
Abstract
Mistletoes constitute instructive study cases with which to address the role of generalist consumers in the study of plant–animal interactions. Their ranges of safe sites for recruitment are among the most restricted of any plant; therefore, frugivores specializing in mistletoe have been considered almost indispensable for the seed dispersal of these parasitic plants. However, the absence of such specialists in numerous regions inhabited by many mistletoe species raises the question of whether unspecialized vectors may successfully disperse mistletoe seeds to narrowly defined safe sites. Using the European mistletoe Viscum album subsp. austriacum as a study case, we recorded a broad range of 11 bird species that disperse mistletoe seeds. For these species, we studied the mistletoe-visitation rate and feeding behavior to estimate the quantity component of dispersal effectiveness, and the post-foraging microhabitat use, seed handling, and recruitment probabilities of different microhabitats as a measure of the quality component of effectiveness. Both endozoochory and ectozoochory are valid dispersal mechanisms, as the seeds do not need to be ingested to germinate, increasing seed-dispersal versatility. Thrushes were the most effective dispersers, although they were rather inefficient, whereas small birds (both frugivores and non-frugivores) offered low-quantity but high-quality services for depositing seeds directly upon safe sites. As birds behave similarly on parasitized and non-parasitized hosts, and vectors have broad home ranges, reinfection within patches and the colonization of new patches are ensured by an ample assemblage of generalist birds. Thus, a parasitic plant requiring precision in seed dispersal can rely on unspecialized dispersers.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Most theoretical and empirical studies on plant–animal interactions have traditionally outlined the role of specialization as a prominent feature of interactive systems. However, an increasing amount of scientific evidence strongly supports the idea that generalization is more frequent than hitherto thought, with most plants interacting with multiple types of pollinators, seed dispersers, and herbivores (Herrera 1988; Waser et al. 1996; Gómez and Zamora 2000; Olesen and Jordano 2002; Bascompte and Jordano 2007). Mistletoes are widely distributed worldwide as common colonizers of tree canopies and constitute excellent models with which to examine the issue of generalization versus specialization. Their parasitic nature limits them to a narrowly defined range of safe sites for seed recruitment, i.e., particular branch diameters of specific host species (Reid 1989; Van 1993; Sargent 1995; Norton and de Lange 1999; Aukema and Martínez de Río 2002; Arruda et al. 2006). Such strong specificity suggests that mistletoes must depend on highly precise dispersal mechanisms to guide their seeds to safe sites. In this regard, many frugivore species reportedly have close mutualist ties with mistletoes, exchanging effective seed-dispersal services for food rewards (Reid 1990; Wenny 2001; Amico and Aizen 2000; Carlo and Aukema 2005), reflecting a likely coevolution (Reid 1991; Aukema and Martínez del Río 2002). However, this tight dependence of mistletoes on specialized vectors has been increasingly questioned, since many dietary generalists have been described as frequent mistletoe seed dispersers (e.g., Restrepo 1987; Reid 1991; Hawksworth and Wiens 1996; Zuber 2004; Rawsthorne et al. 2011; Arruda et al. 2012). In fact, several mistletoe species inhabiting large territories where adept vectors are completely absent depend exclusively on such dietary generalists, e.g., most mistletoe species dwelling in the Northern Hemisphere, on many ocean islands, and in several regions of the Southern Hemisphere (Reid 1991; Watson and Rawsthorne 2013).
Mistletoes relying on specialist vectors—all bird species (Restrepo et al. 2002)—may take advantage of the exclusive, direct seed dispersal of a few legitimate dispersers (Davidar 1983; Reid 1989; Martínez del Río et al. 1995; Sargent 1995; Larson 1996). These birds, with their extremely restricted diet, ensure that mistletoe fruits are consumed, as they subsist almost completely on them (Reid 1991; Restrepo et al. 2002; Watson 2004); they also guarantee high-quality dispersal services due to special anatomical and behavioral adaptations (Walsberg 1975; Richardson and Wooller 1988) which enhance seed germination or seedling establishment after seeds pass through the digestive tract (e.g., Martínez del Río and Restrepo 1993; Murphy et al. 1993) and encourage seedling recruitment by depositing seeds on safe sites (e.g., Reid 1989; Wenny 2001). On the other hand, mistletoes that depend on generalist vectors may risk the uncertainty that fruits may not be consumed or seeds may not arrive at suitable habitats (Reid 1989; Larson 1996; Montaño-Centellas 2012; Watson 2012). Despite the frequency of these parasitic plants, the way in which mistletoes that rely exclusively on generalists achieve their dissemination remains unknown.
In this study, we investigate dispersal by a group of birds that feed on the mistletoe Viscum album subsp. austriacum. This parasitic plant is widely distributed across Europe, where no mistletoe specialists exist and, therefore, where seed dispersal depends exclusively on generalists (Snow and Snow 1984, 1988; Zuber 2004; Watson and Rawsthorne 2013). We explored how this bird assemblage affects the demography of mistletoe by analyzing relative dispersal effectiveness [i.e., the contribution of a seed disperser to mistletoe fitness, sensu Schupp (1993)] and efficiency (i.e., the probability that a mistletoe seed will arrive at a safe site). For each mistletoe visitor, we studied the visitation rate and feeding behavior to estimate the “quantity” component of the effectiveness; also we examined post-foraging microhabitat use and the consequent seed shadows to assess the “quality” component of effectiveness. Concurrently, through an experimental approach, we quantified recruitment probabilities of different microhabitats within suitable hosts. We also noted whether these birds were capable of depositing seeds at appropriate safe sites beyond the mother plant, acting as colonizers of new infection foci. With this approach, we seek to understand how a parasitic plant requiring precision in seed dispersal can rely on an imprecise, unspecialized dispersal assemblage for reinfection (establishing “infection centers”), as well as for transmission to suitable new hosts.
Materials and methods
Study system and site
Viscum album subsp. austriacum (Wiesb.) Vollman, a mistletoe species native to most regions of Europe, specializes in parasitizing conifers, mainly Pinus species across its distribution range. The study site, the Sierra de Baza (2º51′48″W, 37º22′57″N), represents the southernmost limit of its geographical distribution. This is a mountainous natural reserve of southeastern Spain, which shows a sharp altitudinal gradient of 850–2270 m. The climate is typically Mediterranean, with hot, dry summers (June–September), cold winters (December–March), and rainy autumns and springs. Pine is the dominant forest tree, with Austrian pine (Pinus nigra Arn.) and Scots pine (Pinus sylvestris L.) being major hosts for V. album subsp. austriacum at this site. Other pines, Aleppo (Pinus halepensis Mill.) and maritime (Pinus pinaster Ait.), as well as non-conifer tree species, holm oaks (Quercus ilex L.) and Acer opalus L. subsp. granatense Boiss., are also abundant. As a bird-dispersed plant, V. album subsp. austriacum produces copious crops of whitish fleshy fruits that ripen at the beginning of September and remain available for frugivores until early March. This mistletoe shares an assemblage of seed dispersers with a diverse community of fleshy fruited shrubs formed chiefly by Berberis hispanica subsp. hispanica Boiss. & Reut., Crataegus monogyna Jacq, Lonicera arborea Boiss., Juniperus oxycedrus L., Juniperus communis L., Prunus ramburii Boiss, and Rosa sp.
Mistletoe recruitment and seed-deposition sites
We studied the natural seed-deposition pattern of V. album subsp. austriacum on pine branches after a seed-dispersal season (beginning of March 2010). Mistletoe seed rain was measured on 41 branches of 41 parasitized P. nigra trees of similar parasitic loads (hosting from five to ten large-sized fruiting mistletoes). One of the accessible branches per tree was randomly selected for study. Each branch was divided into four positions according to branch thickness: the thickest (>2.5 cm) and nearest position to the trunk (basal position); the middle position of the branch (2.49–1 cm) (middle position); and two apical positions (0.99–0.2 cm), one uncovered (pine twigs 2–3 years old) (apical uncovered), and the other covered by pine needles (the most recent pine growth) (apical covered). Branch diameter was measured with a precision caliper. For each branch position we quantified the number of mistletoe seeds, considering losses due to seed predation as predators feed mostly on the embryo (Grazi and Urech 2000 in Zuber 2004), leaving easily detectable scrapes on the branch.
Also, we estimated the quality of each branch position for mistletoe recruitment by calculating recruitment probabilities at these sites. To do so, we performed a sowing experiment under field conditions. Ten P. nigra trees, alike in age (40 year), size (diameter at breast height, 10.27 ± 1.38 cm), architecture, and ecological environment, were chosen in Sierra de Baza. Three branches per tree were randomly assigned and divided into four positions, following the same criteria as for seed counts. A total of 900 seeds were placed onto the 30 branches: ten seeds in basal positions (>2.5 cm), ten in middle positions (2.49–1 cm), five in uncovered apical positions (0.99–0.2 cm), and five in needle-covered apical positions. Fruits collected in Sierra de Baza during early March 2010 from 15 source plants growing on different host trees were pooled and randomized. Fruit exocarps were removed to permit seed germination (Ladley and Kelly 1996) and seeds were rapidly and carefully placed on branches, leaving 1.5- to 2-cm spaces between seeds. After 15 months (considered sufficient time for seeds to germinate and to become established), we quantified the number of seedlings at each branch position. Then we estimated recruitment probabilities of each site as the number of seedlings counted at the end of the experiment relative to the initially placed seeds.
Identification of mistletoe seed dispersers and seed-deposition patterns
We investigated the behavioral patterns of all members of the frugivore bird assemblage inhabiting the study site, as well as their linkage to mistletoe dispersal. For this, we recorded number of bird visits to parasitized P. nigra and described their feeding and post-foraging behavior on different host structures. Additional bird watching in non-parasitized trees served to compare avian behavioral patterns when mistletoes were present or absent. For two seed-dispersal seasons, from September to February of 2 consecutive years (2009–2010 and 2010–2011), birds were observed by direct observations as well as videotaping. Direct observations, from 0700 to 1200 hours at four localities of the study site, were randomly conducted over the sampling period from hiding places at a minimum distance of 15 m from focal trees (using binoculars when needed for bird identification). For videotaping, high-resolution video cameras were placed directly in the field, from 0700 to 1200 hours. Recordings were made on 48 parasitized P. nigra trees from distances of 5–15 m, enabling bird identifications and accurate descriptions of feeding behavior.
In the study of avian feeding behavior, each time a bird made contact with the mistletoe was considered a “visit.” During each visit, the visitor was identified to the species level, the activity (feeding or perching), fruit-removal rate (number of fruits swallowed per minute) and visit duration were recorded. We disregarded incomplete observations when estimating visit durations, i.e., birds not observed from the moment they arrived at the mistletoe until the moment they left it. After identification, the birds were classified into four guilds (see Table 1) according to their body size [large frugivores of 60–120 g, small frugivores of 12–20 g; after Mullarney et al. (2000)] and resource acquisition [generalist or opportunist frugivore; after Watson (2012)]. The first group, large generalist frugivores (LG) consisted of large birds which were dietary generalists, feeding on a broad range of fruits during autumn and winter. The second group, small generalist frugivores (SG), was composed of small birds that consumed a diverse range of fruits. The third group, opportunists (O), was formed by small birds, mainly granivores and insectivores that occasionally consumed fleshy fruits. Finally, the last group, small mistletoe visitors (SV), was composed of other small birds, either insectivores or granivores, that visit mistletoe but do not manipulate fruits. The variables Number of visits, Visit duration (minutes) and Fruit-removal rate (fruits per minute) were used to estimate the quantity component of the dispersal effectiveness of each visitor species that could potentially act as a seed disperser.
Once the mistletoe visitors were identified, we analyzed their preferences for different host structures, on both parasitized and non-parasitized trees. First, we divided the trees into three portions: upper third (the treetop), middle third, and the bottom of the tree (lower third). Next, we distinguished between four structures visited by birds: the crown of the pine; and three positions within branches—basal, middle and apical. The branches were divided following the same criteria as for the exploration of seed-deposition and seedling-distribution patterns. The frequency of use of different host structures was used to assign locations with higher probabilities of receiving seeds. This, multiplied by the mistletoe recruitment probabilities previously calculated, was used to characterize the dispersal quality of each bird species. By multiplying the “quantity” and the “quality” components, we calculated the dispersal effectiveness of each bird species. Finally, disperser efficiency was estimated by dividing the quality term by the quantity term.
Seed condition
A sowing experiment was performed to evaluate the suitability of mistletoe seeds after they were internally or externally dispersed. For this, we collected bird-dispersed and non-dispersed mistletoe seeds that were placed on safe sites of P. nigra branches. For bird-dispersed seeds, we gathered seeds in the field with clear signs of previous consumption by frugivore birds, expelled either by regurgitation or defecation (simulating an endozoochorous dispersal event); they were mixed to randomization and immediately placed onto tree branches. For non-dispersed seeds, fruits were collected directly from mother plants, the exocarps manually removed (simulating an ectozoochorous dispersal event), and the seeds randomized and placed on host branches. During early March 2010, a total of 900 dispersed seeds and another 900 non-dispersed seeds were inoculated onto twigs (0.99–0.2 cm in diameter) of three limbs of 70 P. nigra trees (all alike in age, size, and architecture); then, seeds were monitored every 30–40 days for up to 15 months after planting. In the first monitoring the number of seeds that were lost before their initial attachment to the host bark was quantified. During seed monitoring, we noted whether a seed was absent or present, and if present, seed condition (alive or dead). We distinguished two stages within live seeds: germinated and established. Germinated included seeds that had started haustoria development, whereas established included seeds with a functional haustoria and emerged cotyledons. Proportions of seed germination (number of germinated seeds vs. the total remaining on tree branches after the first month) and seedling establishment (number of germinated seeds vs. the total remaining on tree branches) were calculated for further analyses.
Statistical analysis
To analyze the distribution of mistletoe seeds and seedlings in the different positions of pine branches, we used generalized linear models (GLMs), assuming a Poisson error distribution and log function, followed by a Tukey honest significant difference (HSD) pair-wise comparison method. The portion of host used (upper third, middle third or bottom third) and host structures used (crown and branch positions: basal, middle or apical) were compared between avian dispersing guilds (LG, SG and O) with a Pearson’s χ2-test. Simulated p-values based on 2,000 replicates were used when needed. Germination and establishment success of internally and externally dispersed seeds were compared using GLMs, assuming a binomial distribution of errors and logit-link functions, followed by a Tukey HSD pair-wise comparison method. R software (version 2.10.0, R Development Core Team 2010) was used to perform the statistical analyses, while figures were made in StatView 5.0.1. Tukey’s pairwise comparisons with 95 % confidence level were conducted with the multcomp package (Hothorn et al. 2008).
Results
Mistletoe recruitment and seed-deposition sites
Recruitment probabilities differed between branch positions (GLM, χ 2 = 66.30, df = 3, P < 0.0001), being nil at the thickest sites (basal and middle positions of branches), 0.67 % for apical uncovered twigs and 17.3 % for needle-covered twigs (Fig 1). Seed deposition also differed between branch positions (GLM, χ 2 = 649.11, df = 3, P < 0.0001), although this followed a different pattern (Fig. 1): 393 seeds (66.16 % of total seeds) were on basal positions, 150 seeds (25.25 %) were on needle-covered apical twigs, only 50 seeds (12.67 %) reached the middle positions, while apical uncovered twigs hardly received one seed (0.002 %).

Pattern of mistletoe seed deposition (black squares) and recruitment probability (gray bars) according to branch diameter. Branches were divided into four categories: >2.5 cm (closest to the tree trunk) (Basal); 2.49–1 cm (Middle); and two apical positions—uncovered, 0.99–0.2 cm (Apical uncovered); and needle-covered, 0.99–0.2 cm (Apical covered). Number of seeds, mean ± SE, n = 41 branches [generalized linear model (GLM), χ2 = 649.11, df = 3, P < 0.0001]; differences among branch positions are indicated by different letters. Recruitment probabilities, mean ± SE, n = 30 branches (GLM, χ2 = 66.30, df = 3, P < 0.0001); ***P < 0.0001
Identification of mistletoe seed dispersers and seed-deposition patterns
During bird observations, we recorded 15 different bird species that visited fruiting V. album subsp. austriacum. Eight species were classified as generalist frugivore seed dispersers, three as opportunists, and four as mistletoe visitors that never manipulated fruits. Of the seed dispersers, six were large and two small birds; opportunists and other visitors were all small (Table 1). All seed dispersers swallowed the fruits and defecated or regurgitated the seeds, while opportunists only pecked at the fruit pericarp and externally dispersed mistletoe seeds that adhered to their feathers or beaks. During 98 videotaping hours, for 48 parasitized P. nigra trees, we recorded 148 birds visiting mistletoe. For 208 h over 50 days of direct bird observations, we recorded another 361 visits to mistletoe on parasitized pines. An additional 564 bird observations were made on non-parasitized trees, which were used to compare the birds’ behavioral patterns when mistletoe was absent. Large generalist frugivores accounted for 90 % of the visits, small generalist frugivores for 4.26 % and opportunists for 5.74 %. From all the bird visits, we clearly discerned 282 feeding events (in which birds were observed pecking or swallowing fruits) of which 50 were clear episodes of endozoochory (in which birds defecated seeds after feeding on whole mistletoe fruits), all involving Turdus spp. and Sylvia atricapilla. Also, we recorded scattered events of ectozoochory by Parus caeruleus (n = 9 events), Parus major (n = 3 events) and Parus ater (n = 4 events). In these cases, as birds pecked at fruits to feed on the pulp, sticky seeds occasionally adhered to their beaks, which were immediately rubbed against pine needles at the tip of the branches to remove the seeds. Quantitatively, thrushes, mainly Turdus viscivorus, were major contributors to mistletoe seed dispersal (Table 1).
The three guilds of birds (“other visitors” were excluded as they showed no interaction with mistletoe fruits) differed in the use of host structures (χ 2 = 731.76, df = 6, P < 0.0001) and frequented different portions of the tree (χ 2 = 207.25, df = 4, P < 0.0001) (Table 2). Small birds (generalist frugivores and opportunists) differed in the use of the host’s structures (χ 2 = 50.11, df = 3, P = 0.0005) but not in the portion of the tree (χ 2 = 2.92, df = 2, P = 0.272). Large generalist frugivores mostly frequented structures located at the treetop (72.5 %), while the majority of small passerines visited medium heights of the pine (82.6 %). About half of the large birds (all Turdus species, except for Turdus merula) visited mistletoes located at the treetop, and fed on fruits while perched on the crown. After feeding, they kept on watch from the crown, moved to a nearby crown, or moved to basal (20 % of the visits) and middle positions (30 % of the visits) of pine branches. T. merula, however, frequently fed on fruits fallen on the ground. Small birds, either generalist frugivores or opportunists, visited the middle portions of the tree. However, while small generalists commonly perched on the middle parts of branches (47 %) and at times used apical twigs (26 %), opportunists followed the opposite pattern, mostly perching on branch tips (85 %) and rarely using thicker structures (15 %). Small opportunists offered the highest quality seed-dispersal services, followed by the large generalist frugivore T. viscivorus and the small generalist frugivore S. atricapilla (Table 2). Overall, T. viscivorus was the most effective seed disperser, while opportunist species were the most efficient dispersers (Table 3).
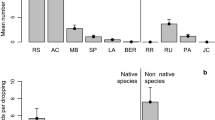
Birds behaved similarly in parasitized and non-parasitized pines regarding the portion of the host used (high, middle, low) by each group of dispersers (LG, χ 2 = 0.37, df = 2, P = 0.83; SG, χ 2 = 10.17, df = 2, P = 0.062; O, χ 2 = 2.28, df = 1, P = 0.13; the low position was eliminated from the latter analysis because of the lack of data). No differences were found, either, in the structures (crown or branch positions: basal, middle and apical) preferred by birds when the host was parasitized or not (LG, χ 2 = 16.73, df = 3, P = 0.051; SG, χ 2 = 2.54, df = 3, P = 0.11; O, χ 2 = 1.28, df = 3, P = 0.26) (Fig. 2). Overall, the preferred structure by LG was the tree crown, i.e., 41 % of the visits in parasitized trees and 53 % in non-parasitized.

Post-foraging microhabitat use of mistletoe seed dispersers on parasitized and non-parasitized trees. Shown is the relative use frequency of different host structures by major groups of mistletoe seed dispersers. Tree structures are indicated as: treetop (Crown); and three different positions within branches—closest position to the tree trunk (Basal), middle position (Middle), branch periphery (Apical). Groups of seed dispersers are shown as large generalist frugivores (LG), including Turdus viscivorus, Turdus torquatus, Turdus philomelos, Turdus merula, Turdus iliacus and Turdus pilaris; small generalist frugivores (SG), including Sylvia atricapilla and Erithacus rubecula; opportunists (O), including Parus ater, Parus major and Parus caeruleus. Pearson's χ 2-test, LG, χ 2 = 16.73, df = 3, P = 0.051; SG, χ 2 = 2.54, df = 3, P = 0.11; O, χ 2 = 1.28, df = 3, P = 0.26. Sample sizes are shown above the bars
Seed condition
With regard to seed handling, no differences were found between treatments in seed germination and seedling establishment. Of the seeds remaining on host branches (disregarding seeds that failed in their initial attachment to host branches), 69 % of internally dispersed seeds and 71.4 % of those externally dispersed germinated (GLM, χ 2 = 0.17,df = 1, P = 0.68), while 8.2 % of externally dispersed seeds and 5.6 % of internally dispersed seeds successfully established (GLM, χ 2 = 2.24, df = 1, P = 0.14).
Discussion
This study explicitly examines the poorly explored topic of how generalist birds, in the absence of specialized dispersers, provide seed dispersal for a parasitic plant with strong recruitment constraints. We first identified the potential group of seed dispersers of V. album subsp. austriacum in a Mediterranean mountain of southeastern Spain. Then, by linking species-specific behavior to seed-dispersal patterns and to demographic consequences for the parasite in different microhabitats, we discovered that this mistletoe can rely on unspecialized vectors to disperse seeds to narrowly defined safe sites.
A broad and diverse group of 11 bird species (except for strict granivores or insectivores) disperse seeds of V. album subsp. austriacum in Sierra de Baza. Any bird can be a potential disperser, as the seeds do not need to be ingested to germinate. This flexibility guarantees that V. album subsp. austriacum, irrespective of the composition and abundance of the avian community in a pine forest, will satisfy its seed-dispersal needs. Regardless of the dispersal mode, all vectors have to deposit seeds on proper safe sites—narrowly limited to thin branches of the tree periphery—in order to exert a real impact on mistletoe demography. Small epizoochorous and endozoochorous birds disperse, respectively, by rubbing their beaks against pine needles or directly defecating on twigs. Meanwhile, large endozoochorous birds disperse by haphazardly and abundantly defecating or regurgitating from the treetop. In short, any bird able to deposit a seed on a suitable site qualifies as a seed disperser.
Among endozoochorus vectors, large generalist frugivores constitute the most effective dispersal guild; however, they provide low-efficiency services, consuming numerous fruits in order to suitably disperse a single seed. Their preferences for firm and robust structures as perching sites finally result in abundant seed rain with most seeds reaching basal, thicker parts of branches. At these sites the seeds have no chance of survival, the haustorium being unable to penetrate the xylem of the host, and furthermore are likely to become desiccated by extreme summer temperatures or to be depredated (Mellado and Zamora 2014). On the other hand, as thrushes provide high-quantity dispersal services, the few times (1.1 %) they used apical sites of branches led to a considerable number of favorable dispersal events, which notably contributed to the final recruitment of the parasite. In the light of these results, large generalist frugivores might act as effective seed dispersers whenever mistletoe fruit availability is sufficient to offset their high inefficiency. V. album subsp. austriacum can afford this wasteful dispersal mechanism by producing abundant and constant crops of small fruits over most of the fruiting season (Zuber 2004). In contrast, although small generalist frugivores provide better quality and more efficient dispersal, they were so scarce that their contribution to the parasite was almost negligible during our study. On the other hand, opportunist tits can be considered very efficient dispersers by moving seeds externally and directly to safe sites of the tree periphery. This external seed transport by dietary generalists and opportunist frugivores has been frequently reported among Viscaceae mistletoes (e.g., Punter and Gilbert 1989; Restrepo et al. 2002), which may confer several advantages to the parasite. First, single deposition onto safe sites may help seeds to escape high post-dispersal seed predation—both because of lower seed densities (Davidar 1983) and, particularly in this system, because of the higher protection of seeds under needle coverage, where they have optimum recruitment conditions (Mellado and Zamora 2014). Secondly, once seedlings become established, sibling competition may be diminished (Davidar 1983). In this sense, the demographic consequences of tits would be akin to those of mistletoe specialists, if not for the scarcity of the dispersal events. This indicates that morphological adaptations do not necessarily confer advantages for mistletoe dispersal over less specialized vectors, and further highlights the great importance that unexpected dispersal agents can have for the demography of a plant (Calviño-Cancela 2002; Heleno et al. 2011; Frick et al. 2013).
An interesting finding is that birds visit parasitized as well as non-parasitized pines, behaving similarly on both. This, coupled with the large home ranges of dispersal vectors (especially thrushes) and the wide and heterogeneous range of seed dispersers, may increase the overall dispersal effectiveness of the mistletoe population, with several bird species moving seeds at different distances; this ensures local population functions, including reinfection processes, and the establishment of new populations or expansion of the plant’s range, which might relieve host populations from intense local reinfection processes (Watson and Rawsthorne 2013). V. album subsp. austriacum, being able to cope with massive seed losses (by producing large fruit crops) and to accept any dispersal mechanism (either endozoochorous or ectozoochorous), can achieve effective dispersal by an unspecialized dispersal group. In conclusion, the mutualistic interaction between the European mistletoe and zoochorous birds is a generalized system; almost all birds feeding on fruits can act as effective seed dispersers. Plant dispersal success relies on the correspondence between a large fruit crop and a diverse guild of endozoochorous and/or ectozoochorous dispersal vectors. These generalistic features enable the spatio-temporal replacement of dispersers, allowing the presence of Viscaceae species in geographical ranges lacking specialist dispersers, such as occur in Europe.
References
Amico G, Aizen MA (2000) Mistletoe seed dispersal by a marsupial. Nature 408:929–930. doi:10.1038/35050170
Arruda R, Carvalho LN, Del-Claro K (2006) Host specificity of a Brazilian mistletoe, Struthanthus aff. Polyanthus (Loranthaceae), in cerrado tropical savanna. Flora 201:127–134
Arruda R, Fadini R, Carvalho LN, Del-Claro K, Mourão FA, Jacobi CM, Teodoro G, Van den Berg E, Caires CS, Dettke GA (2012) Ecology of Neotropical mistletoes: an important canopy-dwelling component of Brazilian ecosystems. Acta Bot Bras 26:264–274
Aukema JE, Martínez del Río C (2002) Where does a fruit-eating bird deposit mistletoe seeds? Seed deposition patterns and an experiment. Ecology 83:3489–3496
Bascompte J, Jordano P (2007) The structure of plant–animal mutualistic networks: the architecture of biodiversity. Annu Rev Ecol Evol Syst 38:567–593
Calviño-Cancela M (2002) Spatial patterns of seed dispersal and seedling recruitment in Corema album (Empetraceae): the importance of unspecialized dispersers for regeneration. J Ecol 90:775–784. doi:10.1046/j.1365-2745.2002.00711.x
Carlo TA, Aukema JE (2005) Mistletoe seed dispersal by a marsupial. Ecology 86:3245–3251. doi:10.1890/05-0460
Davidar P (1983) Birds and neotropical mistletoes: effects on seedling recruitment. Oecologia 60:271–273
Frick WF, Price RD, Heady PA, Kay KM (2013) Insectivorous bat pollinates columnar cactus more effectively per visit than specialized nectar bat. Am Nat 181:137–144. doi:10.1086/668595
Gómez JM, Zamora R (2000) Spatial variation in the selective scenarios of Hormathophylla spinosa (Cruciferae). Am Nat 155(5):657–668
Hawksworth FG, Wiens D (1996) Dwarf mistletoes: biology, pathology, and systematic. Agriculture handbook no. 450. Forest Service, US Department of Agriculture, Washington DC
Heleno RH, Ross G, Everard AMY, Memmott J, Ramos JA (2011) The role of avian ‘seed predators’ as seed dispersers. Ibis 153:199–203
Herrera CM (1988) Variation in mutualisms: the spatio-temporal mosaic of a pollinator assemblage. Biol J Linn Soc 35:95–125
Hothorn T, Bretz F, Westfall P (2008) Simultaneous inference in general parametric models. Biometr J 50:346–363
Ladley JJ, Kelly D (1996) Dispersal, germination and survival of New Zealand mistletoes (Loranthaceae) dependence on birds. N Z J Ecol 20:69–79
Larson D (1996) Seed dispersal by specialist versus generalist foragers: the plant’s perspective. Oikos 76:113–120
Martínez del Río C, Restrepo C (1993) Ecological and behavioral consequences of digestion in frugivorous animals. Vegetatio 107:205–216
Martínez del Río C, Hourdequin M, Silva A, Medel R (1995) The influence of cactus size and previous infection on bird deposition of mistletoe seeds. Aust J Ecol 20:571–576
Mellado A, Zamora R (2014) Linking safe sites for recruitment with host-canopy heterogeneity: the case of a parasitic plant, Viscum album subsp. austriacum (Viscaceae). Am J Bot 101(6):1–8. doi:10.3732/ajb.1400096
Montaño-Centellas FA (2012) Effectiveness of mistletoe seed dispersal by tyrant flycatchers in a mixed Andean landscape. Biotropica 45:209–216. doi:10.1111/j.1744-7429.2012.00909.x
Mullarney K, Svensson L, Zetterström D, Grant PJ (2000) Birds of Europe. Princeton University Press, Princeton
Murphy SR, Reid N, Yan Z, Venables WN (1993) Differential passage time of mistletoe fruits through the gut of honeyeaters and flowerpeckers: effects on seedling establishment. Oecologia 93(2):171–176
Norton DA, de Lange PJ (1999) Host specificity in parasitic mistletoes (Loranthaceae) in New Zealand. Funct Ecol 13:552–559. doi:10.1046/j.1365-2435.1999.00347.x
Olesen JM, Jordano P (2002) Geographic patterns in plant-pollinator mutualistic networks. Ecology 83(9):2416–2424
Punter D, Gilbert J (1989) Animal vectors of Arceuthobium americanum seed in Manitoba. Can J For Res 19:865–869
R Development Core Team (2010) R Foundation for statistical computing. Austria, Vienna
Rawsthorne J, Watson DM, Roshier DA (2011) Implications of movement patterns of dietary generalist for mistletoe seed dispersal. Austral Ecol 36:650–655. doi:10.1111/j.1442-9993.2010.02200.x
Reid N (1989) Dispersal of mistletoes by honeyeaters and flowerpeckers: components of seed dispersal quality. Ecology 70:137–145. doi:10.2307/1938420
Reid N (1990) Mutualistic interdependence between mistletoes (Amyema quandang), and spiny-cheeked honeyeaters and mistletoebirds in an arid woodland. Aust J Ecol 15:175–190. doi:10.1111/j.1442-9993.1990.tb01526.x
Reid N (1991) Coevolution of mistletoe and frugivorous birds. Aust J Ecol 16:457–469. doi:10.1111/j.1442-9993.1991.tb01075.x
Restrepo C (1987) Aspectos ecológicos de la diseminación de cinco especies de muérdagos por aves. Humboldtia 1:65–116
Restrepo C, Sargent S, Levey DJ, Watson DM (2002) The role of vertebrates in the diversification of new world mistletoes. In: Levey DJ, Silva WR, Galetti M (eds) Seed dispersal and frugivory: ecology, evolution and conservation. CAB International, Wallingford, pp 83–98
Richardson KC, Wooller RD (1988) The alimentary tract of a specialist frugivore, the mistletoebird, Dicaeum hirundinaceum, in relation to its diet. Aust J Zool 36:373–382
Sargent S (1995) Seed fate in a tropical mistletoe: the importance of host twig size. Funct Ecol 9:197–204
Schupp EW (1993) Quantity, quality and effectiveness of seed disper-sal by animals. Vegetatio 107:15–29
Snow BK, Snow DW (1984) Long-term defense of fruit by mistle thrushes (Turdus viscivorus). Ibis 126:339–349
Snow BK, Snow DW (1988) Birds and berries. A study of an ecological interaction. Poyser, Calton
Van Z (1993) Germination and seedling development of two mistletoes, Amyema preissii and Lysiana exocarpi: host specificity and mistletoe-host compatibility. Aust J Ecol 18:419–429. doi:10.1111/j.1442-9993.1993.tb00469.x
Walsberg GE (1975) Digestive adaptations of Phainopepla nitens associated with the eating of mistletoe berries. Condor 77:169–174
Waser NM, Chittka L, Price MV, Williams NM, Ollerton J (1996) Generalization in pollination systems, and why it matters. Ecology 77(4):1043–1060
Watson DM (2004) Mistletoe—a unique constituent of canopies worldwide. In: Loman M, Rinker B (eds) Forest canopies, 2nd edn. Academic Press, New York, pp 212–223
Watson DM (2012) The relative contribution of specialists and generalists to mistletoe dispersal: insights from a neotropical rain forest. Biotropica 45:195–202. doi:10.1111/j.1744-7429.2012.00905.x
Watson DM, Rawsthorne J (2013) Mistletoe specialist frugivores: latterday ‘Johnny Appleseeds’ or self–serving market gardeners? Oecologia 172:925–932. doi:10.1007/s00442-013-2693-9
Wenny DG (2001) Advantages of seed dispersal: a re-evaluation of directed dispersal. Evol Ecol Res 3:51–74
Zuber D (2004) Biological flora of Central Europe: viscum album L. Flora 199:181–203
Acknowledgments
We are grateful to Ramón Ruiz Puche for his excellent work in the field, Tomás A. Carlo for his constructive comments on an earlier version of this manuscript, Julia Koricheva, David M. Watson and Rafael Arruda for their helpful comments as reviewers of the manuscript, and David Nesbitt for checking the English of this paper. We thank the Consejería de Medio Ambiente (Junta de Andalucía) and the Director of the Sierra de Baza Natural Park for permission for the field work. This study was supported by the Spanish Ministry of Science and Innovation (MICINN) projects MUERDAME CGL2008-04794 and CLAVINOVA CGL2011-29910 and a FPI grant BES-2009-023219 to A. M. G. from the Spanish MICINN.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Julia Koricheva.
Rights and permissions
About this article

Cite this article
Mellado, A., Zamora, R. Generalist birds govern the seed dispersal of a parasitic plant with strong recruitment constraints. Oecologia 176, 139–147 (2014). https://doi.org/10.1007/s00442-014-3013-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00442-014-3013-8