
Abstract
Microbialites, bioaccretionary structures formed during the growth and metabolism of microorganisms (principally cyanobacteria) were the dominant lifeform in shallow late-Archean and Proterozoic oceans. During the Cambrian radiation of metazoan life, which began ~540 Mya, microbialite abundance and diversity further declined following a peak in the Mesoproterozoic. Notwithstanding contention, grazing and bioturbation effects of metazoans have been hypothesized as the dominant driver of modern microbialite scarcity. However, this metazoan–microbialite exclusion has not been fully explored in the few extant microbialites. Here we provide further evidence showing that living marine layered microbialites (stromatolites) coexist with a persistent assemblage of benthic macro-invertebrates, as has previously been demonstrated in some thrombolitic (clotted) microbialites. Surprisingly, these metazoans have active habits, such as burrowing, which should be expected to disrupt the layered matrix. As other studies have shown, through a network of burrows, metazoans can exploit local diurnal oxygen refugia within microbialites as well as escape predation. Our results, therefore, add novel evidence in support of the hypotheses that geologically, metazoans are not always incompatible with stromatolites, while ecologically, microbialites may act as micro-refugia for modern metazoans and historically have performed a similar inferred role in past ecosystems.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
As the dominant lifeform in pre-Cambrian oceans (Riding 2000, 2006) and as a consequence of their ability to trap sediment in excreted extrapolymeric substances (Frantz et al. 2015; Reid et al. 2000) and precipitate calcium carbonate (Riding 2012) during growth and metabolism, many benthic cyanobacterial mats have been preserved in extensive layered microbialite (stromatolite) deposits from 0.5 to 3.5 Bya (Grotzinger and Knoll 1999; Riding 2000). Their important role in the Earth’s history is exemplified by the increased levels of atmospheric oxygen realized in the Phanerozoic, which is attributed to the evolution of oxygenic photosynthesis in cyanobacteria (Dismukes et al. 2001). However, aerobic respiration, which contributed towards enabling the evolution and radiation of the metazoans during the Cambrian (Marshall 2006), corresponded with a further decline into the Neoproterozoic of stromatolite abundance and diversity (Riding 2000, 2006), which has not recovered since.
While the factors driving historical stromatolite decline have been debated (Riding 2000), both geological evidence (Mata and Bottjer 2012; Walter and Heys 1985) and comparisons with modern microbialite analogues (Feldmann and McKenzie 1998; Garrett 1970) suggest that metazoans have a negative effect on stromatolite formation through grazing and burrowing. This has also been experimentally demonstrated, whereby estuarine cyanobacterial benthic mats form stromatolitic layering if metazoans are excluded but revert to a homogenized, non-layered matrix when metazoans are present (Fenchel 1998). Consequently, most modern stromatolites are restricted to marginal, harsh environments where metazoans are largely excluded (Grotzinger and Knoll 1999; Riding 2011), whereas in the presence of metazoans, thrombolites (clotted microbialites) rather than laminated stromatolites may form (Feldmann and McKenzie 1998; Konishi et al. 2001; Planavsky and Ginsburg 2009; Walter and Heys 1985). Recently-discovered peritidal stromatolites along the southern African coastline (Perissinotto et al. 2014; Rishworth et al. 2016; Smith et al. 2011) were used in this study to further assess metazoan coexistence and lamination in comparison to other modern stromatolites (Edgcomb et al. 2014; Garcia-Pichel et al. 2004; Tarhan et al. 2013) in light of the ecologically and evolutionarily relevant metazoan–stromatolite exclusion hypothesis (Garrett 1970; Mata and Bottjer 2012; Riding 2000; Walter and Heys 1985).
Materials and methods
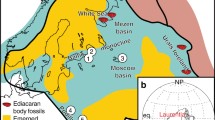
Three sites were selected along the coastline near Port Elizabeth, South Africa (Fig. 1): Cape Recife (34°02′42.13″S, 25°34′07.50″E), Schoenmakerskop (34°02′28.23″S, 25°32′18.60″E), and Seaview (34°01′03.16″S, 25°21′56.48″E) (Perissinotto et al. 2014; Rishworth et al. 2016). Each site is characterized by a freshwater inlet stream which flows into a ‘barrage’ tufa stromatolite pool or series of pools in the upper intertidal zone. Dominant mat-building microalgae include diatoms and cyanobacteria (Rishworth et al. 2016), specifically genera such as Lyngbya, Plectonema, Leptolyngbya and Schizothrix (Perissinotto et al. 2014). The stromatolites accrete at an optimal rate of 2.1–3.6 mm year−1, forming sequential primary and successional layers (Fig. 2a; Smith et al. 2005).

From: Perissinotto et al. (2014)
Three sampling locations (A Cape Recife, B Schoenmakerskop, and C Seaview) along the South African coastline, with all 540 stromatolite sites recorded thus far indicated by black dots

Microstructure and metazoan activities within living marine stromatolites from the Eastern Cape, South Africa. a Scanning electron microscope (SEM) micrograph of an actively growing stromatolite indicating the climax layer with an arrow (Smith et al. 2011) between lithified cyanobacterial filaments. Scale 500 μm. b Lithified, spent (non-growing) stromatolite section. Scale 1 cm. c Burrow network formed by metazoans within a growing stromatolite, indicated by arrows, dorsal view. Scale 1 mm. d Examples of Naididae oligochaetes (Clitellata) fixed ventrally within the stromatolite matrix, as indicated by arrows, demonstrating the association of metazoans with microbialite lamination. Scale 1 mm
Macrofaunal samples were collected using a stainless steel corer with an internal diameter of 1.7 cm. At each site, three samples were extracted from the upper 2 cm of the partially lithified stromatolite matrix from the main stromatolite barrage pool as well as from the lower elevation, seawards pool and upper elevation, freshwater inlet pool. Core samples ensured that all macrofauna directly associated with the stromatolite matrix were collected (Tarhan et al. 2013). All samples were preserved in 5 % formaldehyde buffered with seawater. The stromatolite matrix was carefully broken up on a 500 μm sieve, thereby extracting all macrofauna visually after staining with Phloxin-B. All specimens were visually identified to the lowest taxonomic level possible, using available local and international reference guides as well as in consultation with relevant taxonomic experts. Samples were collected monthly from each of the three pools at each site during spring low tide from January to July 2014.
In addition, physico-chemical properties of the water in the lower, barrage, and upper stromatolite pools were recorded using a YSI 6600 multi-parameter meter (YSI Incorporated, Yellow Springs, OH, USA), the details of which are presented elsewhere (Rishworth et al. 2016). These included temperature, salinity, and dissolved oxygen (mg L−1).
Results and discussion
Tufa stromatolites occurring in South Africa form at the interface of groundwater seepage and the ocean high-water mark (Perissinotto et al. 2014; Rishworth et al. 2016), constructing rimstone dams through the layered precipitation of inflowing calcium carbonate and the trapping of sediment (Smith et al. 2011, 2005). Within the clearly layered, growing stromatolite matrix (Fig. 2a), which has been preserved at the study sites in areas where freshwater inflow has ceased or the high-water mark shifted (Fig. 2b), a persistent assemblage of benthic invertebrates (metazoans) was encountered (Table 1). This uncommon microbialite–metazoan association has been documented in other ecosystems where, although in lower diversity compared to microbes (Allen et al. 2009), examples of extant microbialites forming in the spectrum from fresh to hypersaline waters have been shown to support metazoan infauna (Table 2). Counterintuitively, the co-occurring metazoans in these clearly layered systems are not passive inhabitants of stromatolites but rather have active habits, such as grazing and burrowing (Table 1), which might be expected to disrupt, homogenize and bioturbate (Fenchel 1998; Garrett 1970; Pike et al. 2001; Smith et al. 2000) the microalgal mats, rather than form a layered substrate. Following the evolution of metazoans, this is indeed what disrupted and reduced the microbial mats which dominated pre-Cambrian oceans (Buatois et al. 2014). However, active burrowing, specifically (Fig. 2c), is apparently not restrictive to stromatolite formation and layering in the presented example, with burrowing metazoans, such as oligochaetes and nereid polychaetes, regularly observed amongst the layered substrate (Fig. 2d). Grazing has been inferred from known invertebrate behaviors (Table 1), with trophic linkages and feeding ecology yet to be determined. In our study, metazoans do not appear to be bioturbating the stromatolite into its clotted (thrombolite) form, despite in situ observations of sediment-containing faecal pellets and debris adjacent to burrows. These results suggest that rather than being formed by metazoan activities which ‘remodel’ a layered matrix (Planavsky and Ginsburg 2009; Walter and Heys 1985), thrombolites are likely constructed by distinct microbial communities with different deposition- and sediment-trapping mechanisms (i.e. layering versus non-layering) compared to those which form stromatolites (Myshrall et al. 2010). Nonetheless, both fossil evidence and modern comparisons suggest a higher prevalence of thrombolite–metazoan associations over stromatolite examples (Konishi et al. 2001; Walter and Heys 1985). Thus far, all mesofabric types encountered at the South African sites are layered (stromatolitic), with another complementary study currently underway which would reveal any thrombolite material if present.
The stromatolite–metazoan relationship extends beyond the potential disruptive effect of grazers and burrowers. Higher oxygen concentrations within the microbialite-forming photosynthetic microalgal mats (Des Marais 2003; Herman and Kump 2005) might have provided a micro-refuge for early metazoans to escape the unfavourable, anoxic ambient conditions of the Ediacaran (Tarhan et al. 2013). This is argued as a factor which drove the evolution of metazoan mobility, allowing animals to occupy the anoxic, food-rich benthos while moving between and extracting the metabolically beneficial oxygen being formed within the microbialites (Gingras et al. 2011). However, this hypothesis is confounded by the anoxic conditions which would develop at night (Des Marais 2003; Mobberley et al. 2015) as well as the range of metabolic modes of microbialites, in addition to photoautotrophy (Ruvindy et al. 2016). Nonetheless, similar micro-refugia would be present within some modern stromatolites that support metazoans (Table 2), although not strictly assessed in terms of oxygen concentration gradients in the current study, these being accessed through a network of burrows within the matrix (Fig. 2c). The micro-habitat characteristics of extant microbialites may, thereby, be providing localized respite for invertebrates from predation (Dinger et al. 2006; Konishi et al. 2001) and harsh physical conditions, as well as offering an oxygen refuge (Des Marais 2003; Gingras et al. 2011; Herman and Kump 2005; Mobberley et al. 2015). For example, in the South African stromatolite pools during the warmer months, anoxic events have been observed whereby a milky benthic layer with a distinctive hydrogen sulphide odor forms [oxygen concentrations as low as 0.24 mg L−1, compared to supersaturated levels of up to 37.0 mg L−1 above the stromatolites (Perissinotto et al. 2014; Rishworth et al. 2016)], from which metazoans might use the adjacent stromatolite matrix as a temporary oxygen refuge during daylight hours. It is known from a variety of other extant microbialites that oxygen concentrations are elevated within the microalgal layer, compared to the surrounding media (Des Marais 2003; Gingras et al. 2011; Herman and Kump 2005; Mobberley et al. 2015). Furthermore, on a geographical scale, the South African stromatolites appear to be functioning as unique ecological niches, supporting noteworthy species such as Tuoba cf. poseidonis, the first record of a halophilic centipede for the southern African region (Barber 2011), and Bolttsia minuta, an estuarine-relict amphipod known originally only from a land-locked coastal lake (Griffiths 1976). The microbial community for some microbialites appear distinctive from adjacent waters (White III et al. 2016), however, it is unclear whether this holds true for any of the metazoans encountered in this analysis, of which the majority appear to be typical estuarine residents (Table 1).
In conclusion, the developing recognition, to which the current study adds further evidence, that stromatolites and metazoans are not always mutually exclusive, neither historically (Riding 2006) nor currently (Table 2), is important from an evolutionary perspective. Metazoans certainly contributed towards the decline of the microbialites (Buatois et al. 2014; Mata and Bottjer 2012), particularly the stromatolites, and their exclusion in modern habitats has enabled some microbialites to thrive (Eckman et al. 2008). However, the surprising coexistence of clearly layered stromatolites with active burrowing and grazing metazoans (Fig. 2d; Table 1) highlights the complexity of this relationship (Garcia-Pichel et al. 2004; Tarhan et al. 2013), suggesting that other environmental factors, such as changing water chemistry (Grotzinger 1990) or nutrient limitation (Elser et al. 2006), might indeed have contributed to the scarcity of the stromatolites by the Holocene. Microbial mats seem to have offered one of the essential catalysts to enable the evolution of animals by providing accessible oxygen micro-reservoirs in the largely anoxic Ediacaran oceans (Gingras et al. 2011). Therefore, the observed, albeit rare, coexistence of these two biological groups might largely be a consequence of the benefit derived by metazoan infauna in terms of oxygen resources and predator avoidance, which would select against the destructive effect of metazoans on their micro-refugia.
References
Allen MA, Goh F, Burns BP, Neilan BA (2009) Bacterial, archaeal and eukaryotic diversity of smooth and pustular microbial mat communities in the hypersaline lagoon of Shark Bay. Geobiology 7:82–96. doi:10.1111/j.1472-4669.2008.00187.x
Barber A (2011) Geophilomorph centipedes and the littoral habitat. Terr Arthropod Rev 4:17–39. doi:10.1163/187498311x546986
Buatois LA, Narbonne GM, Mangano MG, Carmona NB, Myrow P (2014) Ediacaran matground ecology persisted into the earliest Cambrian. Nat Commun 5:3544. doi:10.1038/ncomms4544
Des Marais DJ (2003) Biogeochemistry of hypersaline microbial mats illustrates the dynamics of modern microbial ecosystems and the early evolution of the biosphere. Biol Bull 204:160–167. doi:10.2307/1543552
Dinger EC, Hendrickson DA, Winsborough BM, Marks JC (2006) Role of fish in structuring invertebrates on stromatolites in Cuatro Ciénegas, México. Hydrobiologia 563:407–420. doi:10.1007/s10750-006-0040-4
Dismukes GC, Klimov VV, Baranov SV, Kozlov YN, DasGupta J, Tyryshkin A (2001) The origin of atmospheric oxygen on Earth: the innovation of oxygenic photosynthesis. Proc Natl Acad Sci 98:2170–2175. doi:10.1073/pnas.061514798
Eckman JE et al (2008) Wave and sediment dynamics along a shallow subtidal sandy beach inhabited by modern stromatolites. Geobiology 6:21–32. doi:10.1111/j.1472-4669.2007.00133.x
Edgcomb VP, Bernhard JM, Summons RE, Orsi W, Beaudoin D, Visscher PT (2014) Active eukaryotes in microbialites from Highborne Cay, Bahamas, and Hamelin Pool (Shark Bay) Australia. ISME J 8:418–429. doi:10.1038/ismej.2013.130
Elser JJ, Watts J, Schampel JH, Farmer J (2006) Early Cambrian food webs on a trophic knife-edge? A hypothesis and preliminary data from a modern stromatolite-based ecosystem. Ecol Lett 9:295–303. doi:10.1111/j.1461-0248.2005.00873.x
Feldmann M, McKenzie JA (1998) Stromatolite-thrombolite associations in a modern environment, Lee Stocking Island, Bahamas. Palaios 13:201–212. doi:10.2307/3515490
Fenchel T (1998) Formation of laminated cyanobacterial mats in the absence of benthic fauna. Aquat Microb Ecol 14:235–240
Frantz CM, Petryshyn VA, Corsetti FA (2015) Grain trapping by filamentous cyanobacterial and algal mats: implications for stromatolite microfabrics through time. Geobiology 13:409–423. doi:10.1111/gbi.12145
Garcia-Pichel F, Al-Horani FA, Farmer JD, Ludwig R, Wade BD (2004) Balance between microbial calcification and metazoan bioerosion in modern stromatolitic oncolites. Geobiology 2:49–57. doi:10.1111/j.1472-4669.2004.00017.x
Garrett P (1970) Phanerozoic stromatolites: noncompetitive ecologic restriction by grazing and burrowing animals. Science 169:171–173. doi:10.2307/1730310
Gingras M et al (2011) Possible evolution of mobile animals in association with microbial mats. Nat Geosci 4:372–375. doi:10.1038/ngeo1142
Griffiths CL (1976) Some new and notable Amphipoda from southern Africa. S Afr Mus 72:11–35
Grotzinger JP (1990) Geochemical model for Proterozoic stromatolite decline. Am J Sci 290:80–103
Grotzinger JP, Knoll AH (1999) Stromatolites in Precambrian carbonates: evolutionary mileposts or environmental dipsticks? Annu Rev Earth Planet Sci 27:313–358. doi:10.1146/annurev.earth.27.1.313
Herman EK, Kump LR (2005) Biogeochemistry of microbial mats under Precambrian environmental conditions: a modelling study. Geobiology 3:77–92. doi:10.1111/j.1472-4669.2005.00048.x
Konishi Y, Prince J, Knott B (2001) The fauna of thrombolitic microbialites, Lake Clifton, western Australia. Hydrobiologia 457:39–47. doi:10.1023/a:1012229412462
Marshall CR (2006) Explaining the Cambrian “explosion” of animals. Annu Rev Earth Planet Sci 34:355–384. doi:10.1146/annurev.earth.33.031504.103001
Mata SA, Bottjer DJ (2012) Microbes and mass extinctions: paleoenvironmental distribution of microbialites during times of biotic crisis. Geobiology 10:3–24. doi:10.1111/j.1472-4669.2011.00305.x
Mobberley JM, Khodadad CL, Visscher PT, Reid RP, Hagan P, Foster JS (2015) Inner workings of thrombolites: spatial gradients of metabolic activity as revealed by metatranscriptome profiling. Sci Rep 5:12601. doi:10.1038/srep12601
Myshrall KL et al (2010) Biogeochemical cycling and microbial diversity in the thrombolitic microbialites of Highborne Cay, Bahamas. Geobiology 8:337–354. doi:10.1111/j.1472-4669.2010.00245.x
Perissinotto R et al (2014) Tufa stromatolite ecosystems on the South African south coast. S Afr J Sci 110:89–96. doi:10.1590/sajs.2014/20140011
Pike J, Bernhard JM, Moreton SG, Butler IB (2001) Microbioirrigation of marine sediments in dysoxic environments: implications for early sediment fabric formation and diagenetic processes. Geology 29:923–926. doi:10.1130/0091-7613(2001)029<0923:momsid>2.0.co;2
Planavsky N, Ginsburg RN (2009) Taphonomy of modern marine Bahamian microbialites. Palaios 24:5–17. doi:10.2110/palo.2008.p08-001r
Reid RP et al (2000) The role of microbes in accretion, lamination and early lithification of modern marine stromatolites. Nature 406:989–992. doi:10.1038/35023158
Riding R (2000) Microbial carbonates: the geological record of calcified bacterial–algal mats and biofilms. Sedimentology 47:179–214. doi:10.1046/j.1365-3091.2000.00003.x
Riding R (2006) Microbial carbonate abundance compared with fluctuations in metazoan diversity over geological time. Sediment Geol 185:229–238. doi:10.1016/j.sedgeo.2005.12.015
Riding R (2011) The nature of stromatolites: 3500 million years of history and a century of research. In: Reitner J, Quéric NV, Arp G (eds) Advances in stromatolite geobiology, vol 131. Springer, Berlin Heidelberg, pp 29–74
Riding R (2012) A hard life for cyanobacteria. Science 336:427–428. doi:10.1126/science.1221055
Rishworth GM, van Elden S, Perissinotto R, Miranda NAF, Steyn PP, Bornman TG (2016) Environmental influences on living marine stromatolites: insights from benthic microalgal communities. Environ Microbiol 18:503–513. doi:10.1111/1462-2920.13116
Ruvindy R, White RA III, Neilan BA, Burns BP (2016) Unravelling core microbial metabolisms in the hypersaline microbial mats of Shark Bay using high-throughput metagenomics. ISME J 10:183–196. doi:10.1038/ismej.2015.87
Smith CR, Levin LA, Hoover DJ, McMurtry G, Gage JD (2000) Variations in bioturbation across the oxygen minimum zone in the northwest Arabian Sea. Deep Sea Res Part II Top Stud Oceanogr 47:227–257. doi:10.1016/S0967-0645(99)00108-3
Smith AM, Uken R, Thackeray Z (2005) Cape Morgan peritidal stromatolites: the origin of lamination. S Afr J Sci 101:107–108
Smith AM et al (2011) Rock pool tufa stromatolites on a modern South African wave-cut platform: partial analogues for Archaean stromatolites? Terra Nova 23:375–381. doi:10.1111/j.1365-3121.2011.01022.x
Tarhan LG, Planavsky NJ, Laumer CE, Stolz JF, Reid RP (2013) Microbial mat controls on infaunal abundance and diversity in modern marine microbialites. Geobiology 11:485–497. doi:10.1111/gbi.12049
Walter MR, Heys GR (1985) Links between the rise of the metazoa and the decline of stromatolites. Precambrian Res 29:149–174. doi:10.1016/0301-9268(85)90066-X
White RA III et al (2016) Metagenomic analysis suggests modern freshwater microbialites harbor a core and distinct microbial community. Front Microbiol 6:1531. doi:10.3389/fmicb.2015.01531
Acknowledgments
The authors thank the numerous field and laboratory assistants, taxonomic experts (including Charles Griffiths, Carol Simon, Torkild Bakken, Lucio Bonato, Nelson Miranda, Magdalena BŁażewicz-Paszkowycz, Christer Erseus) and the Centre for High Resolution Transmission Electron Microscopy at the Nelson Mandela Metropolitan University, South Africa, who helped with this research. This research is funded by the South African Research Chairs Initiative (SARChI) of the Department of Science and Technology (DST) and the National Research Foundation (NRF) of South Africa (Unique Grant No. 84375); the NRF also provided a bursary to GMR. The three anonymous reviewers are thanked for their constructive comments which helped improve an earlier version of this manuscript.
Author contribution statement
GMR drafted the manuscript as well as collected and processed the data; RP conceived, designed and coordinated the study, collected field data, and helped draft the manuscript; MSB helped draft the manuscript and facilitated the process of sample collection to laboratory processing. All authors gave final approval for publication.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Ulrich Sommer.
This study clearly demonstrates that metazoans are not always restrictive of stromatolite layering, as several evolutionary hypotheses suggest, but may rather utilize microbialites as micro-refugia.
Rights and permissions
About this article

Cite this article
Rishworth, G.M., Perissinotto, R. & Bird, M.S. Coexisting living stromatolites and infaunal metazoans. Oecologia 182, 539–545 (2016). https://doi.org/10.1007/s00442-016-3683-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00442-016-3683-5