded forests are more closely related than expected by chance. Moreover, phylobetadiversity analyses revealed that the dominant taxa in seasonally flooded forests belong to the eudicot clade, mainly from families within the Malpighiales and Fabales orders (ESM 3).
Species diversity in local seasonally flooded communities is low in comparison to terra firme forests, with a mean of 60.5 species per plot (Table 1), which is expected to yield high NRI values because it has been previously reported that low species diversity correlates with low phylogenetic diversity (ESM 4) (Honorio-Coronado et al. 2015). Additionally, total species diversity of all the seasonally flooded forests plots (10) is 443, which is significantly lower than the 1212 total species for all terra firme plots (22).
Our results indicate a role for habitat filtering (Wiens and Graham 2005; Losos 2008; Crisp and Cook 2012) in flooded forest community assembly. It seems that despite the great diversity of lineages that can be found in the study forests, few magnoliids have evolved traits necessary to survive the environmental pressure of flooding. Wittmann et al. (2013) found similar floristic composition within seasonally flooded forests from the Amazon and Orinoco regions, and argued that the prolonged stable conditions of flooded environments since the Early Miocene (Hoorn et al. 2010) in Northern South America has allowed for taxa to specialize and adapt to these habitats (Wittmann et al. 2013). We agree with this interpretation and suggest that specialization to different types of substrates and nutrient contents has occurred since the uplift of the Andes, mainly during the late Miocene when the drainage systems in the region radically changed (Mora et al. 2011). As expected, species diversity of seasonally flooded forests in the Amazon basin is higher than in the Orinoco and Magdalena basins. This pattern could be explained by the time-integrated species-area effect (Fine and Ree 2006) where taxa have had longer to diversify in prolonged flooding conditions in the Amazon basin (Hoorn et al. 2010) and less time to diversify in the river systems that resulted from the Andean orogeny (Antonelli et al. 2009).
We assessed alpha and phylobetadiversity in tropical forest communities using an integrated framework that permitted us to evaluate which angiosperm lineages contributed the most to local phylogenetic clustering or overdispersion patterns. Examining the relationship between NRI values and PCPS scores is a valuable way to integrate alpha phylogenetic structure and phylobetadiversity (Carlucci et al. 2016). Previous studies have used the tool nodesig (Webb et al. 2008) to identify lineages over-represented in sets of communities, e.g., delimited by a habitat type (Fine and Kembel 2011), an objective similar to ours in the present study. However, in nodesig information on non-random patterns of taxa abundance in communities is interpreted with respect to the phylogeny; whereas in PCPS phylogenetic information is interpreted at the community scale because an ordination score of phylogenetic weighted community composition is attributed to each community. Consequently, although PCPS and nodesig may be used to answer similar questions, PCPS enables identifying lineages that are more common in sets of communities while assessing phylobetadiversity across communities. Moreover, PCPS enables a continuous evaluation across communities, with no need to categorize communities into different habitat types.
While investigating the traits necessary for trees to adapt to seasonal flooding in the Amazon, Parolin (2008) found that the main strategy used by many species was that of general flood tolerance, as opposed to survival or escape, which is accomplished by a great diversity of different metabolic and morphological adaptations. Similar to our results, Pizano and García (2014) found the composition of Neotropical dry forests included a high dominance of eudicots (Pizano and García 2014). We thus postulate that adaptations to extreme environments could be important in promoting eudicot dominance in dry and seasonally flooded forests. Field and experimental studies assessing metabolic and morphological adaptations of plant species to life in flooded and dry forests can be used in the future to test this idea.
We conclude that seasonally flooded forests of Northern South America have low taxonomic and phylogenetic diversity compared to surrounding terra firme forests due to the effect of historical environmental filtering leading to adaptive radiations within lineages of eudicots. Our results suggest that eudicots possess some advantage in relation to magnoliids that has allowed them to evolve the necessary traits to adapt to extreme environments such as seasonal floodplains. We propose that further research should focus on investigating the traits that confer flood tolerance to plants of this important clade of angiosperms.
References
Aldana AM, Beltrán M, Torres-Neira J, Stevenson PR (2008) Habitat Characterization and population density of brown spider monkeys (Ateles hybridus) in Magdalena Valley, Colombia. Neotrop Primates 15:46–50. doi:10.1896/044.015.0203
Antonelli A, Nylander JA, Persson C, Sanmartín I (2009) Tracing the impact of the Andean uplift on Neotropical plant evolution. Proc Natl Acad Sci 106:9749–9754
APG (2009) An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III. Bot J Linn Soc 161:105–121
Baldeck CA, Kembel SW, Harms KE et al (2016) Phylogenetic turnover along local environmental gradients in tropical forest communities. Oecologia. doi:10.1007/s00442-016-3686-2
Bell CD, Soltis DE, Soltis PS (2010) The age and diversification of the angiosperms re-revisited. Am J Bot 97:1296–1303. doi:10.3732/ajb.0900346
Cano A, Stevenson PR (2008) Diversidad y composición florística de tres tipos de bosque en la Estación Biológica Caparú, Vaupés. Colomb For 12:63. doi:10.14483/udistrital.jour.colomb.for.2009.1.a06
Carlucci MB, Seger GDS, Sheil D et al (2016) Phylogenetic composition and structure of tree communities shed light on historical processes influencing tropical rainforest diversity. Ecography (Cop). doi:10.1111/ecog.02104
Cavender-Bares J, Kozak KH, Fine PVA, Kembel SW (2009) The merging of community ecology and phylogenetic biology. Ecol Lett 12:693–715. doi:10.1111/j.1461-0248.2009.01314.x
Correa-Gómez DF, Stevenson PR (2010) Estructura y diversidad de bosques de los llanos orientales colombianos (Reserva Tomo Grande, Vichada). Revista Orinoquia 14:31–48
Crisp MD, Cook LG (2012) Phylogenetic niche conservatism: what are the underlying evolutionary and ecological causes? New Phytol 196:681–694
Duarte LDS (2011) Phylogenetic habitat filtering influences forest nucleation in grasslands. Oikos 120:208–215. doi:10.1111/j.1600-0706.2010.18898.x
Duarte LDS, Prieto PV, Pillar VD (2012) Assessing spatial and environmental drivers of phylogenetic structure in Brazilian Araucaria forests. Ecography (Cop) 35:952–960. doi:10.1111/j.1600-0587.2011.07193.x
Duarte LDS, Both C, Debastiani VJ et al (2014) Climate effects on amphibian distributions depend on phylogenetic resolution and the biogeographical history of taxa. Glob Ecol Biogeogr 23:213–222. doi:10.1111/geb.12089
Eiserhardt WL, Svenning J-C, Baker WJ et al (2013) Dispersal and niche evolution jointly shape the geographic turnover of phylogenetic clades across continents. Sci Rep 3:1164. doi:10.1038/srep01164
Emerson BC, Gillespie RG (2008) Phylogenetic analysis of community assembly and structure over space and time. Trends Ecol Evol 23:619–630. doi:10.1016/j.tree.2008.07.005
Fine P (2015) Ecological and evolutionary drivers of geographic variation in species diversity. Annu Rev Ecol Evol Syst 46:369–392. doi:10.1146/annurev-ecolsys-112414-054102
Fine PVA, Baraloto C (2016) Habitat endemism in white-sand forests: insights into the mechanisms of lineage diversification and community assembly of the Neotropical flora. Biotropica 48:24–33. doi:10.1111/btp.12301
Fine PVA, Kembel SW (2011) Phylogenetic community structure and phylogenetic turnover across space and edaphic gradients in western Amazonian tree communities. Ecography (Cop) 34:552–565. doi:10.1111/j.1600-0587.2010.06548.x
Fine PVA, Ree RH (2006) Evidence for a time-integrated species-area effect on the latitudinal gradient in tree diversity. Am Nat 168:796–804. doi:10.1086/508635
Gerhold P, Cahill JF, Winter M et al (2015) Phylogenetic patterns are not proxies of community assembly mechanisms (they are far better). Funct Ecol 29:600–614. doi:10.1111/1365-2435.12425
Gonzalez-Caro S, Umana MN, Alvarez E et al (2014) Phylogenetic alpha and beta diversity in tropical tree assemblages along regional-scale environmental gradients in northwest South America. J Plant Ecol 7:145–153. doi:10.1093/jpe/rtt076
Gotelli NJ, Colwell RK (2001) Quantifying biodiversity: procedures and pitfalls in the measurement and comparison of species richness. Ecol Lett 4:379–391
Guevara JE, Damasco G, Baraloto C et al (2016) Low phylogenetic beta diversity and geographic neo-endemism in Amazonian white-sand forests. Biotropica 48:34–46. doi:10.1111/btp.12298
Haugaasen T, Peres CA (2006) Floristic, edaphic and structural characteristics of flooded and unflooded forests in the lower Rio Purús region of central Amazonia, Brazil. Acta Amaz 36:25–36
Honorio-Coronado EN, Dexter KG, Pennington RT et al (2015) Phylogenetic diversity of Amazonian tree communities. Divers Distrib 21:1295–1307. doi:10.1111/ddi.12357
Hoorn C, Roddaz M, Dino R et al (2010) The Amazonian craton and its influence on past fluvial systems (Mesozoic-Cenozoic, Amazonia). In: Hoorn C, Wesselingh FP (eds) Amazonia: landscape and species evolution. Wiley-Blackwell Publishing Ltd., Oxford, pp 101–122
Kembel SW (2009) Disentangling niche and neutral influences on community assembly: assessing the performance of community phylogenetic structure tests. Ecol Lett 12:949–960. doi:10.1111/j.1461-0248.2009.01354.x
Kembel SW, Cowan PD, Helmus MR et al (2010) Picante: r tools for integrating phylogenies and ecology. Bioinformatics 26:1463–1464. doi:10.1093/bioinformatics/btq166
Kissling WD, Eiserhardt WL, Baker WJ et al (2012) Cenozoic imprints on the phylogenetic structure of palm species assemblages worldwide. Proc Natl Acad Sci 109:7379–7384. doi:10.1073/pnas.1120467109
Losos J (2008) Phylogenetic niche conservatism, phylogenetic signal and the relationship between phylogenetic relatedness and ecological similarity among species. Ecol Lett. doi:10.1111/j.1461-0248.2008.01229.x
Maestri R, Luza AL, de Barros LD, Hartz SM, Ferrari A, de Freitas TRO, Duarte LDS (2016) Geographical variation of body size in sigmodontine rodents depends on both environment and phylogenetic composition of communities. J Biogeogr 43:1192–1202. doi:10.1111/jbi.12718
Montes C, Cardona A, Jaramillo C et al (2015) Middle Miocene closure of the Central American seaway. Science 348:226–229. doi:10.1126/science.aaa2815 (80-)
Mora A, Baby P, Roddaz M et al (2011) Tectonic history of the Andes and sub-andean zones: implications for the development of the Amazon drainage basin. In: Hoorn C, Wesselingh FP (eds) Amazonia: landscape and species evolution. Wiley-Blackwell Publishing Ltd., Oxford, pp 38–60
Oksanen J, Guillaume Blanchet F, Kindt R et al (2015) vegan: community ecology package. R package version 2.3-0
Parmentier I, Malhi Y, Senterre B et al (2007) The odd man out? Might climate explain the lower tree α-diversity of African rain forests relative to Amazonian rain forests? J Ecol 95:1058–1071. doi:10.1111/j.1365-2745.2007.01273.x
Parolin P (2008) Submerged in darkness: adaptations to prolonged submergence by woody species of the Amazonian floodplains. Ann Bot 103:359–376. doi:10.1093/aob/mcn216
Parolin P, Wittmann F (2010) Struggle in the flood: tree responses to flooding stress in four tropical floodplain systems. AoB Plants 2010:plq003–plq003. doi:10.1093/aobpla/plq003
Pennington R (2009) Woody plant diversity, evolution, and ecology in the tropics: perspectives from seasonally dry tropical forests. Annu Rev. doi:10.1146/annurev.ecolsys.110308.120327
Pérez-Valera E, Goberna M, Verdú M (2015) Phylogenetic structure of soil bacterial communities predicts ecosystem functioning. FEMS Microbiol Ecol 91:fiv031. doi:10.1093/femsec/fiv031
Pillar VD, Duarte LDS (2010) A framework for metacommunity analysis of phylogenetic structure. Ecol Lett 13:587–596. doi:10.1111/j.1461-0248.2010.01456.x
Pizano C, García H (eds) (2014) El Bosque Seco Tropical en Colombia. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt, Bogotá
Prance G (1989) American tropical forests. Trop Rain For Ecosyst Ecol Stud. doi:10.1016/B978-0-444-42755-7.50012-2
R Core Team (2015) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Stevens PF (2013) Angiosperm Phylogeny Website, version 13. http://www.mobot.org/MOBOT/research/APweb/. Accessed 2 Aug 2016
Stevenson PR, Aldana AM (2008) Potential effects of ateline extinction and forest fragmentation on plant diversity and composition in the Western Orinoco Basin, Colombia. Int J Primatol 29:365–377. doi:10.1007/s10764-007-9177-x
Stevenson PR, Suescún M, Quiñones M (2004) Characterization of forest types at the CIEM, Tinigua Park, Colombia. F Stud Fauna Flora La Macarena Colomb 14:1–20
Swenson NG (2009) Phylogenetic resolution and quantifying the phylogenetic diversity and dispersion of communities. PLoS One 4:e4390. doi:10.1371/journal.pone.0004390
Umaña MN, Norden N, Cano A, Stevenson PR (2012) Determinants of plant community assembly in a mosaic of landscape units in central Amazonia: ecological and phylogenetic perspectives. PLoS One 7:e45199. doi:10.1371/journal.pone.0045199
Webb CO, Ackerly DD, McPeek MA, Donoghue MJ (2002) Phylogenies and community ecology. Annu Rev Ecol Syst 33:475–505. doi:10.1146/annurev.ecolsys.33.010802.150448
Webb CO, Ackerly DD, Kembel SW (2008) Phylocom: software for the analysis of phylogenetic community structure and character evolution. Bioinformatics 24:2098–2100
Wiens JJ, Graham CH (2005) Niche conservatism: integrating evolution, ecology, and conservation biology. Annu Rev Ecol Evol Syst 36:519–539. doi:10.1146/annurev.ecolsys.36.102803.095431
Wittmann F, Schöngart J, Junk WJ (2011) Amazonian floodplain forests. Springer, Dordrecht
Wittmann F, Householder E, Piedade MTF et al (2013) Habitat specificity, endemism and the neotropical distribution of Amazonian white-water floodplain trees. Ecography (Cop) 36:690–707. doi:10.1111/j.1600-0587.2012.07723.x
Acknowledgments
We thank Ángela Cano, Sasha Cárdenas, Luisa Fernanda Casas, Diego Felipe Correa, Mabel Suescún, María Natalia Umaña, Boris Villanueva for the information provided from the plots they established. AMA would like to thank James Richardson and Toby Pennington for helpful discussions on the results reported here and the Universidad de Los Andes for providing funding to visit the University of California, Berkeley where this research was envisioned. MBC received fellowships from CAPES-Brazil (Grants BEX7913/13-3 and PNPD #1454013). We are thankful to Dr. Ethan Householder and two anonymous reviewers who helped improve an earlier version of this manuscript.
Author contribution statement
PRS and AMA formulated the idea. AMA, MBC and PVAF developed the methodology. AMA and MBC wrote the manuscript; other authors provided advice and edited the manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Katherine L. Gross.
Evaluation of the phylogenetic diversity of forest communities in the Neotropics is needed to understand the evolutionary process that have led to high species diversity in these forests. We explored the phylogenetic patterns of flooded and terra firme forests in Northern South America and found that flooded forests are low in species diversity as well as in phylogenetic diversity, due to the fact that only a few lineages of the eudicot clade are able to succeed in these extreme conditions. As such, this research helps elucidate a fragment of the evolutionary history of Neotropical forests, and should stimulate tests of new hypotheses.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Aldana, A.M., Carlucci, M.B., Fine, P.V.A. et al. Environmental filtering of eudicot lineages underlies phylogenetic clustering in tropical South American flooded forests. Oecologia 183, 327–335 (2017). https://doi.org/10.1007/s00442-016-3734-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00442-016-3734-y