
Abstract
Behavioral syndromes arise when individual behavior is correlated over time and/or across environmental contexts, often resulting in inter-population behavioral differences. Three main hypotheses have been suggested to explain the evolution of behavioral syndromes. The constraint hypothesis suggests that behaviors originate from a shared mechanism with a strong genetic or physiological basis. In contrast, according to the adaptive hypothesis, behavioral syndromes depend on specific selective pressures in each environment, and thus should evolve when specific behavioral combinations are advantageous. Finally, behavioral syndromes can also arise owing to neutral stochastic processes. We tested here for variation in the foraging syndromes of pit-building antlions originating from different populations along a climatic gradient. Although inter-population variation existed in some traits, foraging syndromes were similar across populations, supporting the constraint hypothesis. These findings suggest that stabilizing selection, acting on the foraging behavior of antlions during their larval phase, outweighs local selection pressures, resulting in “constraint syndromes.” We also explored behavioral repeatability of foraging-related traits within and among habitats (natural, novel and disturbed habitats), and detected different levels of repeatability: pit diameter was more repeatable than response time to prey, followed by prey exploitation efficiency. Behavioral repeatability of the same trait differed according to context, suggesting that repeatability is a trait in itself and should not be considered identical even when studying the same behavioral trait.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Behavioral syndromes evolve when individual behaviors are correlated over time and/or across environmental and ecological contexts (Gosling 2001; Réale et al. 2007; Sih et al. 2004). That is, although this phenomenon refers to the individual level, it is a population feature (Bell 2007). Consistent behavior can also spillover to unfavorable conditions, resulting in suboptimal behavior (Duckworth 2006; Quinn and Cresswell 2005). Such behavioral spillovers, resulting from a link between traits, can explain the maintenance of several different behavioral types within a population (Dingemanse and Wolf 2010; Wolf and Weissing 2010). Understanding how behavioral syndromes evolve requires exploring whether and to what extent such “suites” of behavioral traits are consistent under different environmental contexts (Bell 2007; Dall et al. 2012; Sih et al. 2004).
Three main hypotheses have been suggested to explain the evolution of behavioral syndromes (Bell 2005). According to the constraint hypothesis, behaviors originate from a shared and fixed mechanism, such as the pleiotropic effects of genes. Therefore, correlated behaviors should be viewed as a “suite” of traits, rather than independent ones (Price and Langen 1992; Wilson et al. 2010). The mechanisms underlying such correlated traits assume a genetic and/or physiological basis: a change in one behavior should thus lead to a shift in another linked behavior. For example, Pruitt et al. (2010) found that geographically distant populations of the socially polymorphic spider Anelosimus studiosus share similar syndromes, suggesting a constraint on the evolution of behavior in this species.
The alternative adaptive hypothesis suggests that behavioral syndromes depend on the local selective pressures characterizing the environment, leading to differences among habitats. In other words, behavioral syndromes should evolve when some combinations of behavioral traits are locally adaptive (Bell and Sih 2007; Dingemanse et al. 2004; Dochtermann and Jenkins 2007), and are thus expected to be population specific (Bell 2007; Wilson 1998). Dingemanse et al. (2007) tested for variation in behavioral syndromes among six predator-naïve and six predator-sympatric populations of the three-spined stickleback Gasterosteus aculeatus. They found correlations between activity, aggression, and exploration in the predator-sympatric populations, but not in the predator-naïve ones. Their observations indicated inter-population variation in behavioral syndromes, rejecting the constraint hypothesis, while suggesting that distinct trait combinations are adaptive in different environments. Population variation in behavioral syndromes might also arise due to neutral stochastic processes (Armbruster and Schwaegerle 1996; Whitlock et al. 2002). If the populations under study exhibit variation in their syndromes, the constraint hypothesis should be rejected; nonetheless, in order to support the adaptive hypothesis, the syndromes should be examined and explained in light of the specific environmental conditions.
Geographic variation in behavior is common (e.g., Foster 1999; Foster and Endler 1999). Such variation evolves due to varying biotic and abiotic conditions prevailing in different environments, which translate into differential selective pressures (Endler and Houde 1995; Refsnider et al. 2014). Therefore, investigating different populations along a geographic gradient should challenge the phenomenon of behavioral syndromes and serve as an excellent platform to test the constraint and adaptive/stochastic hypotheses (Bell 2005).
Two commonly studied environmental gradients are those of climate and predation risk. The former involves latitudinal or altitudinal clines, which affect the life history, morphology, and physiology of organisms (Chown and Gaston 1999; Stillwell 2010). Ectotherms at higher latitudes are often, but not always, larger, take longer to develop, and are more resistant to starvation (Arnett and Gotelli 1999b; Roff 2002). The opposite pattern (smaller body size) is expected when animals in low-temperature environments compensate for short seasons by short development, resulting in a smaller body size [the converse Bergmann’s cline (Arnett and Gotelli 1999a; Blanckenhorn and Demont 2004)]. Grasshoppers from northern populations, for example, develop faster, have higher growth rates and reach a smaller body size than individuals from southern populations (Parsons and Joern 2014). Latitudinal variation is also studied in the context of adaptations to stress. Karan and Parkash (1998) found higher desiccation tolerance among the Drosophila kikkawai populations of northern India, probably due to the high summer temperatures characterizing this region. Higher starvation endurance was evident in D. kikkawai populations of southern India, experiencing more favorable environmental conditions, but that are probably subjected to stronger competition.
The current study had the following goals: (1) to explore behavioral repeatability in foraging-related traits of pit-building antlion larvae, originating from different populations along a climatic gradient, within and among habitats (natural, novel, and disturbed habitats); (2) to determine whether foraging syndromes are consistent (a constraint) or differ (adaptive/stochastic) among these populations; and (3) to test for clinal variation in morphology and link it to behavior. As a model system we used the pit-building antlion Myrmeleon hyalinus (Neuroptera: Myrmeleontidae), which exhibits substantial clinal variation in behavioral, morphological, and life history traits along a classic Mediterranean-to-desert climate gradient (Rotkopf and Ovadia 2014; Scharf et al. 2008, 2009a).
Pit-building antlion larvae are opportunistic predators that depend heavily on their physical environments (Farji-Brener 2003; Gotelli 1993; Scharf and Ovadia 2006). Substrate particle size is an example of such dependency, as it affects metabolic costs of pit construction and maintenance, the time required for capturing the prey and the prey-escape probability (Devetak et al. 2012; Lucas 1982, 1985b). Therefore, antlion larvae prefer substrates with a specific grain size (Farji-Brener 2003; Klokočovnik et al. 2012). Temperature is another important abiotic factor affecting foraging performance: the larvae of the antlion Myrmeleon immaculatus reared at higher temperatures constructed and maintained pits more frequently than those reared at lower temperatures (Arnett and Gotelli 2001). Because antlion larvae have not yet reached the reproductive stage, they focus their efforts on foraging (Eltz 1997; Scharf et al. 2011). Consequently, their foraging behavior should be under a strong selection pressure, and repeatable behaviors are expected to evolve. If syndromes stem from a shared mechanism of specific links or trade-offs between traits, the foraging syndromes of M. hyalinus should be consistent among different populations along the climatic gradient. Alternatively, if distinct behavioral correlations were favored in different antlion populations, or if these correlations evolved independently through neutral stochastic processes, we should detect inter-population variation in foraging syndromes.
Materials and methods
Study species and habitat of origin
Myrmeleon hyalinus is the most abundant pit-building antlion in Israel (Simon 1988). The larvae construct their pits in shaded areas under small trees and bushes, and wait for small arthropod prey to fall into their traps. They undergo a complete metamorphosis: the larval stage lasts up to 1 year and includes three instar stages, this is followed by the pupal stage (~1 month) and then weak-flying and short-lived adults emerge (Scharf et al. 2009a). During September 2013, we collected M. hyalinus larvae from different sandy sites along Israel’s climatic gradient (Fig. 1): two Mediterranean populations—Caesarea (32°29′N,34°54′E) and Zikim (31°37′N, 34°31′E); and two desert populations—Mashabei Sadeh (31°00′N, 34°46′E) and Holot Agur (30°56′N, 34°24′E). This climatic gradient is characterized by a decrease in relative humidity and rate of annual rainfall and an increase in temperature (Goldreich 2003; pp 56, 72, 99, 123), accompanied by a decrease in plant cover (Abramsky 1988; Dall’Olmo and Karnieli 2002) and in insect diversity (Sogavker 2004).

Sites (filled triangle) from which Myrmeleon hyalinus larvae were collected: Caesarea (1), Zikim (2), Mashabei Sadeh (3) and Holot Agur (4). Sandy regions are marked in gray. Major cities are marked with circles: Haifa (H), Tel Aviv (TA), Jerusalem (J), Beer Sheva (BS) and Eilat (E)
In order to examine the prey capture success of antlions, we collected 300 individuals of Messor ebeninus (Hymenoptera: Formicidae) from Nahal Secher (31°60′N, 34°49′E), a semi-arid area located 15 km south of the city of Beer-Sheva. All these ants were collected from the field in the morning of the prey-capture-success assay, and thus there was no need to maintain them in the collection boxes for more than 2 h under laboratory conditions. Messor ebeninus is a common harvester ant in Israel, and a natural prey of Myrmeleon hyalinus. This ant species lives in subterranean nests and its foragers consume seeds and other plant materials (Ofer 2000).
Experimental design
During the first month of the experiment each antlion larva was fed once a week with one mealworm larva (Tenebrio molitor) and then weighed using an analytical scale (CP224S, accuracy of 0.1 mg; Sartorius, Göttingen, Germany), in order to select individuals with similar body masses (48 individuals from Zikim and 60 individuals from each of the other three populations; n = 48 + 3 × 60 = 228). Next, the antlion larvae were starved for 5 days in small plastic containers (8.5-cm diameter, filled with 5-cm-depth substrate). This entire habituation period was used to standardize their body condition (Scharf et al. 2009b). All larvae were placed in a small transparent petri dish, with a graph paper underneath it, and then photographed using a digital camera (Micropublisher 5.0; QImaging, Surrey, BC), connected to a stereoscope (SMZ 800; Nikon, Kawasaki). Using the software Image J (Abràmoff et al. 2004), we measured five morphological traits: head width (HW), mandible width (MW), mandible length (ML), abdomen width (AW), and abdomen length (AL). The two first traits were used to identify and select only the third-instar larvae (Alcalay et al. 2014a; Scharf et al. 2008; Simon 1988), while all five traits were used to explore morphological differences among third-instar larvae originating from different populations along Israel’s climate gradient. Experiments were performed under an identical day/night photoperiod (12:12 h), temperature of 25.0 °C ± 0.5 °C, and relative humidity of 59.3 % ± 0.78 (averages and SDs of three daily measurements in the test room).
We examined the performances of individuals from the four populations in their natural habitat (sandy soil) and in a novel habitat (wheat semolina, with coarser particle sizes than sand). Wheat semolina particle size was as follows: 45.71 % >0.71 mm, 49.01 % 0.71–0.50 mm, 2.00 % 0.50–0.42 mm, and 3.28 % <0.42 mm. Sand particles were differently distributed: 7.97 % >0.25 mm, 78.65 % 0.125–0.25 mm, 11.54 % 0.062–0.125 mm, and 1.84 % <0.062 mm (Danin 1978). We aimed at examining if and to what extent the behavioral repeatability and syndromes change when the antlion larvae face a novel substrate that they would never have encountered under natural conditions. Since, M. hyalinus is a habitat generalist, naturally occurring in both fine (e.g., loess) and coarse (e.g., sand) textured soils (Barkae et al. 2012), we used the artificial substrate, wheat semolina, as the novel habitat. We focused on the following behavioral traits: (1) pit diameter, recorded using a digital caliper (±0.1 mm); (2) response time to prey: we placed a single mealworm within the antlion’s pit and measured the time it took the antlion to respond by throwing sand. If the antlion did not respond after 60 s, we used this maximal value as the response time, assuming that it was not interested in the prey (similar to Scharf et al. 2010); and (3) prey exploitation efficiency, calculated by dividing the difference in antlion mass before and 1 day after feeding by the prey body mass (Scharf et al. 2009b). The value of prey exploitation efficiency ranges between 0 (no consumption) and 1 (complete consumption of the prey), and reflects the antlion efficiency in translating prey mass into its own body mass. Alternatively, considering each mealworm as a patch, the decision to stop consuming the prey may be equivalent to the decision to leave a patch and continue to the next one (Brown 1988; Lucas 1985a).
Response time to prey and pit diameter were measured twice for both natural and novel habitats. In order to minimize the possible effect of prey size on the response time, we selected mealworms of similar size in the first trail (the prey was removed after the response time was measured) and distributed them randomly. In the second trail, antlions were fed with mealworms that were weighed before (0.014–0.040 g), to calculate the prey exploitation efficiency measurement. Prey exploitation efficiency was measured only once, because over-feeding of the antlions might have induced an undesirable variance in body masses between populations. We also examined the prey capture success of antlions encountering a natural ant prey. To this end, we placed a single M. ebeninus within the antlion’s pit and recorded whether it managed to capture the ant, or whether the ant escaped from the pit.
Next, we examined the change in pit diameter after disturbance. Each individual was placed in a separate experimental container for 4 h. We used a small aspirator that releases air at low pressure to disturb each pit. The aspirator was activated 5 cm above the experimental cup until the pit was completely destroyed. This manipulation was conducted three times, at 1-h intervals. Pits were measured twice: 2 h after the final disturbance, and in the following morning. Antlions in nature often suffer from frequent disturbances caused by relocating conspecifics, other moving animals, wind, rainfall, and litter falling from bushes and trees (Barkae et al. 2010; Farji-Brener et al. 2008). Here we aimed at mimicking such successive natural disturbances in order to investigate their effects on the decision of antlions to construct pits.
Statistical analysis
Behavioral repeatability
Repeatability is the pre-condition for the establishment of behavioral syndromes. We used intra-class correlation coefficients (ICC) (Hayes and Jenkins 1997) in order to examine repeatability at each of the following levels:
-
1.
Within habitats (natural and novel), considering pit diameter and response time to prey. Each behavior was tested twice at an interval of 1 day.
-
2.
Between natural and novel habitats, considering pit diameter, response time to prey, and prey exploitation efficiency.
-
3.
Between natural and disturbed habitats, considering pit diameter.
-
4.
Between behavioral traits, considering all populations together.
We calculated ICC to determine which behavioral traits are generally more repeatable than others.
In order to obtain a single trait value per population per habitat, we averaged the two values of pit diameter for each habitat (natural, novel, and disturbed), based on its high repeatability, resulting in three values. We then took the minimal value of the response time to prey, because of its low repeatability within habitats, which also resulted in three values, one per habitat. Growth efficiency was measured only once per habitat, and this was the value we used for further analyses.
Behavioral differences between populations, single traits
We used either the average or minimal traits, as noted above. Behavioral differences were compared using median tests (Zar 1999), due to their skewed distribution and the high variation of the data. Analyses of behavioral traits in the novel and disturbed habitats were computed relative to performance in the natural habitat. Thus, we calculated the proportional change of each trait in the novel/disturbed habitat relative to the value of the same trait in the natural habitat: \( \frac{{{\text{Trait}}\,X_{\text{novel/disturbed}} - {\text{Trait}}\,X_{\text{natural}} }}{{{\text{Trait}}\,X_{\text{natural}} }}. \)
Seven median tests were performed, two for pit diameter, response time to prey and prey exploitation efficiency (in natural and novel habitats), and one for pit diameter after disturbance (only in the natural habitat). Two extreme values (>75th percentile + 3IQ or <25th percentile − 3IQ; IQ = 75th − 25th percentile) of proportional change in pit diameter (one in the novel habitat and another one in the disturbed habitat) and three extreme values of proportional change in prey exploitation efficiency (all three in the novel habitat) were removed from the analyses. Capture success of natural prey was compared between populations using a Pearson χ2-test.
Foraging syndromes
We first performed a principal component analysis (PCA) on all behavioral traits across populations (seven behavioral traits). Next, we performed a PCA on each population separately, to determine whether the same general pattern holds for each one of them. If it does, it supports the constraint hypothesis, whereas if each population shows a different syndrome, it supports adaptive/stochastic hypotheses. Prior to the PCA we applied a Fisher’s Z transformation (Zar 1999). We referred only to the PC axes with eigenvalues larger than one and explained/referred in each PC axis only to loadings larger than 0.3 (Tabachnick and Fidell 2007).
Morphological differences between populations
We used PCA, similar to the analysis described above, to examine morphological differences between populations, comprising the five measured traits (HW, ML, MW, AL and AW). Following the PCA we used two one-way ANOVAs on the two important PC axes, with population as the explanatory variable.
All statistical analyses were performed in SYSTAT version 12 (SYSTAT software) and STATISTICA, version 12.0. We also used R (R Core Team 2014) package psy developed by Falissard (2012) (http://cran.r-project.org/web/packages/psy/index.html) to calculate the ICC and its 95 % confidence intervals.
Results
Behavioral repeatability
-
1.
Within habitats the main difference in repeatability was based on the behavioral trait measured (Table 1). Pit diameter was highly repeatable across all populations in the natural habitat and in three out of four populations in the novel habitat. Repeatability in pit diameter was lower in the novel habitat than in the natural one. In contrast, we detected low behavioral repeatability in response time to prey in both the natural and novel habitats.
-
2.
Between natural and novel habitats pit diameter was repeatable in all populations. In contrast, neither response time to prey nor prey exploitation efficiency were repeatable between natural and novel habitats in any of the populations (Table 2).
-
3.
Between natural and disturbed habitats pit diameter was repeatable in all populations except that from Holot Agur (Table 2).
-
4.
Between behavioral traits we detected significant and highly repeatable behavior considering pit diameter, medium-level repeatability in response time to prey, and low repeatability in prey exploitation efficiency between habitats (Table 3).
Behavioral differences between populations, single traits
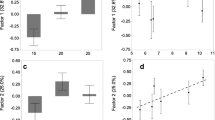
In the natural habitat, the semi-arid population (Mashabei Sadeh) constructed the largest pits, followed by the arid population (Holot Agur), and the southern Mediterranean one (Zikim). The northern Mediterranean population (Caesarea) constructed the smallest pits (χ 2 = 10.29, df = 3, P = 0.016; Fig. 2a). Prey capture success gradually decreased along the north-to-south gradient (χ 2 = 11.54, df = 3, P = 0.009; Fig. 2b). Prey exploitation efficiency of the southern Mediterranean population (Zikim) tended to be higher than that of the semi-arid population (Mashabei Sadeh), but this pattern was marginally non-significant (χ 2 = 7.29, df = 3, P = 0.063). Finally, we could not detect any difference in the response time to prey between populations (χ 2 = 3.10, df = 3, P = 0.376).

Behavioral differences among populations: a median pit diameter in the natural habitat. Boxes represent the inter-quartile range, vertical bars represent 95th and 5th percentiles, and black dots the outliers. b Prey capture success when encountering natural ant prey
Both pit diameter and prey exploitation efficiency were higher in the novel than in the natural habitat (Fig. 3a, b). The proportional change in pit diameter (novel relative to natural) was higher in the Mediterranean than in the desert populations (χ 2 = 11.95, df = 3, P = 0.007; Fig. 3a). In contrast, the proportional change in prey exploitation efficiency was higher in the desert than in the Mediterranean populations (χ 2 = 9.22, df = 3, P = 0.026; Fig. 3b). Finally, we could not detect any difference in the proportional change in the response time to prey among populations (χ 2 = 0.15, df = 3, P = 0.985). Following disturbance, pit diameters of all populations became smaller relative to the natural habitat; however, the proportional change differed among populations, with the largest difference between the semi-arid (Mashabei Sadeh) and the southern Mediterranean populations (Zikim) (χ 2 = 8.77, df = 3, P = 0.032; Fig. 3c).

Median proportional changes in a pit diameter and in b prey exploitation efficiency in the novel relative to the natural habitat, and in c pit diameter after disturbance relative to the natural habitat. Boxes represent the inter-quartile range, vertical bars represent 95th and 5th percentiles, and black dots the outliers
Foraging syndromes
The PCA analyses revealed an unequal number of significant PC axes (eigenvalue larger than one) for the different populations. We sought to compare the general foraging syndrome with those of different populations, and thus focused on comparing only the first PC axis for each population; see Appendix 1 for all PC axes, performed on separate populations and the general syndrome. The first PC axis of the general foraging syndrome explained 32.11 % of the variance, and the eigenvalue was 2.09. This PC was composed of positive values of pit diameter (in all habitats) and prey exploitation efficiency in the novel habitat, and negative values of response time to prey in both the natural and novel habitats (Table 4). It therefore represents foraging performance: individuals with high loading on this axis constructed larger pits, responded faster to prey and exploited their prey efficiently in novel habitat. We detected similar patterns also when the analysis was performed on each population separately (Table 4). Specifically, the values of pit diameter in three populations in the natural habitat and those of all four populations in the novel and disturbed habitats were positive, similar to the general foraging syndrome. Response time to prey had a negative sign in both habitats in the general foraging syndrome. This pattern was also true in three populations in the natural habitat and in two populations in the novel habitat. Finally, prey exploitation efficiency had a positive sign in the novel habitat in the general foraging syndrome, showing the same pattern in two out of the four populations.
Morphological differences between populations
The first PC axis of the morphological PCA explained 49.51 % of the variance and was composed of positive values of all five morphological traits, thus representing body size (Table 5). The second PC axis explained 27.50 % of the variance with positive values of both AW and AL and negative values of HW and ML (Table 5). This PC axis reflects the body reserves in the abdomen of the antlion larvae: individuals with high loading on this axis had larger abdomens at the expense of narrower heads and shorter mandibles. We found a significant difference in body size among the four populations (F 3,224 = 3.94, P = 0.009; Fig. 4): individuals from the arid population (Holot Agur) were smaller than those from the Mediterranean populations (Caesarea and Zikim). Body reserves differed among populations (F 3,224 = 3.54, P = 0.015; Fig. 4), with arid population individuals (Holot Agur) having larger reserves.

Morphological differences between populations in principal component PC1 (body size) vs. PC2 (body reserves)
Discussion
Here we studied for the first time the foraging syndromes of pit-building antlions along a climate gradient. We found that inter-population variation in foraging behavior sometimes followed the latitudinal cline (e.g., the decrease in prey capture success from north to south), and sometimes was simply population specific, regardless of the climate gradient. We also found morphological differences: while individuals from the arid population (Holot Agur) had the smallest body size, their body reserves were the highest (largest abdomen relative to head). Despite this inter-population variation in various traits, the foraging syndromes of the four different populations were similar to the general one, supporting the constraint hypothesis and not the adaptive/stochastic one.
Repeatability in pit diameter was stronger than that observed for response time to prey, which in turn was more repeatable than prey exploitation efficiency. This observation might be explained by the environmental sensitivity of the observed traits. Behaviors which are under morphological or physiological constraints are likely to be more repeatable than behaviors subject to the energetic state or the current social environment (Castellano et al. 2002; Smith and Hunter 2005). Therefore, pit construction, which has already been shown to be associated with the antlion’s morphology—mainly head and mandible size (Barkae et al. 2012; Scharf et al. 2009c)—might be more repeatable than energetically dependent traits such as response time to prey and prey exploitation efficiency.
We also detected lower repeatability in the novel relative to the natural habitat, especially in pit diameter. A previous study on M. hyalinus, documenting relocation distances, revealed a similar pattern—repeatability was lower in an unfamiliar, unfavorable substrate than in a familiar substrate [sand (Alcalay et al. 2014b)]. Behavioral performance usually improves with experience (Brown and Laland 2003; Rosenzweig and Bennett 1996), and it can play a role in stabilizing behavioral types via a positive feedback between behavior and reward (Dingemanse and Wolf 2010). We thus suggest that such a positive feedback increases behavioral repeatability in known situations compared to novel ones. A non-mutually exclusive explanation is that behavioral variance increases in unfamiliar conditions, as animals have not learned the optimal behavior in such cases. This is perhaps comparable to phenotypic divergence under stressful conditions (Hoffmann and Merilä 1999).
Some of the differences among populations were gradient related and can be explained by environmental changes across this climatic gradient. For example, individuals from the Mediterranean region, characterized by higher prey abundance, may simply be better foragers than individuals from desert regions, as expressed in the decrease in prey capture success from north to south. The latter finding may also be linked to morphology: individuals from Holot Agur had the smallest body size, which could also have led to their lower prey capture success. Nonetheless, although all populations increased their prey exploitation efficiency (relative to the natural habitat) when encountering a novel habitat, the desert populations utilized a higher proportion of the prey, probably in order to compensate for their smaller body size.
Not all of the observed behavioral differences were gradient related. For instance, individuals from the semi-arid population constructed the largest pits, followed by the arid population, and then the two Mediterranean populations. Pit-building antlions are sedentary animals with low dispersal ability and are thus restricted to specific habitats (Farji-Brener 2003; Gotelli 1993; Scharf and Ovadia 2006). Consequently, it is possible that specific microclimate conditions masked the geographical gradient, generating a non-latitudinal behavioral gradient, which better fits the micro-climate. For example, a shaded area under large planted trees, common in the semi-arid population (Mashabei Shadeh), may buffer against harsher climatic conditions while also increasing prey availability (Rotkopf and Ovadia 2014). This suggestion was supported by the higher behavioral and morphological similarity of the semi-arid population to the two Mediterranean populations rather to the other arid population. Alternatively, inter-population differences may be the result of a genetic drift. As noted, the adults are weak flying and short lived, and there is thus good reason to assume a low dispersal rate.
Many studies of behavioral syndromes have detected differences among populations, which can be explained by adaptive evolution processes. Such local divergence is triggered by varying predation pressure, competition intensity, or environmental heterogeneity (Bell 2005; Bengston and Dornhaus 2014; Dingemanse et al. 2007; Dochtermann et al. 2012; Herczeg et al. 2009). Here, we detected high similarity in the foraging syndromes among the four populations. During the larval phase, antlions are mainly engaged in foraging, and thus it is likely that foraging performance undergoes strong selection at the species level, outweighing local selection pressures. Due to experimental procedure considerations, we deliberately selected individuals with similar body masses. Thereby, we perhaps overlooked the link between body mass, climate gradient, and behavior (Atkinson 1994; Blanckenhorn and Demont 2004).
Our syndrome comprised only foraging-related traits in different contexts. However, behavior is also linked to physiological and life history traits, often resulting in various trade-offs (e.g., Dupont-Prinet et al. 2010; Guerra and Pollack 2007; Rotkopf et al. 2013). Therefore, the interpretation of the observed syndromes as a constraint should be viewed with caution. In other words, it is likely that the foraging syndromes are part of more complex syndromes that include physiological, life history, and morphological aspects. In summary, we detected strong behavioral and morphological differences between antlions originating from Mediterranean and desert populations along Israel’s climatic gradient. However, their foraging syndromes showed high similarity to the general syndrome, as expected by the constraint hypothesis. The next step should be to add physiological and life history traits to the syndromes detected, such as metabolic rate or starvation endurance. There is also a need for field experiments to take into consideration the natural environmental variation among populations.
Author contribution statement
Y. A., I. S. and O. O. conceived and designed the experiments. Y. A. performed the experiments. All three authors were involved in analyzing the data and writing the manuscript.
References
Abràmoff MD, Magalhães PJ, Ram SJ (2004) Image processing with ImageJ. J Biophotonics 11:36–43
Abramsky Z (1988) The role of habitat and productivity in structuring desert rodent communities. Oikos 52:107–114
Alcalay Y, Barkae ED, Ovadia O, Scharf I (2014a) Consequences of the instar stage for behaviour in a pit-building antlion. Behav Process 103:105–111
Alcalay Y, Ovadia O, Scharf I (2014b) Behavioral repeatability and personality in pit-building antlion larvae under differing environmental contexts. Behav Ecol Sociobiol 68:1985–1993
Armbruster W, Schwaegerle K (1996) Causes of covariation of phenotypic traits among populations. J Evol Biol 9:261–276
Arnett AE, Gotelli NJ (1999a) Bergmann’s rule in the ant lion Myrmeleon immaculatus DeGeer (Neuroptera: Myrmeleontidae): geographic variation in body size and heterozygosity. J Biogeogr 26:275–283
Arnett AE, Gotelli NJ (1999b) Geographic variation in life-history traits of the ant lion, Myrmeleon immaculatus: evolutionary implications of Bergmann’s rule. Evolution 53:1180–1188
Arnett AE, Gotelli NJ (2001) Pit-building decisions of larval ant lions: effects of larval age, temperature, food, and population source. J Insect Behav 14:89–97
Atkinson D (1994) Temperature and organism size—a biological law for ectotherms? Adv Ecol Res 25:1–58
Barkae ED, Scharf I, Subach A, Ovadia O (2010) The involvement of sand disturbance, cannibalism and intra-guild predation in competitive interactions among pit-building antlion larvae. Zoology 113:308–315
Barkae ED, Scharf I, Abramsky Z, Ovadia O (2012) Jack of all trades, master of all: a positive association between habitat niche breadth and foraging performance in pit-building antlion larvae. PLoS One 7:e33506
Bell AM (2005) Behavioural differences between individuals and two populations of stickleback (Gasterosteus aculeatus). J Evol Biol 18:464–473
Bell AM (2007) Future directions in behavioural syndromes research. Proc R Soc B Lond B Biol Sci 274:755–761
Bell AM, Sih A (2007) Exposure to predation generates personality in three spined sticklebacks (Gasterosteus aculeatus). Ecol Lett 10:828–834
Bengston S, Dornhaus A (2014) Be meek or be bold? A colony-level behavioural syndrome in ants. Proc R Soc Lond B Biol Sci 281:20140518
Blanckenhorn W, Demont M (2004) Bergmann and converse Bergmann latitudinal clines in arthropods: two ends of a continuum? Integr Comp Biol 44:413–424
Brown JS (1988) Patch use as an indicator of habitat preference, predation risk, and competition. Behav Ecol Sociobiol 22:37–47
Brown C, Laland KN (2003) Social learning in fishes: a review. Fish Fish 4:280–288
Castellano S, Cuatto B, Rinella R, Rosso A, Giacoma C (2002) The advertisement call of the European treefrogs (Hyla arborea): a multilevel study of variation. Ethology 108:75–89
Chown SL, Gaston KJ (1999) Exploring links between physiology and ecology at macro-scales: the role of respiratory metabolism in insects. Biol Rev Camb Philos Soc 74:87–120
Dall SRX, Bell AM, Bolnick DI, Ratnieks FL (2012) An evolutionary ecology of individual differences. Ecol Lett 15:1189–1198
Dall’Olmo G, Karnieli A (2002) Monitoring phenological cycles of desert ecosystems using NDVI and LST data derived from NOAA-AVHRR imagery. Int J Remote Sens 23:4055–4071
Danin A (1978) Plant species diversity and plant succession in a sandy area in the Northern Negev. Flora 167:409–422
Devetak D, Novak T, Janžekovič F (2012) Effect of substrate density on behaviour of antlion larvae (Neuroptera: Myrmeleontidae). Acta Oecol 43:1–7
Dingemanse NJ, Wolf M (2010) Recent models for adaptive personality differences: a review. Philos Trans R Soc Lond B Biol Sci 365:3947–3958
Dingemanse NJ, Both C, Drent PJ, Tinbergen JM (2004) Fitness consequences of avian personalities in a fluctuating environment. Proc R Soc Lond B Biol Sci 271:847–852
Dingemanse NJ, Wright J, Kazem AJ, Thomas DK, Hickling R, Dawnay N (2007) Behavioural syndromes differ predictably between 12 populations of three-spined stickleback. J Anim Ecol 76:1128–1138
Dochtermann NA, Jenkins SH (2007) Behavioural syndromes in Merriam’s kangaroo rats (Dipodomys merriami): a test of competing hypotheses. Proc R Soc Lond B Biol Sci 274:2343–2349
Dochtermann NA, Jenkins SH, Swartz MJ, Hargett AC (2012) The roles of competition and environmental heterogeneity in the maintenance of behavioral variation and covariation. Ecology 93:1330–1339
Duckworth RA (2006) Behavioral correlations across breeding contexts provide a mechanism for a cost of aggression. Behav Ecol 17:1011–1019
Dupont-Prinet A, Chatain B, Grima L, Vandeputte M, Claireaux G, McKenzie D (2010) Physiological mechanisms underlying a trade-off between growth rate and tolerance of feed deprivation in the European sea bass (Dicentrarchus labrax). J Exp Biol 213:1143–1152
Eltz T (1997) Foraging in the ant-lion Myrmeleon mobilis Hagen 1888 (Neuroptera: Myrmeleontidae): behavioral flexibility of a sit-and-wait predator. J Insect Behav 10:1–11
Endler JA, Houde AE (1995) Geographic variation in female preferences for male traits in Poecilia reticulata. Evolution 49:456–468
Falissard B (2012) psy: Various procedures used in psychometry. R package version 1.1. http://CRAN.R-project.org/package=psy
Farji-Brener AG (2003) Microhabitat selection by antlion larvae, Myrmeleon crudelis: effect of soil particle size on pit-trap design and prey capture. J Insect Behav 16:783–796
Farji-Brener AG, Carvajal D, Gei MG, Olano J, Sanchez JD (2008) Direct and indirect effects of soil structure on the density of an antlion larva in a tropical dry forest. Ecol Entomol 33:183–188
Foster SA (1999) The geography of behaviour: an evolutionary perspective. Trends Ecol Evol 14:190–195
Foster SA, Endler JA (1999) Thoughts on geographic variation in behavior. In: Foster SA, Endler JA (eds) Geographic variation in behavior. Oxford University Press, New York
Goldreich Y (2003) The climate of Israel: observation, research, and application. Kluwer Academic/Plenum, New York
Gosling SD (2001) From mice to men: what can we learn about personality from animal research? Psychol Bull 127:45–86
Gotelli NJ (1993) Ant lion zones: causes of high-density predator aggregations. Ecology 74:226–237
Guerra PA, Pollack GS (2007) A life history trade-off between flight ability and reproductive behavior in male field crickets (Gryllus texensis). J Insect Behav 20:377–387
Hayes JP, Jenkins SH (1997) Individual variation in mammals. J Mamm 78:274–293
Herczeg G, Gonda A, Merilä J (2009) Predation mediated population divergence in complex behaviour of nine-spined stickleback (Pungitius pungitius). J Evol Biol 22:544–552
Hoffmann AA, Merilä J (1999) Heritable variation and evolution under favourable and unfavourable conditions. Trends Ecol Evol 14:96–101
Karan D, Parkash R (1998) Desiccation tolerance and starvation resistance exhibit opposite latitudinal clines in Indian geographical populations of Drosophila kikkawai. Ecol Entomol 23:391–396
Klokočovnik V, Devetak D, Orlačnik M (2012) Behavioral plasticity and variation in pit construction of antlion larvae in substrates with different particle sizes. Ethology 118:1102–1110
Lucas JR (1982) The biophysics of pit construction by antlion larvae (Myrmeleon, Neuroptera). Anim Behav 30:651–664
Lucas JR (1985a) Partial prey consumption by antlion larvae. Anim Behav 33:945–958
Lucas JR (1985b) Metabolic rates and pit-construction costs of two antlion species. J Anim Ecol 54:295–309
Ofer J (2000) Let’s go to the ant. A field guide to the ants of Israel (in Hebrew). Yuval Ofer, Jerusalem
Parsons SM, Joern A (2014) Life history traits associated with body size covary along a latitudinal gradient in a generalist grasshopper. Oecologia 174:379–391
Price T, Langen T (1992) Evolution of correlated characters. Trends Ecol Evol 7:307–310
Pruitt JN, Riechert SE, Iturralde G, Vega M, Fitzpatrick BM, Aviles L (2010) Population differences in behaviour are explained by shared within-population trait correlations. J Evol Biol 23:748–756
Quinn JL, Cresswell W (2005) Personality, anti-predation behaviour and behavioural plasticity in the chaffinch Fringilla coelebs. Behaviour 142:1377–1402
R Core Team (2014) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. http://www.R-project.org/
Réale D, Reader SM, Sol D, McDougall PT, Dingemanse NJ (2007) Integrating animal temperament within ecology and evolution. Biol Rev 82:291–318
Refsnider JM, Milne-Zelman C, Warner DA, Janzen FJ (2014) Population sex ratios under differing local climates in a reptile with environmental sex determination. Evol Ecol 28:977–989
Roff DA (2002) Life history evolution. Sinauer, Sunderland
Rosenzweig MR, Bennett EL (1996) Psychobiology of plasticity: effects of training and experience on brain and behavior. Behav Brain Res 78:57–65
Rotkopf R, Ovadia O (2014) Examining growth rate and starvation endurance in pit-building antlions from Mediterranean and desert regions. Ecol Entomol 39:94–100
Rotkopf R, Alcalay Y, Bar-Hanin E, Barkae ED, Ovadia O (2013) Slow growth improves compensation ability: examining growth rate and starvation endurance in pit-building antlions from semi-arid and hyper-arid regions. Evol Ecol 27:1129–1144
Scharf I, Ovadia O (2006) Factors influencing site abandonment and site selection in a sit-and-wait predator: a review of pit-building antlion larvae. J Insect Behav 19:197–218
Scharf I, Filin I, Golan M, Buchshtav M, Subach A, Ovadia O (2008) A comparison between desert and Mediterranean antlion populations: differences in life history and morphology. J Evol Biol 21:162–172
Scharf I, Filin I, Ben-Yehoshua D, Ovadia O (2009a) Phenotypic plasticity and variation in morphological and life-history traits of antlion adults across a climatic gradient. Zoology 112:139–150
Scharf I, Filin I, Ovadia O (2009b) A trade-off between growth and starvation endurance in a pit-building antlion. Oecologia 160:453–460
Scharf I, Golan B, Ovadia O (2009c) The effect of sand depth, feeding regime, density, and body mass on the foraging behaviour of a pit-building antlion. Ecol Entomol 34:26–33
Scharf I, Barkae ED, Ovadia O (2010) Response of pit-building antlions to repeated unsuccessful encounters with prey. Anim Behav 79:153–158
Scharf I, Lubin Y, Ovadia O (2011) Foraging decisions and behavioural flexibility in trap-building predators: a review. Biol Rev 86:626–639
Sih A, Bell AM, Johnson JC, Ziemba RE (2004) Behavioral syndromes: an integrative overview. Q Rev Biol 79:241–277
Simon D (1988) Ant-lions (Neuroptera: Myrmeleontidae) of the coastal plain: systematical, ecological, and zoogeographical aspects with emphasis on the coexistence of a species guild of the unstable dunes. Ph.D. thesis, Tel-Aviv University, Israel
Smith MJ, Hunter D (2005) Temporal and geographic variation in the advertisement call of the booroolong frog (Litoria booroolongensis: Anura: Hylidae). Ethology 111:1103–1115
Sogavker U (2004) The relationship between productivity and ant-species richness at regional and local scales. M.Sc. thesis, Ben-Gurion University of the Negev, Israel
Stillwell RC (2010) Are latitudinal clines in body size adaptive? Oikos 119:1387–1390
Tabachnick B, Fidell L (2007) Using multivariate statistics, 5th edn. Pearson, Boston
Whitlock MC, Phillips PC, Fowler K (2002) Persistence of changes in the genetic covariance matrix after a bottleneck. Evolution 56:1968–1975
Wilson DS (1998) Adaptive individual differences within single populations. Philos Trans R Soc Lond B Biol Sci 353:199–205
Wilson AD, Whattam EM, Bennett R, Visanuvimol L, Lauzon C, Bertram SM (2010) Behavioral correlations across activity, mating, exploration, aggression, and antipredator contexts in the European house cricket, Acheta domesticus. Behav Ecol Sociobiol 64:703–715
Wolf M, Weissing FJ (2010) An explanatory framework for adaptive personality differences. Philos Trans R Soc Lond B Biol Sci 365:3959–3968
Zar JH (1999) Biostatistical analysis, 4th edn. Prentice Hall, New Jersey
Acknowledgments
The research leading to this manuscript was partially funded by a start-up grant of the US-Israel Binational Science Foundation no. 2013086 to I. S.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Roland A. Brandl.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Alcalay, Y., Scharf, I. & Ovadia, O. Foraging syndromes and trait variation in antlions along a climatic gradient. Oecologia 178, 1093–1103 (2015). https://doi.org/10.1007/s00442-015-3284-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00442-015-3284-8